
Abstract
Embryologic events in mammalian myogenesis remain to be fully defined. Recent evidence supports the presence of a common progenitor arising in the dermomyotome that gives rise to both embryologic and adult muscle and postnatal myogenic stem cells (satellite cells). In this study, we utilize the technique of early intra-amniotic gene transfer to target nascent muscle progenitors as they traverse the primitive streak before formation of the dermomyotome. This technique robustly transduced both epaxial and hypaxial muscle groups. Marker gene expression is observed in up to 100% muscle fibers in the lower extremities and is sustained for the lifetime of the mouse. We next analyzed transduced muscle for satellite cell transduction using highly sensitive methodology. Surprisingly, despite high levels of sustained transgene expression in muscle fibers, satellite cells lacked the marker transgene. Our data suggest that dermatomyotome is a heterogeneous structure and that not all myogenic progenitors of dermatomyotome give rise to satellite cells.
Introduction
T
Embryologically, our injections were performed at E7.5–E8, which corresponds to Theiler stage 11–13, which can be confirmed by ultrasound at the time of injection (Fig. 1A, C). This is the presomitic or late stage of gastrulation when cells streaming through the caudal aspect of the remaining primitive streak colonize the paraxial mesoderm that subsequently segments into the more posterior somites that ultimately give rise to the lower body and limb musculature (Fig. 1B) [5,6]. Our injections into the amniotic cavity target cells, like this presomitic epiblast traversing the primitive streak that will become the mesoderm of the lower body. This is our best explanation of how intra-amniotic injection results in sustained (life-long) transgene expression in a high percentage of muscle fibers in the trunk and lower limbs without expression in the upper body. This technique therefore represents an opportunity for fate mapping of this very primitive cell population that ultimately gives rise to the skeletal muscle compartment.
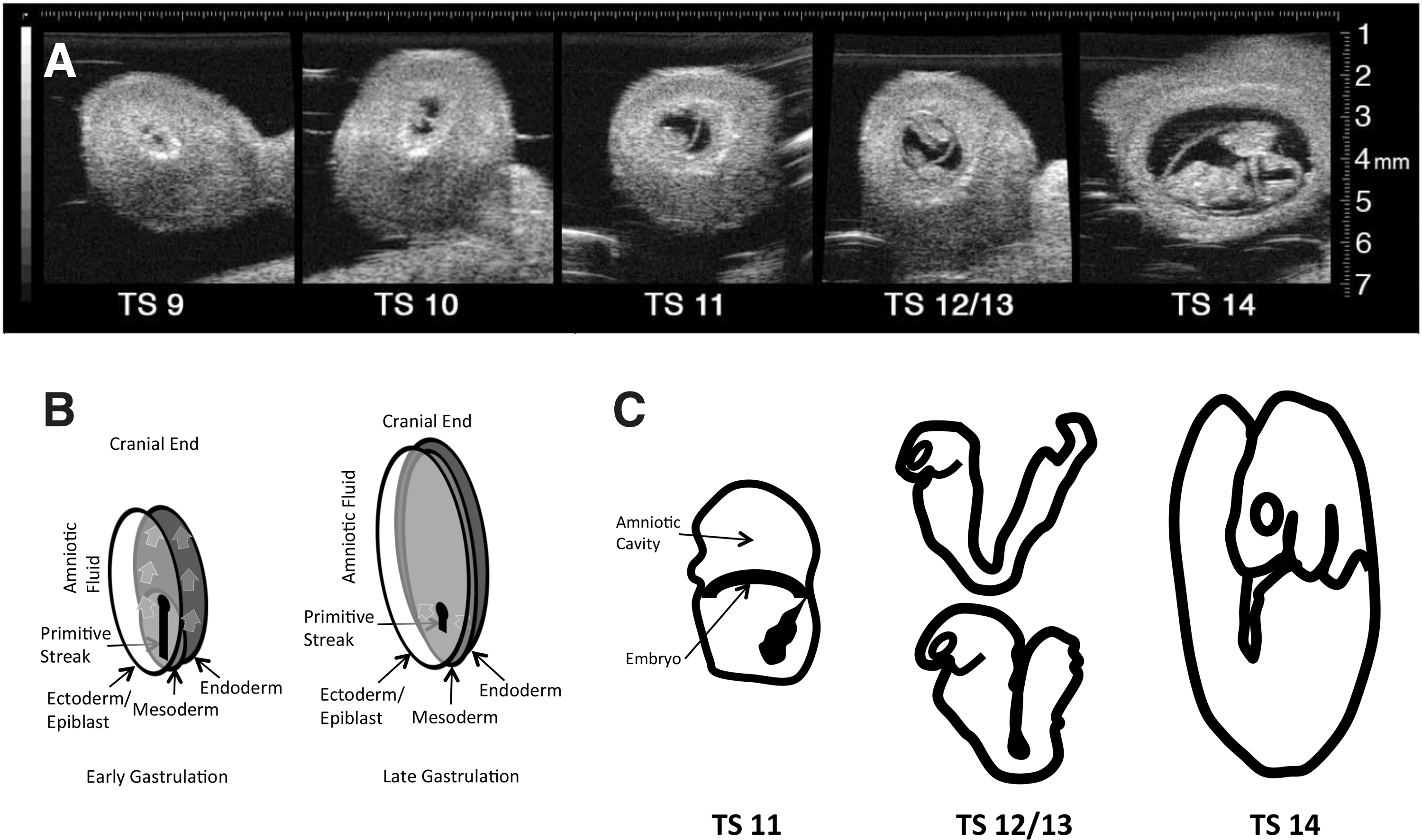
Ultrasound images of gravid mouse uterus following laparotomy at different gestational ages.
