
Abstract
Human umbilical cord blood (UCB) is rich in diverse hematopoietic stem cells that are competent to differentiate into various cell types with immunological compatibility at transplantation. Thus, UCB is a potential source for the preparation of dendritic cells (DCs) to be used for cell therapy against inflammatory disorders or cancers. However, the immunological properties of UCB-derived DCs are not fully characterized. In this study, we investigated the phenotypes and functions of UCB monocyte-derived DCs (UCB-DCs) in comparison with those of adult peripheral blood (APB) monocyte-derived DCs (APB-DCs). UCB-DCs contained less CD1a+ DCs, which is known as immunostimulatory DCs, than APB-DCs. UCB-DCs exhibited lower expression of CD80, MHC proteins, and DC-SIGN, but higher endocytic activity, than APB-DCs. Lipopolysaccharide stimulation of UCB-DCs minimally augmented the expression of maturation markers and production of interleukin (IL)-12 and tumor necrosis factor (TNF)-α, but potently expressed IL-10. When UCB-DCs were cocultured with CD14+ cell-depleted allogeneic peripheral blood mononuclear cells, they weakly induced the proliferation, surface expression of activation markers, and interferon (IFN)-γ production of T lymphocytes compared with APB-DCs. UCB possessed higher levels of prostaglandin E2 (PGE2) than APB, which might be responsible for tolerogenic phenotypes and functions of UCB-DCs. Indeed, APB-DCs prepared in the presence of PGE2 exhibited CD1a−CD14+ phenotypes with tolerogenic properties, including weak maturation, impaired IL-12 production, and negligible T lymphocyte activation as UCB-DCs did. Taken together, we suggest that UCB-DCs have tolerogenic properties, which might be due to PGE2 highly sustained in UCB.
Introduction
D
Autologous DCs from cancer patients have been tried as anti-cancer immunotherapy. However, low immunogenicity of the DCs often hinders successful treatment [6]. On the other hand, cell therapy using DCs from unrelated healthy donors may exacerbate allogeneic immune rejection as they can activate alloreactive T lymphocytes by crosspresentation of allogeneic antigens [7]. As DCs in the transplanted tissues highly express endogenous antigens combined with MHC proteins and are capable of providing costimulatory signals to recipient T lymphocytes [5], they are usually considered to be potent initiators of allogeneic immune rejection [8]. Umbilical cord blood (UCB) is a promising source of stem cells for transplantation because it shows a low incidence of GvHD [9] and contains hematopoietic progenitor cells that are competent to differentiate into various immune cells [9]. In addition, the convenience of accessibility and human leukocyte antigen (HLA) matching increases the potential for use of UCB in the transplantation. However, its application is limited to only children since the absolute number of hematopoietic stem cells in a unit of UCB is too low to be applied to adults [9]. Previous studies have demonstrated that UCB-DCs display a low level of costimulatory molecules, low responsiveness to microbial components, and low lymphocyte-activating capacities [10,11]. These tolerogenic properties of UCB-DCs lead to alleviation of immune rejection after the transplantation [12].
DCs occupy a small proportion (1%–3%) in the peripheral blood and consist of myeloid and plasmacytoid DCs [13]. Since the number of DCs is restricted, blood monocytes, which can differentiate into various DC types, are commonly used in the preparation of DCs [14]. Monocyte-derived DCs are useful models for in vitro studies, because they share many immunological properties with those of blood DCs, despite differences in the specific phenotypes and functions [15,16]. Monocytes consist of two populations, CD14+CD16− classical monocytes and CD14+CD16+ nonclassical monocytes [17], which differentiate into distinct DC subsets [18,19]. The proportion of CD14+CD16+ monocytes is significantly increased in the blood of patients with systemic inflammatory diseases such as bacterial sepsis [20]. UCB and adult peripheral blood (APB) exhibit difference in the ratio of CD14+CD16+ monocytes to CD14+CD16− monocytes. UCB displays significantly reduced frequency in CD14+CD16+ monocytes [21]. However, it is yet to be elucidated whether the lower proportions of CD14+CD16+ monocytes in UCB have a correlation with the tolerogenic immune responses of UCB-DCs and what factors are associated with the distinctive immunological features of UCB-DCs. Therefore, in the present study, we investigated immunological characteristics of UCB-DCs in comparison with those of APB-DCs with respect to their differentiation, activation, and functions.
