
Abstract
Mesenchymal stem cells (MSC) possess a range of immunomodulatory properties which they exert through soluble mediators and through direct cell–cell contact. Due to these immune regulatory properties, the safety and clinical efficacy of MSC treatment has been tested in a number of autoimmune disorders. In this review we analyze the current data from early phase trials into Crohn's disease, systemic lupus erythematosus, and rheumatoid arthritis. In general, no adverse side effects were observed in patients treated with MSC; however, their clinical efficacy is difficult to interpret. Systemic or site-specific administration of MSC has been reported to exert potent immunomodulatory effects in 7 of the 11 trials discussed. Nonetheless, the mechanism(s) by which MSC exert their regulatory effects in vivo remain largely unknown. We discuss potential limitations or safety concerns associated with MSC therapy, including the heterogeneity of MSC and their potential contribution to disease pathogenesis, which need to be considered when designing future clinical trials, along with the need to standardize trial design. Although we are bridging the translational gap between scientific observations on MSC function and clinical applications for therapy, our understanding of basic MSC biology is still limited. Despite these issues, large, double-blinded, multicenter clinical trials are already underway. Further research into the endogenous function of MSC is required to elucidate the mechanism by which therapeutic MSC are acting.
Introduction
M
Immunomodulatory Properties of MSC
The immunomodulatory effects of MSC have been extensively studied using bone marrow-derived (BM)MSC [4]. Recent studies indicate that MSC isolated from other, more accessible, tissue sources (eg, human umbilical cord, adipose tissue, and dental pulp) also have the capacity to regulate immune cell function, although to varying degrees [6]. Evidence suggests that they mediate their effects through the production of soluble anti-inflammatory mediators such as prostaglandin E2 (PGE2), indoleamine 2, 3-dioxygenase (IDO), transforming growth factor beta (TGFβ), and interleukin-6 (IL-6) [6]. Some of the literature reviewed below highlights why we have rapidly advanced to using MSC therapy in clinical trials. However, it remains unclear whether observations made using ex vivo-expanded MSC represent the true function of tissue-resident MSC in vivo.
MSC and innate immunity
MSC have been reported to dampen innate immune responses by suppressing the effector functions of neutrophils and natural killer (NK) cells, as well as skewing the differentiation of monocytes toward a more anti-inflammatory phenotype (M2) [4]. For example, resting human BMMSC inhibited reactive oxygen species production and apoptosis of fMLP-treated neutrophils [7]. They also suppressed interferon (IFN)γ secretion and cytotoxic activities of IL-2-activated, but not resting, NK cells in vitro [8]. Suppression of NK cell function required activation of the cells and was mediated by direct NK cell–MSC contact and PGE2 [8]. Murine and human BMMSC also promoted monocyte polarization to IL-10-producing M2 macrophages, once again in a cell contact and soluble mediator (IDO and PGE2)-dependent manner [9,10]. Indeed, IL-10 produced by M2 macrophages reduced neutrophil infiltration and lethality of sepsis in vivo following infusion of BMMSC [10]. Thus, MSC appear to have the ability to turn off inflammatory responses, but may also coordinate the switch from innate to adaptive immunity during protective inflammation.
A recent study showed that toll-like receptor (TLR)-3-stimulated human BMMSC increased neutrophil respiratory burst capabilities to a greater extent than resting BMMSC, indicating that MSC also have the ability to enhance effector functions in certain contexts [11]. Human MSC isolated from thymus, spleen, and adipose tissue also exhibited these immunomodulatory effects in response to poly(I:C)-induced TLR stimulation [11]. In addition to modulating effector functions, MSC have also been reported to produce survival factors that enhance the longevity of leukocytes within tissues. The presence of human BMMSC delayed resting and IL-8-stimulated neutrophils from undergoing apoptosis at 18 and 40 h through the release of IL-6, IFNβ, and granulocyte-macrophage colony-stimulating factor, without the requirement for cell–cell contact [11]. To date, this effect has not been reported for MSC from other sources. One could suggest that such regulatory mechanisms enable MSC to prolong the lifespan of neutrophils within the BM niche, allowing large quantities of cells to be released when a pathogen is encountered [7]. However, what direct effects MSC have on innate immunity in vivo and whether these are context- and/or stimulus-specific or comparable to those described in vitro requires further investigation.