In mice, the muscular migration to the limbs occurs in two stages, an embryonic and fetal phase, starting on postcoital day 11 (E11). First, embryonic myoblasts generate the primary muscle fibers that act as a template for muscle growth. From E14 until the early postnatal period, in the second wave, fetal myoblasts, which give rise to adult muscle and satellite cells, migrate to the template laid down by the embryonic primary myoblasts. These satellite cells are committed progenitor cells that are responsible for the maintenance, growth, repair, and regeneration of postnatal skeletal muscles [7,8]. Satellite cells are thought to primarily arise from cells in the dermomyotome of the developing somite [9 –12].
In this study, we assess whether the predominant postnatal myogenic progenitor cell population (satellite cells) is derived from epiblast cells that ingress through the primitive streak to form postnatal myofibers.
Materials and Methods
Mice (Mus musculus)
The C57BL/6 (Jackson Laboratories), GFP mice (a kind gift from M. Okabe, Osaka University) [13] or Myf5nlacZ/+ mice (a kind gift from M.A. Rudnicki, University of Ottawa) used in this study were mated in our breeding colony. To achieve accurate time-dating mice were mated overnight (10 h) and separated in the morning (E0). Mice were then palpated at E8 for pregnancy. Myf5nlacZ/+ knock in mice have the gene for nuclear localized beta-galactosidase (B-gal) inserted into the Myf5 locus resulting in nuclear expression of B-gal restricted to satellite cells. Myf5 nlacZ/+ mice were generated by crossing Myf5 nlacZ/+ mice since Myf5 nlacZ/nlacZ is embryonic lethal. The Myf5nlacZ/+ mice used in this study had been backcrossed to a C57BL/6 background for 6–8 generations. GFP mice were crossed with Myf5nlacZ/+ mice and offspring were backcrossed to generate double-transgenic GFP/Myf5nlacZ/+ mice. Pups were genotyped postnatally by polymerase chain reaction (PCR). The experimental protocols were approved by the Institutional Animal Care and Use Committee at The Children's Hospital of Philadelphia and followed guidelines set forth in the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Lentiviral vector
The starting materials for generating a self-inactivating HIV-1-based vector were kindly provided by Inder Verma (Salk Institute). Modifications of the CS-CG vector included deletion of the remaining right U3 region except for 23 nucleotides downstream of the 3′ppt, deletion of the residual envelope and ancillary gag/pol sequences, insertion of the central DNA FLAP, insertion of the Rev response element, and insertion of the Woodchuck hepatitis virus post-transcriptional regulatory element, which is modified eliminating the initiation codon for the Woodchuck hepatitis X protein. The EGFP (Clontech Laboratories) was located downstream of the human cytomegalovirus (CMV) immediate early promoter and modified so that all stop codons from the transcription start site to the translation initiation site were removed. Viral vectors pseudotyped with vesicular stomatitis virus envelope were generated as previously described [2].
Intra-amniotic vector injection at E8
Gestational day 8 (E8) fetuses underwent intra-amniotic injection with vector using an ultrasound-guided injection system (Vevo® 660; VisualSonics) [2]. Pregnant mice were anesthetized with isoflurane and placed supine on the ultrasound table. The abdominal hair was removed by a chemical hair remover (Nair®; Church & Dwight) and the surgical area was disinfected with alcohol. A 1 cm ventral midline incision was made through the skin, abdominal wall, and peritoneum and a segment of the uterus containing fetuses was exteriorized and covered in prewarmed sterile ultrasound gel (Acquasonic®; Parker Laboratories). The fetuses were scanned using a 40 MHz probe to confirm appropriate developmental stage. With ultrasound the preturning Theiler stages 12/13 could be distinguished from the post-turning Theiler stage14 (Fig. 1A, C). Only preturning mice were injected. Pulled and beveled glass microcapillary pipettes (outer diameter 1.14 mm, inner diameter, 0.53 mm; Humagen) were prefilled with mineral oil (Sigma), connected to the micropipette holder, and loaded with vector. Under two-dimensional visualization the micropipette tip was inserted through the uterine wall into the amniotic cavity. Three hundred fifty nanoliters of vector (2×1010 TU/mL) was injected using an automated syringe and the micropipette was retracted. This procedure was repeated until all fetuses were injected. Upon completion of the injections, the uterus was returned to the abdominal cavity and the incision was closed in two layers using running 4-0 Vicryl sutures.