Materials and Methods
Reagents and chemicals
Ultra-pure lipopolysaccharide (LPS) from Escherichia coli serotype K12 was purchased from InvivoGen (San Diego, CA). Ficoll-Paque PLUS was from GE Healthcare (Uppsala, Sweden). Fetal bovine serum (FBS) and RPMI 1640 media containing 2.05 mM
Preparation of human monocyte-derived DCs
APB and UCB were provided from the Red Cross and from Allcord, the public cord blood bank of Seoul Metropolitan Boramae Medical Center (Seoul, Korea), respectively. All experiments using human blood were performed under the approval of the Institutional Review Board of the Seoul National University (IRB No. S-D20060001). Both APB and UCB cells were prepared using the same anticoagulant, citrate phosphate dextrose adenine-1. APB and UCB were overlaid on Ficoll, and mononuclear cells (MNCs) were obtained by density gradient centrifugation. CD14+ monocytes were isolated using I Mag™ anti-human CD14 particles-DM (BD Biosciences) followed by magnetic separation. The isolated CD14+ cells were suspended in RPMI 1640 supplemented with 2.05 mM
Phenotypic analysis
Monocytes and DCs were washed with phosphate-buffered saline (PBS) once and then stained with fluorescence-conjugated monoclonal antibodies for 30 min on ice. The antibodies specific for human antigens that were used in the present study were CD80, CD83, CD86, MHC class I, MHC class II, CD14, CD1a, CD16, DC-SIGN, CD205, and CD206. Then the cells were washed three times with PBS containing 2% FBS and subjected to flow cytometric analysis. Ten thousand cells were acquired for each sample, and dead cells or debris were gated out. All flow cytometric data were analyzed using FACSCalibur (BD Biosciences) and FlowJo software (Tree star, San Carlos, CA).
Determination of endocytic capacity
Endocytic capacity of monocytes, unstimulated DCs, and LPS-stimulated DCs was measured using a dextran-FITC uptake assay. Briefly, the cells were washed with PBS and suspended in complete RPMI 1640 containing dextran-FITC (1 mg/mL) and incubated for 1 h at 4°C or 37°C to measure nonspecific or specific uptake, respectively. Then, the cells were washed three times with ice-cold PBS containing 2% FBS, and the geometric mean fluorescence intensity (MFI) of the cells was analyzed by flow cytometry as described above. The actual uptake (net MFI) was calculated by subtracting the MFI at 4°C from the MFI at 37°C.
Preparation of heat-killed E. coli
E. coli BL21 (DE3) was obtained from Stratagene (La Jolla, CA). Bacteria were cultured in LB broth media at 37°C until mid-log phase and harvested by centrifugation. The bacterial cells were suspended in PBS and killed by heating at 60°C for 1 h. To confirm complete killing, heat-treated bacteria were plated onto LB agar plates and cultured overnight at 37°C. No bacterial colony was observed (data not shown).
Quantification of cytokines
DCs (5 × 105 cells/mL) were stimulated with E. coli LPS (100 ng/mL) for 48 h, and the amount of IL-12p40, tumor necrosis factor (TNF)-α, and IL-10 in the cell culture supernatant was measured by enzyme-linked immunosorbent assay (ELISA) (R&D Systems). DCs were stimulated with heat-killed E. coli (HKEC) (1 × 106 CFU/mL) for 24 h, and the concentration of IL-12p70 was analyzed by ELISA (BioLegend). All procedures were conducted according to the manufacturer's instructions.
Analysis of T lymphocyte proliferation and activation
DCs (2.5 × 105 cells/mL) were treated with HKEC (1 × 106 CFU/mL) for 24 h and washed with PBS. Unstimulated and HKEC-stimulated DCs (5 × 104 cells) were cocultured with CFSE-labeled allogeneic peripheral blood MNCs (PBMC, 1 × 106 cells) for 3–5 days. On day 3, the culture supernatant was collected and subjected to ELISA for measuring the amount of interferon (IFN)-γ and IL-10 (R&D Systems). On day 5, CFSE-labeled PBMCs were stained with anti-human CD3 antibodies to detect T lymphocytes among the PBMCs, and proliferation of CD3+ cells and CD3− cells was analyzed by flow cytometry. To examine the expression of activation markers on T lymphocytes, PBMCs were stained with anti-human CD3 antibodies, anti-human CD25 antibodies, and anti-human HLA-DR, DP, DQ antibodies, followed by flow cytometry as described above.
Prostaglandin E2 quantification
To obtain plasma, heparinized UCB and APB were centrifuged at 1,320× g at 4°C for 10 min. The plasma was used to determine PGE2 levels using a PGE2 Competitive ELISA Kit from Arbor Assays (Ann Arbor, MI) according to the manufacturer's instruction.
Statistical analysis
Experimental data were compared using Student's t-test or ANOVA test (Tukey's multiple comparison test). P values under 0.05 were considered statistically significant. Results are indicated as mean ± standard error of the mean (SEM).
Results
UCB-derived monocytes successfully differentiate into immature DCs
Enrichment of pure and functional monocytes is crucial for successful differentiation into DCs. In this study, we examined purity of UCB- and APB-derived CD14+ monocytes before and after the magnetic separation. Flow cytometric analyses showed that UCB exhibited significantly lower frequencies of CD14+ cells than APB (Fig. 1A). After magnetic separation, the enriched CD14+ cells from both sources displayed 96%–99% purities on average, but the expression of CD14 on the UCB-derived monocytes was lower than that of APB-derived monocytes (Fig. 1B). When the monocytes differentiated into immature DCs, UCB-DCs displayed similar morphologies to those of APB-DCs (Fig. 1C, F). In addition, both UCB- and APB-DCs showed significant changes in the phenotypes and endocytic abilities upon differentiation. Although UCB-DCs exhibited CD14+ and CD1a− phenotypes, unlike CD14− and CD1a+ APB-DCs, more than 88% of the cells expressed DC-SIGN and PD-L1, which are other indicative markers for DCs (Fig. 1D, G). Furthermore, UCB-DCs and APB-DCs displayed enhanced endocytic abilities compared to their precursors (Fig. 1E, H). These results suggest that highly pure monocytes isolated from UCB successfully differentiated into immature DCs.