MSC and adaptive immune responses
The ability of BMMSC to suppress T cell responses, in terms of proliferative capacity or effector function, has been well documented in both murine and human systems [12]. These types of assays now represent a standard means to test and confirm the potency of MSC following isolation or FACS sorting into a homogeneous population. Numerous studies have shown that human BMMSC can promote the survival of T cells, while maintaining them in a quiescent state by suppressing proliferation [12] and the production of proinflammatory cytokines (eg, IFNγ [4]). In a recent study, adipose, dental pulp, and umbilical cord-derived MSC were shown to be capable of suppressing T cell proliferation and effector functions to the same extent as that reported for BMMSC [6]. MSC mediate their effects on T cells through at least two mechanisms: release of soluble mediators such as PGE2, TGFβ, and IDO [13], and direct cell–cell contact [12]. The reasons why certain circumstances require cell–cell contact in addition to soluble mediators remain unknown and require further elucidation. BMMSC also promoted the expansion of the TReg cell population in purified peripheral blood mononuclear cells [14]. The ability of MSC to maintain T cells in a quiescent state either directly or through TReg cells may be indicative of their endogenous functions within the BM niche [12,14]. It may also explain, in part, the beneficial effects of MSC infusion in preclinical models of graft versus host disease [14].
MSC influence leukocyte recruitment during inflammation
As well as the direct effects of MSC on immune cell functions, we have previously shown that MSC can indirectly regulate the recruitment of circulating leukocytes through interactions with vascular endothelium [3]. In a MSC–endothelial cell in vitro coculture model, human BMMSC or umbilical cord-derived MSC potently downregulated endothelial responses to cytokine stimulation (ie, TNFα [15]). This reduced the ability of endothelial cells to support binding of purified neutrophils and lymphocytes from flow, in an IL-6 and TGFβ-dependent manner [3]. Fetal umbilical cord-derived MSC had a greater suppressive effect than adult BMMSC on TNFα-induced endothelial recruitment of leukocytes [15]. These findings support the concept that MSC may act as endogenous regulators of inflammation, but also highlights that donor age may influence the effectiveness of MSC-mediated immunosuppression. This, in itself, could have significant impact for clinical intervention.
Impact of environment on MSC function
Priming MSC with proinflammatory cytokines can enhance their immunomodulatory effects by stimulating the release of regulatory mediators such as IDO and PGE2 [16]. Cytokine stimulation was shown to differentially alter the immunomodulatory properties of human BMMSC and umbilical cord-derived MSC [16]. Cord-derived MSC had a greater suppressive effect than BMMSC when primed with IFNγ as assessed by T cell proliferation and mixed lymphocyte reactions in vitro [16]. That said, priming itself is not essential for the suppressive actions of MSC [4,7,12], but such data suggest that the MSC can respond to their local microenvironment, which in turn could affect their behavior. For example, MSC suppression of leukocyte recruitment required crosstalk between MSC and endothelial cells; in the absence of this cross-talk the suppressive effects were lost [15]. Moreover, other stromal populations, such as fibroblasts and smooth muscle cells, are known to change their phenotype from an endogenous regulatory state to a proinflammatory stimulatory state in chronic inflammatory diseases [17]. Whether priming of MSC in vitro is representative of the in vivo situation requires further research. However, existing evidence strongly suggest that MSC alter their effector functions upon changes in their environment, whether that be cytokines in the inflammatory milieu or through direct contact with the target cell [16,18]. Such plasticity could have implications for MSC as a cell therapy—can we guarantee that the cells administered therapeutically will maintain their immunosuppressive effects in a chronically inflamed site?