Stereoscopic fluorescent microscopy
We used fluorescent stereomicroscopy (MZ16FA; Leica) for macroscopic assessment of GFP expression in mice after intra-amniotic gene transfer (IAGT). Mice were euthanized in the newborn period (P1–P7) or at time points later in life and the skin immediately dissected off the muscle compartments to allow direct visualization of the muscle. To allow comparison of the fluorescence intensity, exposure time and camera gain were maintained at the same settings for all images.
Immunofluorescent histology
Adult skeletal muscles were fixed in 1.5% formalin in 0.1 M phosphate buffer for 16 h at room temperature. Fetuses harvested at E12, E14, and E16 were fixed in 4% formalin in phosphate buffer for 30 min on ice. All tissues were cryopreserved in 20% sucrose for 24 h. Tissues were frozen in TissueTek OCT-embedding medium (Miles) and sectioned using a cryostat (Leica). Fifteen micrometers sections were blocked in 20% normal goat serum (NGS) in PBS with 0.3% Triton X-100, and stained with primary and secondary antibodies in 5% NGS in PBS with 0.1% Triton X-100. Antibodies and concentrations were as follows: chicken anti-GFP (1:1,000; Abcam), rabbit anti-B-gal (1:1,000; Invitrogen), rat anti-Beta2-Laminin (1:1,000; Chemicon), mouse anti-MF-20 [1:1,000; Developmental Studies Hybridoma Bank (DSHB)], and mouse anti-myogenin (1:1,000; DSHB). Secondary antibodies with Alexa conjugates were from Molecular Probes (1:1,000; Invitrogen).
Fluorescence-activated cell sorting analysis
After euthanasia, muscle from mice injected in utero was harvested for satellite cell sorting at P10. Limb and axial muscles were removed from euthanized animals and placed in PBS on ice. Muscles were minced and mixed with 800 μL of Collagenase I–(616 μg/mL, #C0130; Sigma)/dispase Dispase II (2.4 μg/mL, #04942078001; Roche)/CaCl2 (2.5 mM; Sigma)/Hank's balanced salt solution (HBSS) Enzyme mix solution (#14175; Gibco). The digested tissue was filtered through a 70 μm nylon mesh in a sterile funnel. Cells were centrifuged, resuspended in 2–4 mL of HBSS/4% fetal bovine serum (#SH3007.03; Hyclone)/HEPES Buffer (10 mM, #15630; Gibco), filtered again, counted, and exposed to antibodies. Antibodies and reagents utilized for satellite cell analysis and sorting were as follows: anti-Mouse/Human CD45R-PECY7 (Clone #RA3-6B2, #25-0452-82; EBioscience); anti-Mouse Gr-1 (Ly-6G)-PECY7 conjugated (Clone #RB6-8C5, #25-5931-82; EBioscience); anti-Mouse TER-119 (Ly-76)-PECY7 conjugated (Clone #TER-119, #25-5921-82; EBioscience); anti-Mouse CD5 (Ly-1)-PECY7 conjugated (Clone #53-7.3, #25-0051-81; EBioscience); anti-Mouse Sca-I (Ly-6A/E)-PECY7 conjugated (Clone #D7, #25-5981-81; EBioscience); anti-Mouse CD34-Alexa 700 conjugated (Clone #RAM34, #56-0341-82; EBioscience); anti-Mouse a7 Integrin-Biotin conjugated (Clone #CA5.5; Sierra Bioscience); and streptavidin-Alexa 750 (#S21008; Invitrogen). Nonspecific Fcγ receptor binding was blocked with antibody against mouse Fcγ receptor 2.4G2. Isotype- and conjugate matched antibodies with irrelevant specificities served as negative controls. Propidium iodide staining was used to exclude dead cells. Analytical flow cytometry was performed on a FACSCalibur (BD Biosciences) flow cytometer and cell sorting was performed on a FACSAria cell sorter (BD Biosciences).
Quantification of satellite cell expansion
The lower limbs of C57BL/6 mice were irradiated (18cGy) with an XRad irradiator. One hundred sorted satellite cells in 10 μg/mL Notexin in 10 μL HBSS were injected into the tibialis anterior muscle (TA) in the 24-h period following the irradiation. The TA muscle were harvested at 21 days postinjection and either fixed in 1.5% paraformaldehyde or immediately processed for DNA analysis. Controls included irradiated mice that received HBSS injections. The entire TA was sectioned and a section was taken every 200 μm for B-gal staining (as described above) and all B-gal-positive cells were counted and the total muscle area was measured. The total number and density of B-gal cells was calculated by assuming a linear transition in muscle area and in the number of B-gal-positive cells between each counted tissue section.