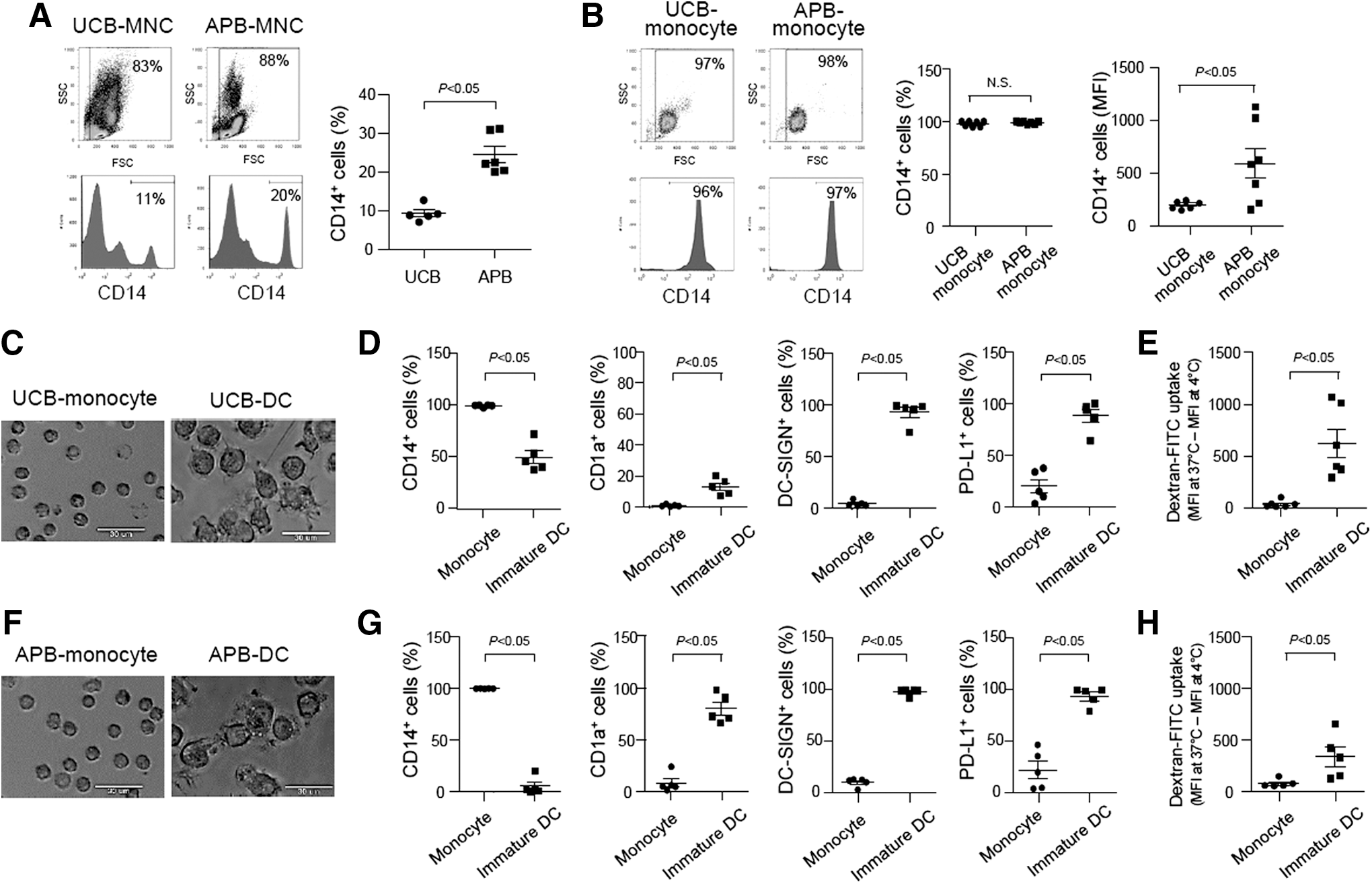
Umbilical cord blood (UCB)-derived monocytes successfully differentiate into immature dendritic cells (DCs). Mononuclear cells (MNCs) were isolated from UCB and adult peripheral blood (APB), respectively, by density gradient centrifugation.
Immature UCB-DCs display distinctive features from immature APB-DCs in phenotypes and endocytic capacity
Next, we comparatively examined the expression of phenotypic markers and endocytic activity of immature DCs differentiated from UCB- or APB-derived monocytes. Cell populations gated for the phenotypic and functional analyses were presented in the upper panel of Fig. 2A. When the monocytes were differentiated into immature DCs, UCB-DCs displayed less CD1a+ cells, but more CD14+ cells than APB-DCs (Fig. 2A, lower panel, B). In addition, UCB-DCs showed significantly lower levels of CD80, MHC class I and II, and DC-SIGN than APB-DCs, whereas the expression of CD86, CD205, and CD206 was not significantly different from those of APB-DCs (Fig. 2C–I). UCB-DCs displayed better endocytic activity than APB-DCs (Fig. 2J). These results indicate that immature UCB-DCs exhibited lower expression of CD1a, CD80, MHC proteins, and DC-SIGN, but higher CD14 expression and endocytic capacity than immature APB-DCs.