In the last 5 years, studies have indicated that MSC can be polarized toward a proinflammatory (MSC1) or an anti-inflammatory (MSC2) phenotype as has been reported for monocytes/macrophages [18 –22]. The terms MSC1 and MSC2 refer to the phenotype of the MSC following TLR activation, rather than cells derived from different origins. Initial reports suggested that engagement of TLR-3 in vitro enhanced the immunosuppressive effects of BMMSC by inducing the release of anti-inflammatory factors (such as IL-10, IDO, and PGE2) and were defined as acquiring a MSC2 phenotype [18]. In contrast, TLR-4 activation of BMMSC abrogated their ability to suppress T cell proliferation [18]. Indeed, these cells released proinflammatory cytokines (eg, TNFα) and deposited collagen, leading to them being defined as acquiring an MSC1 phenotype [18]. Furthermore in a murine model of lung injury, systemic administration of TLR-4-stimulated MSC exacerbated the disease, while treatment with TLR-3-stimulated MSC suppressed it compared to infusion of naive BMMSC [18]. Subsequent in vitro studies presented conflicting findings: IDO or PGE2 secretion and T cell proliferation have been reported to be enhanced, reduced, or unchanged by TLR-3 and TLR-4-stimulated BMMSC [19 –21]. Moreover, TLR-3 and TLR-4 stimulation did not alter the immunosuppressive capabilities of human adipose and umbilical cord-derived MSC, but did alter dental pulp-derived MSC properties, such that they now augmented T cell proliferation when compared to untreated controls [21]. Contradictory outcomes are likely to be due to differences in the MSC source and experimental conditions, such as the duration of treatment with TLR agonists and the number of MSC used. Furthermore, MSC infusion has previously been shown to have beneficial effects in murine models of sepsis where high levels of lipopolysaccharide (a TLR-4 ligand) are present [22]. The MSC did not polarize to a proinflammatory (so called MSC1) phenotype and augment inflammation, but instead dampened it. This suggests that the effects of TLR priming observed in vitro may not be representative of the MSC phenotype in vivo. Moreover, it appears that MSC are able to not only respond to cytokines (as discussed above), but also to other danger signals and bacterial components which have the potential to influence their immune regulatory effects [18]. Significantly, more research is required to understand the impact of the local microenvironment on MSC function and to clearly assess what conditions are suitable for MSC therapy. Indeed this should be fully appreciated before using these cells therapeutically.
MSC in the Treatment of Autoimmune Disease
Early studies have shown that MSC therapy suppressed graft versus host disease forming the basis for 29 ongoing trials in this area [5]. Currently ∼350 trials focusing on the reparative capabilities of MSC therapy, particularly in cardiac syndromes and spinal cord injury, have either been completed or are in progress [5]. Another 49 trials are examining the efficacy of MSC in autoimmune diseases, with a significant number involving patients with Crohn's disease, SLE, and RA (Table 1) [5]. In this study, we have evaluated the evidence from preclinical in vivo models and the findings from the few completed clinical trials in these diseases. The mechanism by which MSC are exerting their effects in these studies is still unknown, but it is likely to be due to their effects on cellular components of the innate and adaptive immune system (Fig. 1). One caveat with evaluating the efficacy