Quantitative polymerase chain reaction for GFP DNA
To look for gene silencing in GFP-negative satellite cells, sorted satellite cells from injected or GFP/Myf5 nlacZ/+ mice were analyzed for GFP DNA. DNA was also isolated from the TA muscle of injected mice, transgenic GFP mice, and uninjected C57/BL6 mice using the standard Ambion Back Extraction Protocol associated with the standard RiboPure RNA isolation (Ambion). Briefly, DNA was isolated by homogenizing cells or a portion of the TA in TRIzol buffer, centrifugation, extraction of DNA from the interphase/organic phase in guanidine buffer (4 M guanidine thiocyanate, 50 mM sodium citrate, and 1 M Tris), followed by a standard DNA isopropanol precipitation and ethanol wash to isolate DNA. All GFP DNA was quantified by standard quantitative polymerase chain reaction (Q-PCR) using TaqMan probes (Applied Biosystems) for GFP using a standard 40-cycle amplification protocol on an Applied Biosystems 7500 Fast Q-PCR machine and normalized to 18 s. To establish a standard curve of GFP DNA detection within a sample, splenocytes from GFP mice were diluted in C57BL/6 splenocytes and total GFP DNA content was quantified. Q-PCR results were compared with an unpaired Student's t-test.
GFP protein quantification
GFP protein was measured on a per myofiber basis by quantifying the average fluorescent emission over the total cross-sectional area of each myofiber with a typical myofiber resolved by about 30 pixel scans. Specifically, a Zeiss 510 confocal microscope was used to image 20 μm thick sections of lightly fixed skeletal muscle. To quantify GFP protein, the tissue sections were excited with 488 nm light (Argon laser) and emissions were collected in the range of 505–545 nm using a series of two HQ and one sputtered filter. Emission photons were quantified with a photomultiplier tube in which the linear dynamic range (as defined by <5% deviation from a “best fit” line generated by standard regression) had been mapped at all gain settings. Because the linear dynamic range of the photomultiplier tubes (PMT) was less than the range of intensity values observed, emissions were quantified at four gain settings in which there was 50% overlap in the linear dynamic range with the neighboring gain setting. Emission quanta detected at all four gain settings were then reverse mapped using the previously generated gain-intensity maps to create a single plot of emissions that was linear over the entire range observed for all myofibers in all experimental and control tissue sections. This same approach was used to quantify serial dilutions of soluble GFP to calibrate the linear PMT response to actual concentration changes.
Results
Early IAGT with lentiviral vector results in sustained high-level transgene expression in lower body muscle
We have previously described the general survival, toxicity, and distribution of transduction using the E8 IAGT approach with lentiviral vectors [4]. We have also demonstrated the transduction of stem cell populations in the three tissues reported to date: skin, central nervous system (CNS), and eye [1,2,4]. In this study we focused on the skeletal muscle compartment.
Following our time-dating strategy with overnight mating and early morning (7 AM) injections on E8, the majority of litters were observed by ultrasound to be preturning (T11–T13). However, if injections were delayed until afternoon, then approximately half of fetuses had turned (T14). No litters with turned fetuses were injected (Fig. 1A, C). Whole body examination of animals that underwent IAGT under a fluorescent stereoscopic microscope revealed strong GFP fluorescence of caudal paraxial and lower limb muscle compartments (Fig. 2C–F). This fluorescence extended cranially as high as the trapezius and latissimus dorsi (Fig. 2B). The upper limb muscles, intercostal muscles, upper deep back muscles, and diaphragm were not transduced (Fig. 2A, B). Within the leg, which contained the most robustly transduced muscles, GFP was directly visualized in most myofibers without antibody staining and in some animals 100% of myofibers clearly contained GFP by direct visualization (Fig. 2G). Despite strong GFP fluorescence in the lower body, no extension of GFP expression to the upper limb muscles was observed. Animals were euthanized for analysis at intervals of up to 2 years of age and maintained a similar pattern and degree of GFP fluorescence throughout their lives.