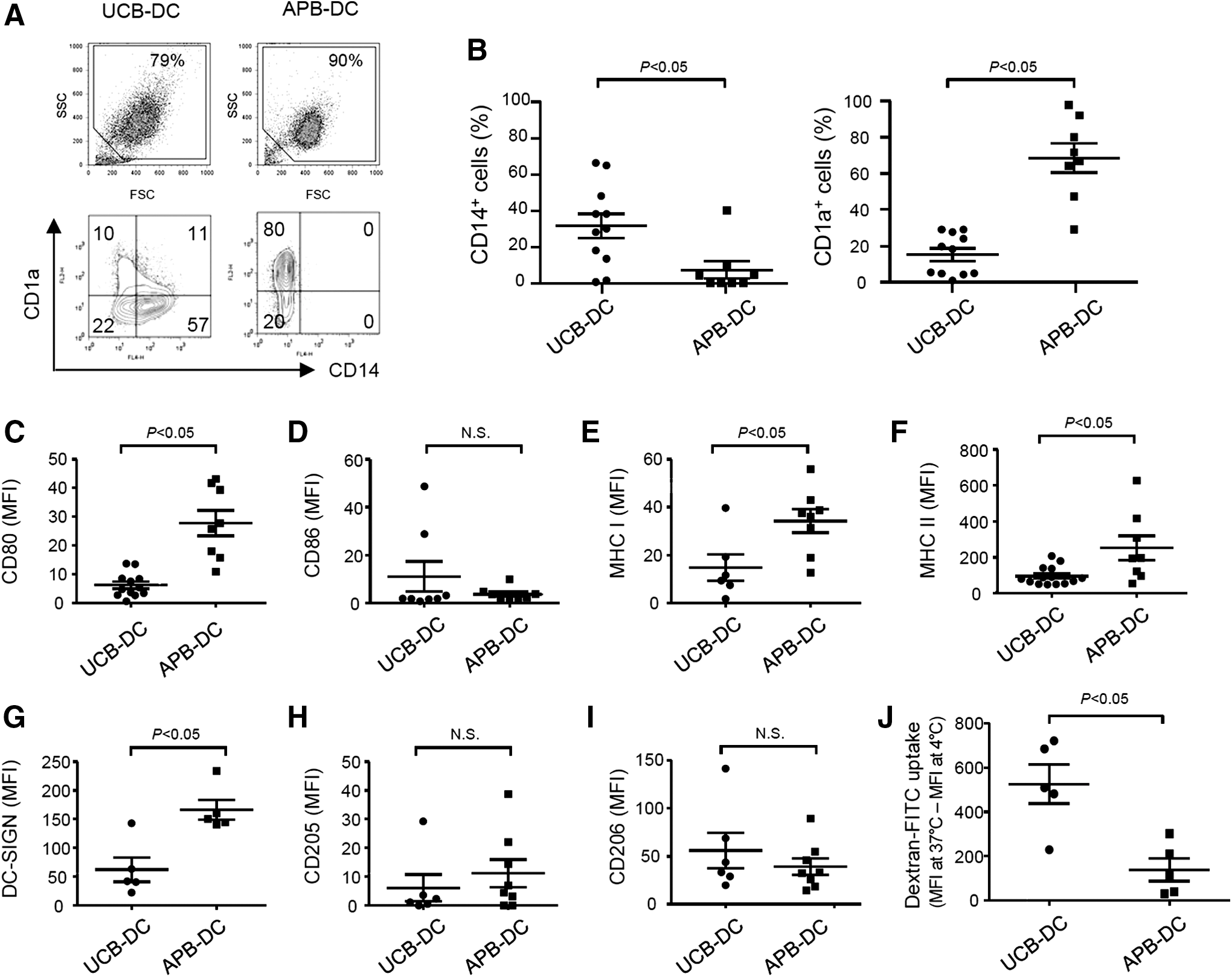
UCB-DCs display distinctive features from APB-DCs in phenotypes and endocytic capacity. CD14+ monocytes (2 × 106 cells/mL) isolated from UCB and APB were differentiated into DCs in the presence of human GM-CSF (800 U/mL) and IL-4 (500 U/mL) for 6 days.