of MSC treatment in autoimmune disorders is that appropriate placebo controls were not performed in many of the current phase I/II studies [5]. Another point to consider is that autoimmune diseases have waves of activity where symptoms worsen and improve cyclically. As such, it is difficult to ascertain whether any improvements in disease outcome after MSC administration are directly due to MSC treatment or the natural history of the disorder. Limitations in trial design, route of administration, and MSC source that could influence the interpretation of the data from these trials are further discussed in the last section. Overall, clinical efficacy of MSC therapy has only been consistently reported in SLE, while the majority of outcomes from larger trials are expected in 2015–16. A summary of the 13 current studies in Crohn's disease, SLE, and RA is shown in Table 1.
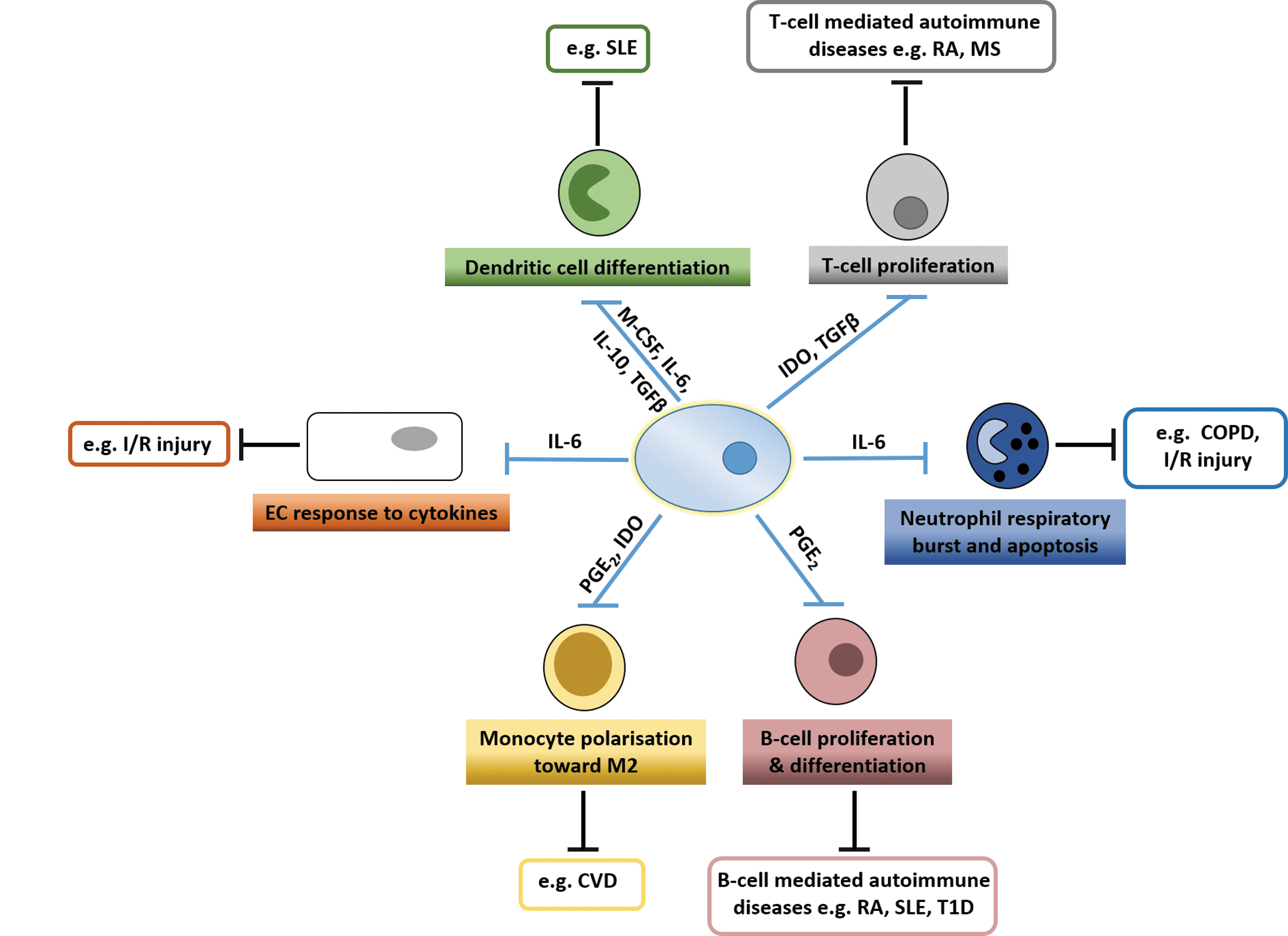
Immunomodulatory effects of MSC and their potential therapeutic action in chronic inflammatory and autoimmune diseases. MSC can directly influence the fate and function of many distinct leukocyte populations primarily through the action of soluble mediators. Numerous studies have shown that MSC can suppress the proliferation and differentiation of B-cells and T cells, as well as dendritic cells. They can also inhibit neutrophil apoptosis and respiratory burst by inhibiting production of reactive oxygen species. Additionally, MSC promote polarization of monocytes toward an M2 phenotype and alter the response of endothelial cells to proinflammatory cytokines. While MSC have been shown to have these effector functions, their exact mode of action in autoimmune diseases is unknown. Since many of these leukocyte and endothelial functions have been implicated in chronic inflammatory disorders and autoimmune diseases (highlighted in the boxes), we propose that MSC may have beneficial effects by suppressing these effector functions. SLE, systemic lupus erythematosus; RA, rheumatoid arthritis; T1D, type1 diabetes; I/R injury, ischemia/reperfusion injury; MS, multiple sclerosis; COPD, chronic obstructive pulmonary disorder; CVD, cardiovascular disease; MSC, mesenchymal stem cells.
Reference [5]
Studies that are either completed, but the data have not been published, or their status is unknown.
Studies that have appropriate placebo controls.
BMMSC, bone marrow-derived MSC; UCMSC, umbilical cord-derived MSC; ADMSC, adipose-derived MSC; MSC-AFP, fistula plug MSC; N.S., not specified; Trial ID, clinicaltrials.gov identifier number; MSC, mesenchymal stem cells; SLE, systemic lupus erythematosus; RA, rheumatoid arthritis.
Crohn's disease
Crohn's disease is a form of inflammatory bowel disorder associated with elevated levels of proinflammatory cytokines, aberrant activation of leukocytes, and fistulization [23]. In a preclinical murine colitis model, a single intraperitoneal infusion of 105–106 adipose-derived MSC at the time of disease onset significantly reduced disease severity, intestinal inflammation, and prevented repeat episodes of colitis [23]. Similar findings were obtained when 106 BMMSC were systemically infused 2 days after disease onset in vivo [24]. Priming BMMSC with IFNγ before infusion into mice with colitis reportedly enhanced these effects [25]. These in vivo studies suggest that MSC treatment may be beneficial to patients at the early stages of chronic irritable bowel syndromes. However, whether MSC will have beneficial effects in models of established refractory Crohn's disease require further investigation.