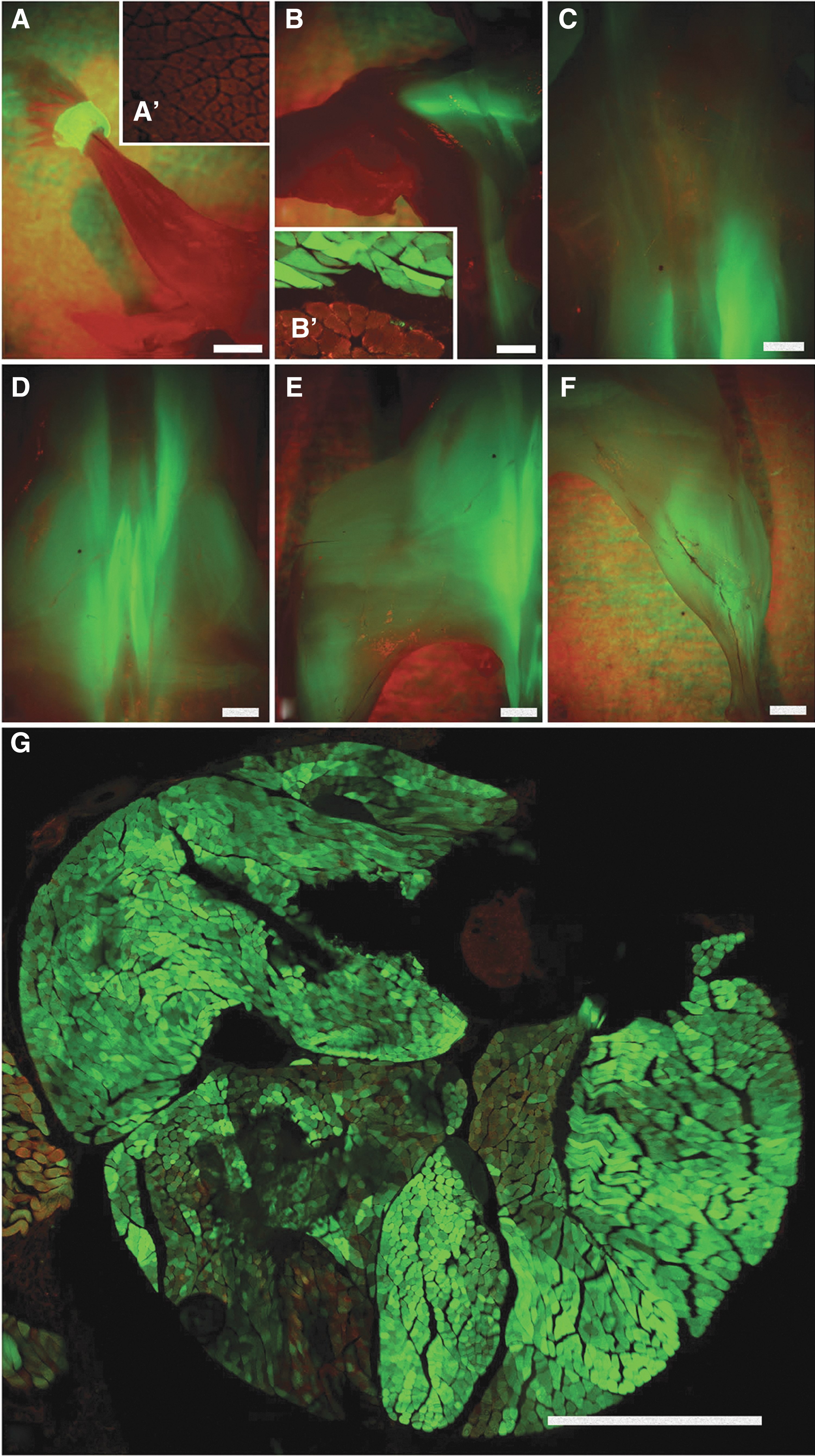
Transduction patterns in a 1-month-old mouse following E8 intra-amniotic injection.
Absence of GFP-positive satellite cells associated with GFP-positive muscle fibers by histology
To assess whether the cells transduced in the primitive streak gave rise to satellite cells, Myf5 nLacZ/+ mice that express nuclear localized B-gal in satellite cells underwent E8 IAGT with lentivirus-GFP. Strongly GFP-positive muscles from 3-month-old injected mice were analyzed by immunohistochemistry and confocal microscopy for GFP, B-gal, and beta-2-laminin. Images were taken with 40×or 100×objectives and the pinhole was set to maintain optical sections of less than or equal to 1 μm to unequivocally determine co-localization of signals. One thousand two hundred B-gal-positive cells were identified in the anatomic position of satellite cells and none were GFP positive by confocal imaging or visual inspection (Fig. 3A–J).

Absence of GFP-expressing satellite cells in recipients of E8 IAGT.
Tracking of GFP expression in the fetal muscle compartment
We assessed the kinetics and distribution of GFP fluorescent cells that had been transduced in the primitive streak and were destined to have a muscle fate. Mice undergoing IAGT at E8 were analyzed at E12, E14, and E16. The fetal muscle compartment was identified by staining for myogenin. In the fetal hindlimbs and somites at E12 and E14, no GFP-positive muscle cells could be identified (Fig. 3L–Q). However, by E16 GFP-positive cells in the paraxial muscles were clearly present (Fig. 3K).
Absence of transduced satellite cells by flow cytometric and PCR analyses
Satellite cells can be prospectively identified in dissociated skeletal muscle by the expression of alpha7-integrin and CD34 and the lack of lineage markers [14]. To further confirm the absence of transduced satellite cells in the transduced muscle compartments, single cell suspensions were prepared from muscles that were strongly GFP positive by stereomicroscopy and stained for the satellite cell lineage markers (Fig. 4A, B). Within the putative satellite cell population, only 0.8% fell within the GFP gate and of these none expressed an emission intensity higher than the dimmest 20% of satellite cells from a GFP transgenic mouse. Given this distribution it is likely that most of these cells fell within the GFP gate due to autofluorescence or random detector noise as opposed to low-level GFP expression (Fig. 4D).