LPS weakly induces the maturation of UCB-DCs and their expression of inflammatory cytokines in comparison with those of APB-DCs
Since maturation of DCs is an important process in the mediation of immune responses, we examined the phenotypic changes and cytokine expression of DCs. Stimulation with LPS hardly or slightly induced costimulatory molecules, including CD80, CD83, and CD86 and MHC class I and II on UCB-DCs, whereas APB-DCs exhibited a remarkable increase in the expression of these molecules (Fig. 3A, B). UCB-DCs still sustained high endocytic activity even after LPS stimulation, while LPS-matured APB-DCs showed remarkably decreased endocytic activity (Fig. 3C). The expression of phagocytosis-related receptors (CD205 and CD206) was minimally changed in LPS-treated UCB-DCs (Fig. 3D). UCB-DCs produced significantly lower IL-12p40 and TNF-α, but higher IL-10 in response to LPS than APB-DCs did (Fig. 3E). HKEC-stimulated UCB-DCs hardly induced IL-12p70 in comparison with HKEC-stimulated APB-DCs (Fig. 3F). Taken together, these results indicate that UCB-DCs weakly elicit phenotypic and functional maturation in response to LPS or HKEC.

LPS weakly induces the maturation of UCB-DCs and their expression of inflammatory cytokines in comparison with those of APB-DCs.
UCB-DCs weakly induce proliferation and activation of allogeneic T lymphocytes
One of the major functions of DCs is to stimulate naive T lymphocytes to differentiate into various effector T cells [22]. To examine T lymphocyte-activating capacities of DCs, unstimulated or HKEC-stimulated DCs were cocultured with CD14+ cell-depleted allogeneic PBMCs. HKEC-stimulated UCB-DCs induced less proliferation of CD3+ T lymphocytes in PBMCs than those of APB-DCs (Fig. 4A, B). Interestingly, APB-DCs stimulated with HKEC markedly augmented the proliferation of CD3− cells. Given the results that most of the proliferated CD3− cells expressed CD56 (data not shown), APB-DCs appear to preferentially induce proliferation and activation of CD56+ natural killer cells besides CD3+ T lymphocytes. Concomitantly, UCB-DCs poorly induced CD25+ MHC class II+ T lymphocytes, whereas APB-DCs increased those cell populations (Fig. 4C, D). In addition, HKEC-stimulated UCB-DCs weakly induced IFN-γ production (Fig. 4E), whereas that of HKEC-stimulated APB-DCs by PBMCs was potently induced. However, the level of IL-10 in the supernatants of PBMCs cocultured with HKEC-stimulated DCs from both sources was not significantly different (Fig. 4F) although slightly more potent IL-10-producing capacities were observed in UCB-DCs than in APB-DCs (Fig. 3E). These results indicate that UCB-DCs weakly induced activation of allogeneic T lymphocytes in comparison with APB-DCs.
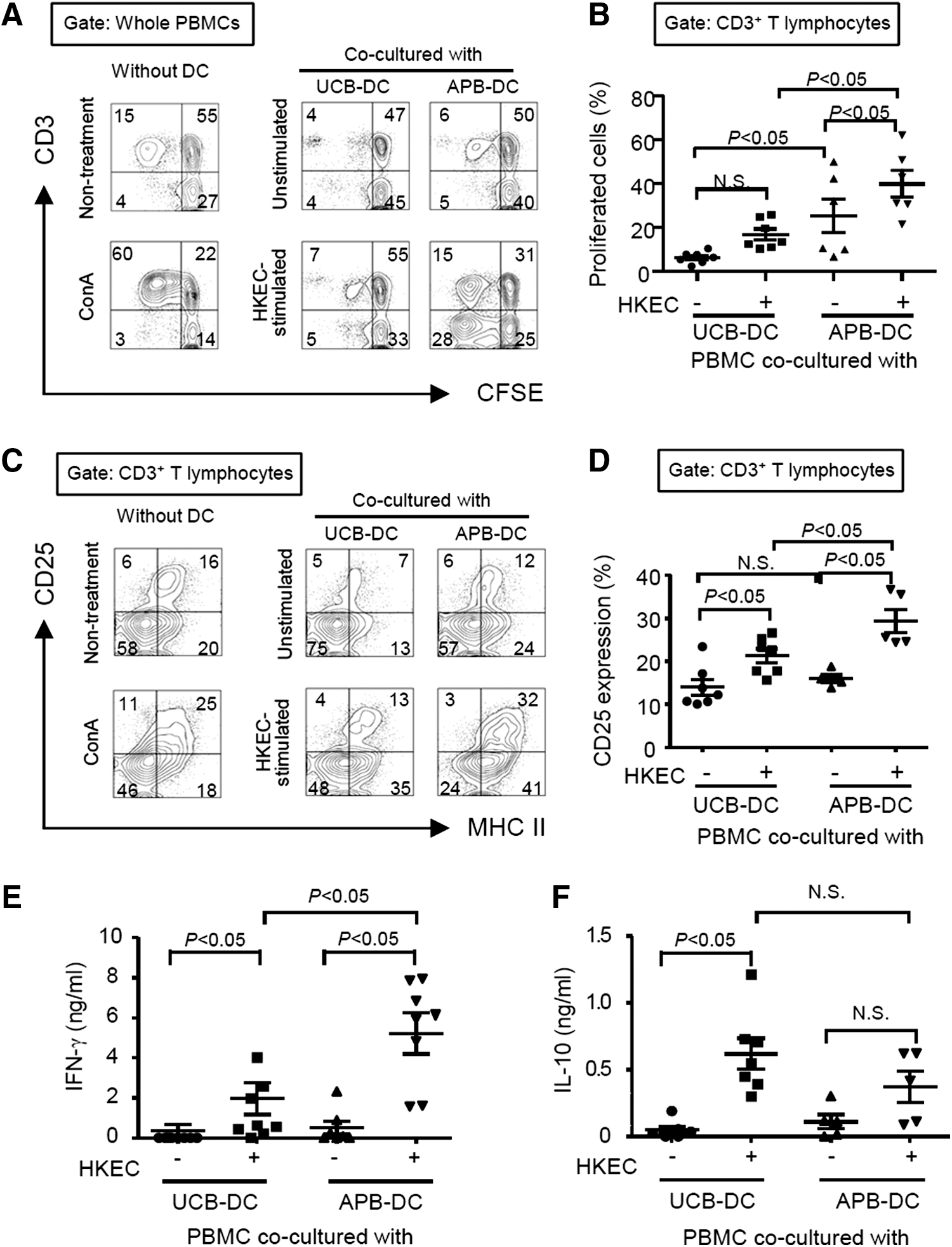
UCB-DCs weakly induce proliferation and activation of T lymphocytes. DCs (5 × 104 cells) were stimulated with HKEC (1 × 106 CFU/mL) for 24 h, and the allogeneic peripheral blood MNCs (PBMCs) (5 × 105 cells) were cocultured with the stimulated DCs for 3–5 days.