The clinical efficacy of MSC therapy for the patient's refractory to current immunosuppressive treatments has yet to be proven. Only 3 of the 13 trials looking at the clinical benefit of MSC treatment in Crohn's disease have been completed [5]. The initial phase I safety study reported no adverse effects from local injection of autologous early passage adipose MSC into eight fistulas in four patients at 12–30 months posttreatment [26]. Excitingly, MSC therapy promoted epithelial regrowth and healing of 75% of injected fistulas [26]. Similar observations were reported in a multicenter randomized phase II trial incorporating a small patient cohort with complex perianal fistulas associated with Crohn's disease [27]. Local administration of ex vivo-expanded adipose-derived MSC (2–6×107) with a fibrin glue caused fistula healing in ∼70% of treated patients when compared to those treated with fibrin glue alone as a control [27]. The reparative properties of MSC that led to complete fistula closure would appear to exceed what is achievable with current surgical and pharmacological interventions [27]. However, the effects of MSC on the inflammatory aspects of Crohn's disease were not assessed in these trials. In contrast, the systemic infusion of autologous culture-expanded BMMSC (1–2×106 cells/kg) reduced the severity of Crohn's disease in only 3/10 patients, while 7/10 patients did not respond to treatment (6 weeks posttreatment) [28]. Despite this, autologous MSC from all patients suppressed T cell proliferation in vitro, indicating that differences in patient outcomes were not caused by defects in the MSC immunomodulatory capabilities, but are potentially due to differences in route of administration [28].
It would appear that site-specific injection of MSC is a more effective treatment than systemic injection in these studies, however, the clinical outcomes measured were different (fistula healing vs. disease activity) [26 –28]. Moreover, data suggest that MSC treatment may only benefit certain patient groups with Crohn's disease, although further preclinical work is needed. Despite these discrepancies, a number of phase I/II clinical trial are currently being carried out to assess the safety and efficacy of MSC treatment on the clinical outcome of patients with Crohn's disease [5].
Systemic lupus erythematosus
SLE is characterized by hyperactive B-cells that produce autoreactive antibodies against RNA-binding proteins, phospholipids, and double-stranded DNA, which can lead to multiorgan dysfunction [29]. Systemic administration (intravenous injections) of 106 human umbilical cord-derived or BMMSC at the time of disease onset reduced disease severity in the lpr-mutant model of SLE, assessed by measuring proteinuria, circulating autoreactive antibody levels, or the degree of interstitial pneumonitis [29,30]. In vitro, human BMMSC suppressed proliferation of lpr-mutant T cell populations in a dose-dependent manner, while 106 umbilical cord-derived MSC induced the expansion of lpr-mutant TReg cells [30,31]. Interestingly in the NZBxNZW F1 model of SLE, priming of 1.25×106 BMMSC with IFNγ enhanced the MSC-induced suppression of B-cell proliferation and differentiation in vitro and reduced lymphocyte infiltration in vivo [32]. In contrast, allogeneic BMMSC treatment exacerbated disease in the NZBxNZW F1 model when administered either before or after disease onset [33]. Therefore, the efficacy of MSC therapy in vivo appears to be dependent on the model and the MSC population used. Whether MSC can ameliorate established disease has not yet been explored.
Despite this, three phase I/II clinical trials are currently examining the safety and therapeutic benefit of MSC therapy in SLE patient's refractory to current treatment protocols [5]. None of these studies include an appropriate control cohort for direct comparison. An initial pilot study in four patients with treatment refractory SLE reported that systemic administration of allogeneic ex vivo-expanded BMMSC (more than 106/kg) improved renal function and reduced SLE disease activity, such that patients were in disease remission for up to 12 months after treatment [34]. Excitingly, two patients suspended further treatment after 3 months follow-up [34]. A subsequent phase I/II study also demonstrated improvement in disease index score, proteinuria, and serological markers following autologous BMMSC infusion (106 cells/kg) in 11 of the 13 patients assessed at 12 months, with the remaining two patients going in to relapse [35]. Similarly, all 16 patients with refractory SLE showed a significant improvement in disease index scoring following systemic allogeneic umbilical cord-derived MSC treatment (106 cells/kg) in a phase II multicenter trial [36]. It has been postulated that the marked improvement in SLE disease activity is, in part, due to expansion of TReg cells, although the exact mechanism remains unknown [36]. None of these trials reported any adverse effects at 12 months posttreatment, however, the follow up period was too short and potential long-term effects in these studies require further investigation [34 –36]. Additionally, the studies lacked the appropriate treatment controls (where no MSC are added), making it difficult to ascertain the true benefit of MSC infusion. Even with these unknowns, these early data strongly suggest that MSC treatment may have substantial benefit for patients suffering from SLE. As a result, several large-scale, prospective, double blind, multicenter trials are now underway to optimize the treatment protocol [5].