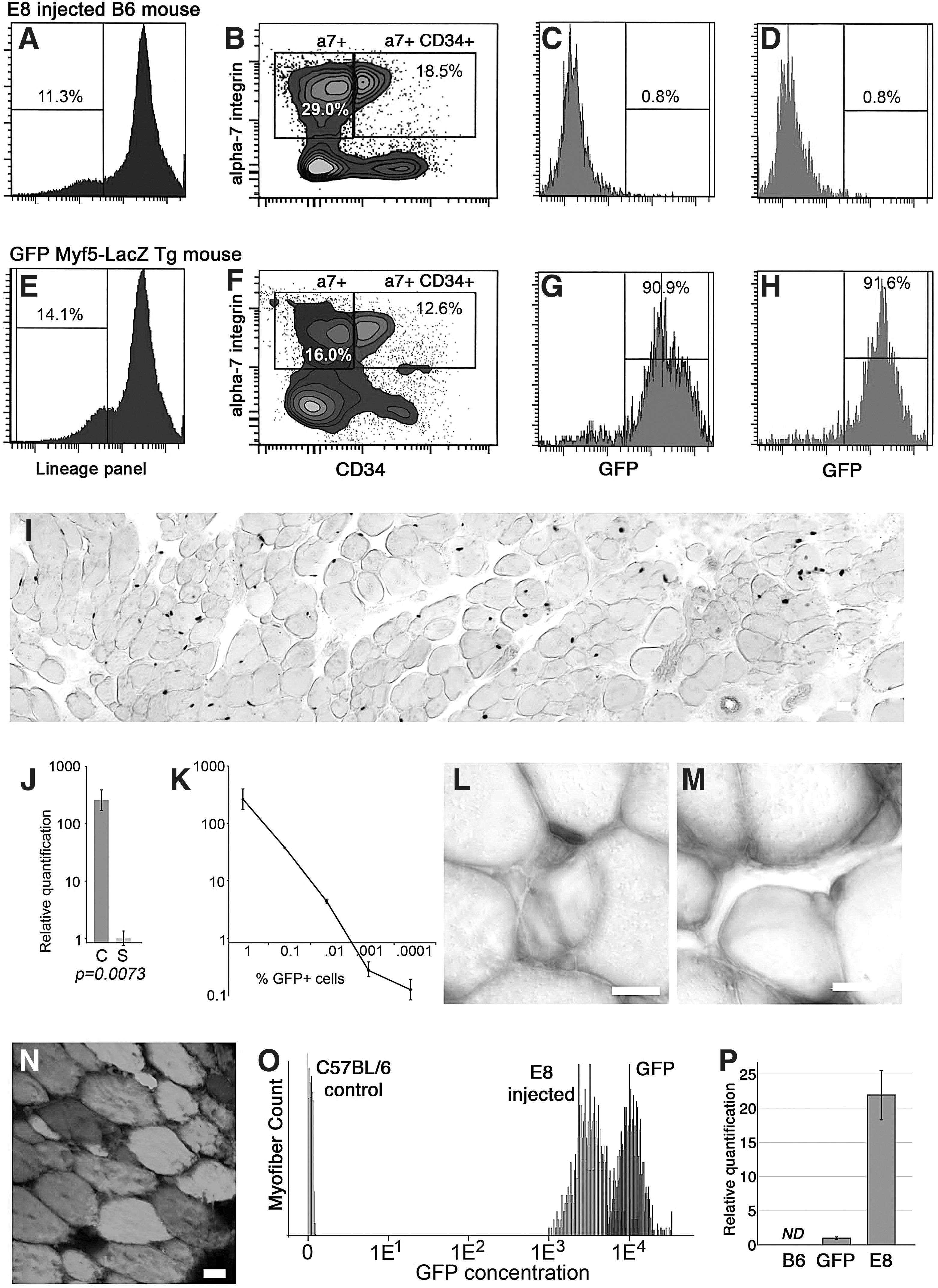
Absence of GFP within isolated satellite cells. Fluorescence-activated cell sorting (FACS) isolation of satellite cells from mice injected with GFP-encoding lentivirus at E8
Because transduction does not necessarily result in marker protein production, particularly within stem cell populations, the sorted satellite cells were analyzed for the presence of GFP DNA. Within the GFP-negative population of satellite cells, GFP was detected at 30.9 cycles, which by comparison with a standard curve of GFP cells from a GFP transgenic mouse mixed in limiting dilution with GFP-negative cells corresponds to an approximate frequency of one GFP cell per 400 total cells (Fig. 4J, K). When GFP transgene was quantified in the total tibialis anterior of 3-month-old mice that had received E8 IAGT, there was a 22-fold higher quantity of GFP transgenes compared with GFP transgenic mice (Fig. 4P). Taken together, in adult mice that had received E8 IAGT, the combined PCR analyses indicate that only extremely rare cells within the sorted satellite cell population were transduced (1 cell per 8,800 if it is assumed that there are 44 GFP transgenes per transduced satellite cell). We consider these results to be strongly negative for the presence of transduced satellite cells by both GFP protein and DNA.