Reduced frequency of CD14+CD16+ monocytes in UCB correlates with increased level of blood PGE2
CD14+CD16− and CD14+CD16+ monocytes exhibit different immunological properties when they differentiate into DCs [19]. To unravel the mechanism responsible for the distinct phenotypes and functions of UCB-DCs, monocyte subtypes of UCB and APB were analyzed. Frequency of CD14+CD16+ monocytes was significantly lower in UCB than in APB (Fig. 5A). PGE2 is a potent immunomodulator that regulates differentiation and activation of various immune cells and is known to exist at a high level in UCB [23]. Thus, we measured PGE2 concentration in the plasma of UCB and APB and examined whether it was related to the frequencies of CD14+CD16+ monocytes. Higher amounts of PGE2 were detected in the plasma of UCB than that of APB (Fig. 5B). It was ensured that PGE2 was not additionally synthesized during the plasma preparation process even in the absence of cyclooxygenase inhibitors (data not shown). The PGE2 level of UCB plasma was inversely correlated with the percentage of CD14+CD16+ monocytes (R 2 = 0.8) (Fig. 5C), but a significant correlation between two parameters was not observed in APB (data not shown). To examine whether PGE2 is directly involved in the regulation of CD14+CD16+ monocyte differentiation, CD14+ monocytes isolated from APB were cultured with or without PGE2. An increase of CD14+CD16+ cells was observed in the monocytes cultured in the absence of PGE2 (Fig. 5D), which is concordant with the previous report [24]. In contrast, CD14+CD16+ cells were less augmented in the monocytes cultured in the presence of PGE2 (Fig. 5D). Furthermore, CD14+ monocytes enriched from UCB-derived MNCs showed lower levels of E-prostanoid (EP) 2 and EP4 than those of APB (Fig. 5E, F). These results suggest that the reduced frequency of CD14+ CD16+ monocyte in the UCB is associated with the levels of PGE2 in the plasma.
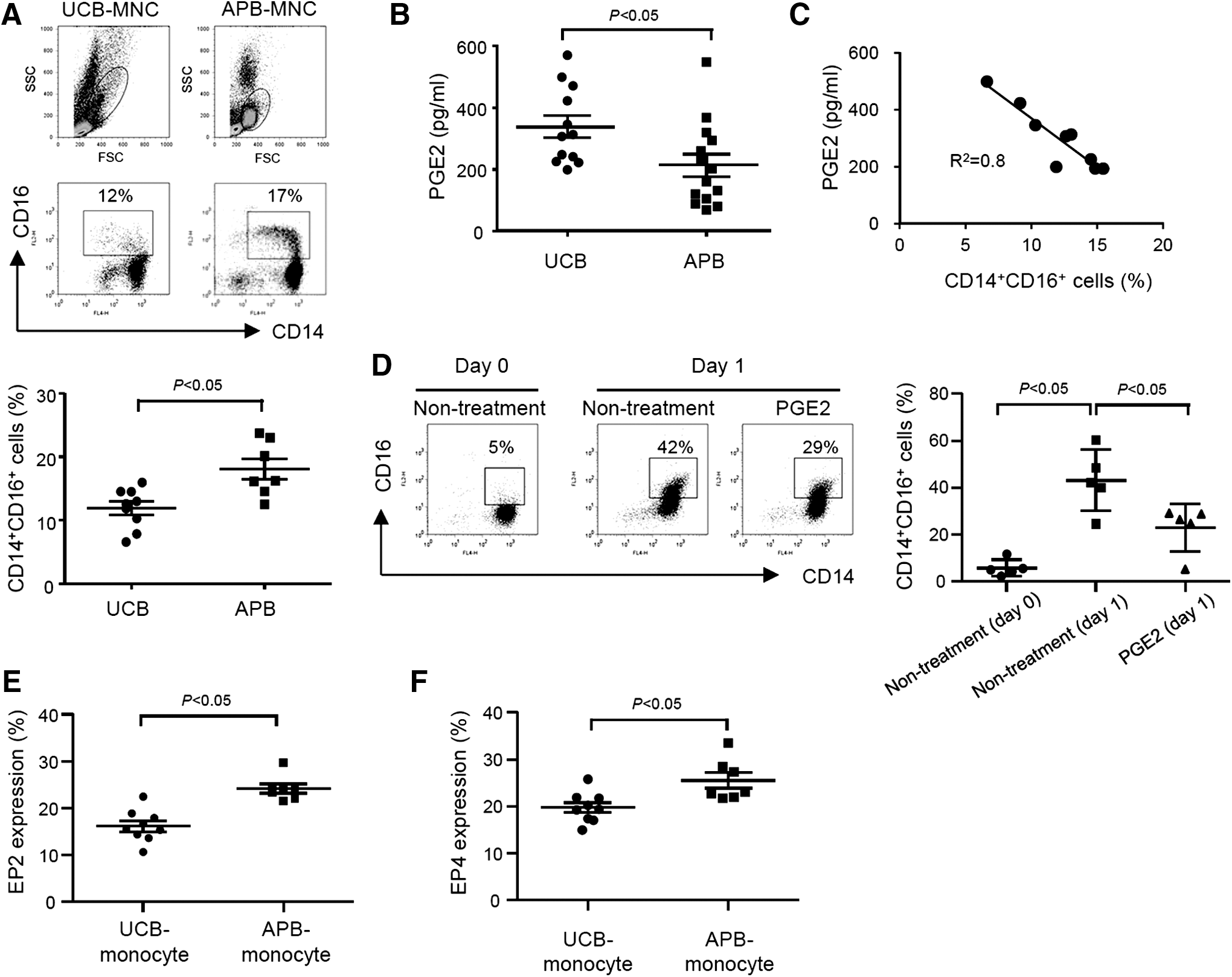
Reduced frequencies of CD14+CD16+ monocytes in UCB correlate with increased level of blood prostaglandin E2 (PGE2).
PGE2 induces differentiation of APB monocytes into UCB-DC-like cells