Rheumatoid arthritis
RA is characterized by the expansion of the synovial fibroblast population, aberrant leukocyte infiltration, and elevation in the levels of cytokines and proteases within the joint leading to bone erosion and cartilage destruction [37]. Studies have reported that treating DBA/1 mice that have established collagen-induced arthritis (CIA) with 1–2×106 BMMSC or umbilical cord-derived MSC (intraperitoneally or intravenously, respectively) reduced disease severity [37 –39]. These groups detected less IL-6 and TNFα and more IL-10 in lymph nodes and joints, as well as an increased number of TReg cells in the spleen and lymph node, and a reduced ratio of Th1:Th17 cells in vivo [37 –39]. Collectively, these data indicate that various types of MSC administered systemically or by intraperitoneal injection could dampen the pathogenic immune response of CIA. However, conflicting findings have been reported in the CIA and adjuvant-induced arthritis (AIA) models and also when different mouse strains have been used [40 –42]. A single systemic infusion of primary (1×106 or 4×106) murine MSC administered to DBA/1 strain mice with established CIA or at the point of disease onset showed no beneficial effects of MSC treatment [40]. Additionally, a single infusion of 1×106 genetically mismatched murine BMMSC from the BALB/c strain into DBA/1 mice has been reported to increase local inflammation and exacerbate the symptoms [41]. Further, BMMSC (5×106) were only able to inhibit AIA onset, rather than ameliorate established disease [42]. It could be argued that the findings from the latter in vivo studies indicate that MSC are sensitive to their environment, which has the potential to alter their therapeutic potential (as alluded to earlier). Indeed, pretreatment of murine C3 MSC with TNFα was sufficient to reverse their immunosuppressive effects in vitro [40]. Thus, MSC may lose their immune regulatory effects when exposed to high levels of proinflammatory cytokines in the inflamed joint. If this was true, it would have a significant impact on the utility of MSC in the treatment of RA. Disparities between these in vivo studies are likely due to differences in the dose of MSC administered and the number of infusions given (see next section).
Trials in RA have lagged behind other autoimmune diseases primarily because of conflicting evidence generated by preclinical models. Currently, three phase I/II trials are underway looking at the effects of intravenous injection of primary human umbilical cord or adipose-derived MSC in patients with treatment refractory RA [5]. Cohort size varies from 20 to 200 patients and the main outcome measures are safety and disease progression [5]. Unfortunately, only 1/3 of these trials include an appropriate placebo control. The outcomes of these trials are expected in 2015–16.
Issues Limiting the Use of MSC as a Therapeutic Intervention for Autoimmune Diseases
Clinical efficacy and safety of MSC treatment has been shown in a number of trials for different autoimmune diseases. However, conflicting outcomes arise as a result of a lack of standardization in protocol design and understanding of basic MSC biology. The effects of MSC isolation and expansion, as well as route of administration may impact the effectiveness of treatment. Furthermore, the long-term effects of therapy are not clearly understood. Herein, we highlight the potential risks and limitations of the current MSC trials.
Heterogeneity and efficacy of different MSC populations
MSC are a heterogeneous population of stromal cells for whom there are currently no specific markers. Our best definition of an MSC is provided in the International Society for Cell Therapy 2006 guidelines, which state that MSC are a plastic adherent population that undergo clonal expansion, express stromal markers CD73, CD90, and CD105, while lacking expression of CD19/20, CD34, and CD45, and are capable of differentiating down mesodermal lineages [4]. More recently, certain cell surface proteins (CD146 and CD271) have been described as markers associated with highly potent (suppressive) MSC populations [43]. However, these markers are not exclusively expressed on MSC and are also expressed on other stromal cells (eg, fibroblasts) indicating that this criterion is too generic for defining a specific population for therapy. In the future, MSC will need to be more stringently defined and purified before being approved as a therapeutic intervention. MSC from different sources have been shown to differentially regulate immune responses [16] and trials have used several different MSC populations to test clinical efficacy. Moreover, there is strong evidence indicating that the conditions under which MSC are expanded ex vivo can affect their phenotype and potentially their function. Examples of these include changes in morphology, cell size, differentiation capacity, and expression of surface markers, including CD44, CD105, CD146, and CD271 [44]. More recently, changes in the nano- and microenvironment (ie, the surface on which the cells are cultured) and the additives contained within the culture media have also been shown to affect the proliferation rate of MSC [45]. It seems likely that this heterogeneity and the plasticity of MSC underlie the different results generated during clinical trials. A standardized culture protocol needs to be devised that minimizes changes in MSC phenotype.