To confirm that the sorted satellite cells met the functional definition of satellite cells, cells were sorted from Myf5-LacZ recipients and 100 Lin(−) a7+CD34+ satellite cells were injected into the irradiated, Notexin-injured tibialis anterior of wild-type B6 mice. The sorted satellite cells readily engrafted and expanded to repopulate the satellite cell population of the irradiated muscle (Fig. 4I, L, M). The average number of LacZ+ cells generated per tibialis anterior muscle was 116,000 (Range: 30,000–459,000). Approximately, 10% of LacZ+ nuclei were observed peripherally within myofibers and likely represent myogenic cells that recently fused into regenerating myofibers. When the identical procedure was done with sorted satellite cells from GFP transgenic mice, large fields of GFP-expressing myofibers were observed (Fig. 4N). Thus, our sorted cells populations fulfill the functional criteria of satellite cells: self-renewal, generation of proliferating progeny, and contribution to skeletal myofiber regeneration.
GFP protein and DNA transcript quantification
To further establish the robust nature of muscle transduction a fluorescent intensity analysis was performed to determine the amount of GFP protein present in myofibers from adult C57BL/6, GFP transgenic, and mice that had received E8 IAGT (Fig. 4O). The GFP concentration in the average myofiber from an adult E8 IAGT recipient was about three-fold less than the myofiber GFP concentration observed in GFP transgenic mice. Given that the variations in GFP concentration typically observed by fluorescence-activated cell sorting (FACS) occur over several orders of magnitude, if satellite cells with a GFP concentration only three-fold less than that of GFP transgenic mice were present in samples for E8 IAGT recipients, they would have fallen well within the GFP-positive population during FACS analysis. So the lack of GFP expression in muscle from E8 IAGT recipients does not appear to be the result of insufficient GFP protein in transduced cells.
Discussion
In utero injection of lentiviral vector into the amniotic cavity targets cell populations that are in contact with amniotic fluid. Following injection, the distribution of GFP expression was relatively homogenous in all ectoderm-derived tissues (skin, CNS and peripheral nervous system) [1 –4]. This included the efficient transduction of dorsal root ganglia at all levels, so we can confirm the animal received a technically good injection. Injected animals are also born with high level GFP expression in lower limb and lower trunk muscles but without expression in upper limb muscle, diaphragm, or thoracic musculature. The technique utilized in this study transduced cells at an earlier stage of development than the chick embryo labeling studies cited above [10]. Our injections were performed at E7.5–E8 in the mouse, which corresponds to Theiler Stage 11–13 or the Downs & Davies OB-EB (no allantoic bud to early allantoic bud); LB-EHF-LHF (late allantoic bud to early head fold to late head fold) presomite stage of development to the 4–8 somite stage [15]. By ultrasound, we can tell that our injections are before Theiler Stage 14, where the body of the embryo turns. This represents a presomitic stage of development for the lower body, before dermomyotome formation, corresponding to the late stage of gastrulation (Fig. 1B). At this gestational point, only mesodermal progenitor that is exposed to the amniotic fluid that would correspond to lower body somites and exclude upper body somites is premesodermal ectoderm, or epiblast (eg, before ectoderm differentiation into myogenic progenitors in the somite). Because transduction occurred before mesoderm formation for the lower somites, our data indicate that the primary target of transduction was premyotome cells that subsequently give rise to the lower somites.
It is also clear that the transduction efficiency was very high by both protein and DNA quantification. GFP DNA quantification by Q-PCR revealed that the average myonuclei in the lower leg contained 44 copies of GFP DNA (assuming that the GFP transgenic mice had two copies per myonuclei). While the expression resulting from this high number of insertions was comparable to that seen in GFP transgenic mice, this is not unexpected since the founders of the GFP transgenic mice were highly selected for an efficient insertion site from a large number of transduced embryos. Backcrossing to the C57BL/6 background has limited the number of silent GFP insertions in the transgenic GFP mouse line and insertion site analyses have provided mixed results of one to three insertion sites (data not shown). However, more than one insertion site in the GFP transgenic mice would only indicate that our IAGT technique was even more efficient at genetically labeling cells and would bolster the results reported here. Because of the uncertainty regarding the number of GFP insertion sites, we have made the conservative assumption that only one insertion site was present in our transgenic GFP controls. Even given this assumption, DNA analysis revealed that the number of genetically marked satellite cells was ∼1 per 8,800 (using the average cellular GFP transgene load of 44 copies per cells). Given that FACS isolation is not a perfect technology and that a rare number of nonsatellite cells from dissociated muscle (many of which were GFP+) likely fell within the sorted satellite cell gate, our data indicate that it is unlikely that any satellite cells contained GFP transgenes.
Transduction of these caudal epiblast cells with lentiviral vector results in a high level of transgene expression for the lifetime of the animal. It was therefore surprising to us that we could not detect transduction of the satellite cells associated with these fibers using highly sensitive methodology. The utilization of the Myf5 nlacz mouse allowed us to discriminate the satellite cell compartment clearly by immunohistochemistry and despite counting over 1,200 B-gal-positive satellite cells in highly transduced muscle, no GFP-transduced cells were identified. This was confirmed by flow cytometric analysis of single cell suspensions of highly transduced muscles for cells co-expressing satellite cell markers and GFP, and was also confirmed by Q-PCR for transgene sequence of sorted satellite cells from transduced muscle to a sensitivity of <1%. We believe that this data strongly support our conclusion that we have transduced nascent progenitors that give rise to postnatal muscle fibers, without the associated transduction of satellite cells.