To further investigate the role of PGE2 in phenotypic and functional characteristics of DCs, APB monocytes were differentiated into immature DCs in the absence (control-DCs) or presence of PGE2 (PGE2-DCs). PGE2-DCs preferentially expressed CD14 rather than CD1a, whereas control-DCs expressed CD1a, but not CD14 (Fig. 6A). PGE2-DCs displayed higher endocytic activity than the control-DCs (Fig. 6B). Unlike control-DCs, PGE2-DCs weakly augmented the expression of costimulatory molecules in response to LPS (Fig. 6C). PGE2-DCs expressed low IL-12p70, but remarkably high IL-10 upon stimulation with LPS. In contrast, the LPS-treated control-DCs efficiently upregulated IL-12p70, but weakly induced IL-10 (Fig. 6D). HKEC-stimulated PGE2-DCs poorly induced allogeneic T lymphocyte proliferation when compared to that of control-DCs (Fig. 6E). Furthermore, less IFN-γ, but more IL-10 was detected in the allogeneic PBMCs cocultured with PGE2-DCs than in PBMCs cocultured with control-DCs (Fig. 6F). These results show that monocytes exposed to PGE2 differentiate into DCs similar to UCB-DCs.

PGE2 induces differentiation of monocytes into UCB-DC-like cells. Monocytes (2 × 106 cells) isolated from APB were differentiated into immature DCs in the absence or presence of exogenous PGE2 (0.1, 1, 10, or 100 nM) for 6 days.
Discussion
In the present study, we investigated immunological characteristics of UCB-DCs. UCB-DCs exhibited unique phenotypes and functions as distinct from those of APB-DCs. UCB-DCs expressed lower CD1a, CD80, MHC proteins, and DC-SIGN than APB-DCs and minimally induced maturation markers and inflammatory cytokines. Moreover, UCB-DCs elicited weaker T cell proliferation and activation than APB-DCs. All of these results are in keeping with those of other groups [23,25,26] and are also coincident with the immunological characteristics of in vitro-generated tolerogenic DCs [27 –29]. The phenotypes and functions of UCB-DCs seem to have relevance to PGE2 level in the plasma in that (i) the UCB monocytes were exposed to a higher level of PGE2 than the APB monocytes, (ii) the higher level of PGE2 was correlated with reduced frequency of CD14+CD16+ monocytes in UCB, and (iii) APB-DCs differentiated in the presence of PGE2 acquired similar phenotypes and functions with those of UCB-DCs.