The optimum clinical grade cell type should be identified and used for all future trials. However, there are currently no functional markers that can be used to isolate MSC and distinguish them from fibroblasts or other MSC-like stromal cells. This has major therapeutic implications as the MSC currently being used for therapy may contain other stromal cell contaminants, the effects of which are currently unknown. The level of contamination may also vary between studies which could, in part, explain the variations in outcomes. For therapy, infusion of a homogeneous population that exerts the required effect would be ideal, however, due to the heterogeneity of MSC cultures this has proven difficult. Despite these issues, Osiris therapeutics has recently received FDA approval to use Prochymal BMMSC for the treatment of grafts versus host disease and Crohn's disease [46]. While these cells have been extensively characterized and rigorously tested in preclinical models, the heterogeneity of the cell population is still poorly understood and will need to be more clearly defined before becoming an off the shelf therapeutic. Orbsen therapeutics has recently isolated a more homogeneous or pure population of MSC from bone marrow, placenta, skeletal muscle, and fat based on expression of the CD362 (Syndecan-2) [47]. Early preclinical studies are testing the potency of these cells and they may provide a more attractive source for therapy [47].
Effect of ex vivo expansion on immunomodulation
Early phase I trials have shown that MSC have clinical efficacy for treating a range of disorders (eg, SLE and Crohn's disease); however, large phase II trials have not shown any significant benefits [23 –26,29 –36]. To test the efficacy of a specific MSC population, the patient cohort will need to be treated with MSC from the same source. Therefore, large-scale ex vivo culture expansion of MSC is often required to generate sufficient numbers of cells. It is well known that MSC lose their proliferative, differentiation, and immunomodulatory capacities with culture and passage [48]. The studies described here have used a varied numbers of MSC, with some administering only 106 cells/kg, while others used 5×106 cells/kg [28,34]. It is unclear whether adding more MSC enhances the clinical efficacy, as some studies that used fewer cells still showed beneficial effects. Adult BMMSC lose their immunosuppressive capabilities quite early (passage 5–7), compared to fetal MSC populations, which retain their abilities during prolonged culture (∼25 passages) [49,50]. The majority of in vitro and early in vivo studies have isolated MSC based on plastic adherence and characterized them at passage 1–3 for expression of the marker panel. Equally, preclinical studies have used MSC from passages 1–5. However, in larger trials MSC will need to be expanded beyond passage 5 to generate sufficient numbers to treat patient cohorts. One major concern is the loss of MSC immune regulatory functions due to their ex vivo culture or adverse response to the culture environment. The effect of ex vivo culture on the integrity of MSC and their functionality needs to be more clearly defined.
Route of administration
MSC have been shown to have a very low homing efficiency (<1%) when systemically infused and the majority of studies suggest that they mostly become trapped in the lungs and the liver [reviewed by 4,51]. Based on these data, the beneficial effects observed posttreatment are more likely due to the mass production of soluble mediators that alter the function of immune cells and are not due to specific effects on target tissues [52]. However, a recent study challenges the current dogma that MSC show good survival postinfusion. Indeed they demonstrated that systemically infused BMMSC became trapped in the lungs and could not be detected 24 h postinjection. Surprisingly, viable infused BMMSC were not detected in any other organ following isolation and reculture in vitro [53]. The study concluded that reports suggesting that MSC redistributed to other tissues were actually detecting fluorescently labeled cell debris or phagocytosed MSC, and postulated that any long-term immunosuppressive effects observed after MSC infusion are mediated by other cell types and not the MSC themselves [53]. Therefore, site-specific injection of MSC may have a greater efficacy than systemic infusion, as observed in RA [39]. Increasing the proximity of MSC to their leukocyte targets will allow them to mediate their suppressive effects through direct cell contact as well as the generation of soluble factors. Standardizing the delivery method of MSC therapy is required to maximize the treatment efficacy. In diseases where MSC recruitment would be low, the benefit of direct site-specific administration will need to be compared to systemic infusion.