Satellite cells are a population of committed progenitor cells that are responsible for the maintenance, growth, repair, and regeneration of postnatal skeletal muscles [8]. Anatomically, satellite cells can be identified by their position beneath the basal lamina but outside the plasmalemmal surface of the muscle fiber. In addition, recent studies have identified a number of phenotypic markers that identify the majority of satellite cells. In response to injury or exercise satellite cells can self-renew, proliferate, and fuse to repair damaged fibers or form new mature muscle cells [8]. Although there may be other populations that may occasionally contribute to muscle regeneration, the majority of evidence supports the satellite cell as the primary source of muscle regeneration in the adult mouse [16].
The embryologic origin of satellite cells has been a source of some controversy. Early experiments using quail-chick chimeras suggested that satellite cells arise from the somite. More specifically, embryonic muscle development occurs in two stages. In the first stage, myocytes migrate from the border of the dermomyotome and form the primitive body muscles or the primary myotome [17,18]. In the second stage, a population of Pax3+/Pax7+ cells emerges from central dermatomyotome migrate to the primary myotome and supply further muscle progenitors including the cells that will become adult satellite cells [9 –12,19,20]. A logical assumption has been that these cells are derived from cells in the primitive streak, and that ultimately, they give rise to both the postnatal muscle fiber compartment, and satellite cells. In the context of this very convincing data, our study raises a number of interesting questions about the early events in myogenesis.
Cells streaming through the caudal aspect of the closing primitive streak have been shown in the murine embryo, using DiI labeling and LacZ transgenic cells, to colonize the paraxial mesoderm that subsequently undergoes segmentation into the posterior somites that ultimately give rise to the lower body and limb musculature [5]. Our study confirms that observation in that transduction of these cells resulted in expression of transgene in a large percentage of lower body muscle fibers in adult mice. However, our data challenge the assumption that the epiblast-derived cells traversing the primitive streak give rise to the well-documented Pax3+/Pax7+ cells in the dermomyotome that subsequently give rise to embryonic muscle and satellite cells. The dermatomyotome is likely a heterogeneous structure and we are targeting a progenitor that gives rise to muscle but not satellite cells. Our injections likely miss the cells that become the central dermatomyotome, which ultimately give rise to the satellite cell compartment.
It is important to note that studies documenting a common progenitor for embryonic muscle and satellite cells begin by analysis of the dermomyotome and have not followed the cells longitudinally into the postnatal animal. Thus, it is possible that the majority of adult muscle fibers are derived from the cells that we have labeled and that these cells do not transit through a Pax3+/Pax7+ progenitor. This is supported by the lack of early lethality of Pax7 knock out mice, which develop a relatively normal skeletal muscle compartment without the presence of satellite cells. It is also supported by observations of the appearance of satellite cells in the peripheral muscle compartment at a point in development after muscle formation. In the context of prior data supporting the Pax3+/Pax7+ common progenitor of fetal muscle and satellite cells, our data suggest an epiblast derivative with myogenic potential that gives rise to embryonic muscle, which is a separate cell population from the cells of the dermatomyotome that become the adult muscle and satellite cells.
A limitation in our study is our inability to track the very early events after transduction. There is an inherent delay in transgene expression with the lentiviral vector driven by the CMV promoter that is cell dependent. Our inability to detect any GFP in the fetal musculature until E16, 8 days after transduction, prevented any early tracking of transduced cells through the dermomyotome or early stages of muscle formation (Fig. 3K–Q). Thereafter however, GFP transgene expression remained strong in transduced muscle groups without evidence of diminution. This observation and the Q-PCR analysis argue against gene silencing as an explanation for the absence of transduced satellite cells.
One of the interesting observations of this study is that the majority of muscle fibers in transduced muscle remain strongly GFP positive for up to 2 years after transduction. The satellite cell has been regarded as the main source of myogenic activity in day-to-day maintenance of skeletal muscles [10,16,21]. In the absence of satellite cell transduction, we would have anticipated significant loss of GFP expression as time passed and the animals aged. This suggests either that an alternative population of muscle regenerative cells was transduced, or that the amount of muscle turnover during murine life is relatively small. We favor the latter interpretation. While early IAGT transduces a variety of other cell types, they are predominantly restricted to the ectoderm and neuroectoderm [4]. We have seen no transduction of bone marrow cells or perivascular cells that are the primary candidates for alternative myogenic populations. From a therapeutic perspective this data suggest that muscle correction, even in the absence of satellite cell transduction, could be sustained for a prolonged period of time. However, the translation of this finding in mice to longer-lived species and humans will require experimental confirmation.
Footnotes
Author Disclosure Statement
No competing financial interests exist.