CD1a+ DC is a typical immunostimulatory DC subtype that produces a high level of IL-12 and has a potent lymphocyte-activating capacity, while CD1a− DC displays tolerogenic functions [30 –32]. Particularly, a high endocytic ability is a distinctive property of CD1a− DCs and it more efficiently takes up dextran, lipid molecules, and formalin-fixed bacteria than CD1a+ DCs [32]. In this study, we found that UCB-DCs mainly consisted of CD1a− DCs and APB-DCs prepared in the presence of PGE2 showed the decreased CD1a expression. In addition, UCB-DCs containing high number of CD1a− cells exhibited higher endocytic capacity than APB-DCs. Furthermore, the APB-DCs differentiated in the presence of PGE2 showed potentiated endocytic ability as well compared to control-DCs differentiated in the absence of PGE2. Thus, the enhanced endocytic ability seems to be a common feature of CD1a− DCs, which were rich in UCB-DCs.
UCB-DCs weakly expressed the maturation markers and inflammatory cytokines in response to LPS or HKEC that are crucial for the activation of subsequent adaptive immunity. These results are in agreement with the previous report that UCB-DCs showed lower expression level of costimulatory molecules, MHC proteins, and IL-12p70 in response to LPS or dying cells, including apoptotic cells and necrotic cells [26]. Therefore, UCB-DCs might be less potent than APB-DCs in the induction of adaptive immunity due to inadequate provision of costimulatory signals and immunostimulatory cytokines to T lymphocytes necessary for the generation of effector T lymphocytes. Indeed, we observed that UCB-DCs weakly induced the proliferation and activation of T lymphocytes at the coculture of UCB-DCs and allogeneic PBMCs. All these properties of UCB-DCs seem to be associated with those of CD1a− DCs, in light of the fact that CD1a− DCs even in response to LPS displayed a decrease in the expression of CD83, CD86, and IL-12p70 and activation of T lymphocytes [32,33].
Differentiation into CD1a+ DCs or CD1a− DCs appears to be differently regulated depending on the types of precursors and microenvironment. For example, unlike our findings, some of the previous studies have demonstrated that CD14+CD16+ monocytes differentiate into CD1a− DCs exhibiting low immunostimulatory potencies, compared to CD1a+ DCs differentiated from CD14+CD16− monocytes [18,19]. In addition, monocytes consisting of mostly CD14+CD16− cells could differentiate into CD1a− DCs upon exposure to inflammatory mediators such as microbial molecular patterns [34] and phospholipid metabolites, including PGE2 [29,32]. In keeping with the previous evidence, we have also shown that monocytes stimulated with PGE2 differentiate into CD1a− DCs. However, under the present experimental condition, PGE2 was not able to induce CD14+CD16+ monocytes as the precursor of CD1a− DCs, because PGE2 treatment suppressed the in vitro culture-induced CD16 on the CD14+CD16− monocytes. Specific mechanisms by which PGE2 modulates immunological characteristics of DCs should be further studied.
PGE2 levels are highly sustained in the human placenta and UCB to maintain pregnancy and help in uterine contractions during parturition [35]. Considering that PGE2 levels soar transiently in both maternal and fetal circulation during periparturition [36], exposure of immune cells and their precursors to high amounts of PGE2 seems to be inevitable. Cells exposed to PGE2 have been reported to desensitize and downregulate the expression of EP receptors [37,38]. In the present study, we found that UCB-derived monocytes expressed significantly lower levels of EP2 and EP4 than APB-derived monocytes, which may be due to PGE2-rich environment of UCB. Additionally, it was observed that the precursors of UCB-DCs were exposed to high concentrations of PGE2, which would enhance tolerogenic properties during differentiation. Remarkably, PGE2 is likely to differentially regulate immune responses of DCs depending on the differentiation stage of the DCs. At the early phase of differentiation, PGE2 suppresses differentiation of monocytes into inflammatory DCs, inducing Th1 cell responses [39]. In contrast, the presence of PGE2 at the late phase of differentiation promotes final maturation of DCs [40]. Therefore, exposure of DC precursors to PGE2 at an early differentiation stage might be critical in the determination of immunological properties of UCB-DCs.
UCB contained a lower number of CD14+CD16+ monocytes, which are known as activated monocytes than APB. Concomitant with our findings, reduction of inflammatory CD14+CD16+ monocytes in UCB and its implications for weak immune responses of newborns to infections has been demonstrated [21]. In this study, we observed that PGE2 level in the UCB plasma has inverse correlation with the frequencies of CD14+CD16+ cells. Indeed, treatment with PGE2 suppressed the differentiation of CD14+CD16− monocytes into CD14+CD16+ monocytes. In addition, PGE2 has been previously reported to suppress activation and differentiation of T and B lymphocytes [41,42]. Taken together, these data suggest that sustaining high PGE2 levels in UCB might be a plausible strategy for protecting the immunocompromised neonates from excessive inflammation. Conclusively, UCB-DCs would be worthy of being developed into immune cell therapeutics against chronic inflammatory disorders, including GvHD and autoimmune diseases.
Footnotes
Acknowledgments
This work was supported by grants from the National Research Foundation of Korea funded by the Korean government (MISIP) (2010-0029116) and the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare (HI14C0469), Republic of Korea. Umbilical cord blood was kindly provided from Allcord, SMG-SNU Boramae Medical Center.
Author Disclosure Statement
No competing financial interests exist.