Long-term side effects of treatment
The clinical studies discussed in this review have generally undertaken follow-up checks on patients up to 18 months after treatment. While early benefits and risks, such as disease relapse, responsiveness to pharmacological therapy, and disease index scorings, were assessed, the long-term effects (ie, 5–10 years posttreatment) have not been performed. It is currently unknown whether the beneficial effects of treatment are short lived, if repeat doses are required to maintain effects, and whether the condition worsens again after a few years. Additionally, the speed by which MSC are cleared from the patient is also unknown. In SLE, patients displayed improvements up to a year after MSC infusion even though the MSC are probably rapidly cleared from the blood [34,52,53]. MSC are thought to activate endogenous regulatory responses which carry out the majority of the anti-inflammatory effects observed. However, these have not been clearly defined and will need to be elucidated. As MSC are only just emerging as a potential therapy, their response to other pharmacological interventions, potential tumorigenicity, and tissue distribution upon administration are key areas in need of further investigation.
Potential contribution of MSC to disease pathogenesis
The effect of the inflammatory microenvironment on the endogenous MSC phenotype has not been well characterized. MSC may undergo transformation in response to cytokines in the inflammatory milieu and as a consequence lose their immunomodulatory capabilities or indeed become proinflammatory. Polarization of MSC to a proinflammatory phenotype has been shown to occur through stimulation of TLR4 [18]. It is unclear whether other inflammatory mediators may also induce this switch in MSC phenotype. The inflammatory microenvironment has previously been shown to modify the phenotype of other stromal cell types. In RA, fibroblasts promote joint inflammation in response to the proinflammatory microenvironment [17]. As MSC are sensitive to their environment, it is likely that their function will be altered during chronic inflammation. Indeed, evidence suggests that MSC isolated from patients with RA lose proliferative potential, but maintain the ability to differentiate and immunomodulate [54]. Moreover, synovial fluid from RA patients was able to alter the transcriptional profile of healthy MSC, indicating that MSC are sensitive to factors in the inflammatory milieu [55]. A recent study has also shown that endogenous MSC-like perivascular progenitor cells may contribute to fibrosis after organ damage [56]. Collectively, these studies suggest that MSC are sensitive to their tissue environment and could potentially switch from an anti-inflammatory to a proinflammatory state depending on the environmental cues that they receive. The effect of the inflammatory microenvironment on MSC function will require further examination.
Alternative treatment strategies
As the majority of effects induced by MSC are thought to be mediated by soluble factors, identifying and administering specific mediators released by the MSC may be a better therapeutic strategy. Indeed, preclinical studies have shown that MSC-conditioned medium possesses similar bioactivity to MSC themselves [57]. Therefore, MSC-derived factors or conditioned medium should also be tested for efficacy in trials of autoimmune disease. Such strategies could potentially minimize the risks of MSC therapy, although multiple dosages may be required as the cells will continue to release immunomodulatory mediators over their lifespan. To date, no clinical studies testing the effect of MSC-derived soluble mediators in autoimmune disease have been conducted.
Conclusion
In conclusion, preclinical and clinical trial data have shown efficacy in MSC treatment inflammatory disorders, although the results are often inconclusive and clinical benefit varies between trials [23,27,29,34,38]. This is likely to be due to differences in trial design, MSC dosage, the evaluation of clinical benefit, cell isolation protocols, and the MSC populations used. The generation of clinical-grade MSC shows that we are now bridging the translational gap between scientific observations on MSC function and clinical applications for therapy. To test the true efficacy of MSC, the treatment protocol will need to be standardized to minimize the off-target effects and safety risks. In vitro data strongly suggest that MSC act as endogenous regulators of tissue inflammation, but whether this translates to the clinical stage remains to be seen. Key to this will be what effect the inflammatory microenvironment has on the phenotype of administered MSC and our ability to minimize any potential side effects. Until these issues are addressed, the safety and efficacy of MSC treatment cannot be fully ascertained.
Footnotes
Acknowledgments
H.M. was supported by a BBSRC University of Birmingham PhD studentship. H.M.M. was supported by an Arthritis Research UK Career Development Fellowship (19899) and Systems Science for Health, University of Birmingham (5212).
Author Disclosure Statement
No competing financial interests exist.