
Abstract
There is considerable evidence that stem/progenitor cells reside in the vasculature during the prenatal and postnatal stages. The stromal vascular fraction (SVF) of human adipose tissue is markedly rich in blood vessels, and it is a source of mesenchymal/stromal cells (MSCs). Therefore, we hypothesized that, in addition to MSCs, the SVF may contain other mesodermal precursors. However, the SVF has a high content of CD34+ cells with high proliferative capacity, which can prevent the growth of the most quiescent cells. By using an antifibroblast (FIB) antibody coupled to microbeads, we show that ∼90%–95% of the nonhematopoietic CD34+ cells were retained in the CD45−FIB+ fraction. Reverse transcription-polymerase chain reaction analysis revealed that the CD45−FIB−CD34− cell fraction expressed higher mRNA levels of KDR and GATA2 than its complementary CD45−FIB−CD34+ cell fraction, which contained the SVF endothelial cells. Surprisingly, when CD45−FIB−CD34− cells were cultured in endothelial growth medium, they gave rise to endothelial colonies and mesenchymal colonies. Moreover, when CD45−FIB−CD34− cells were cultured in embryonic stem cell expansion medium, they gave rise to cells exhibiting the full range of phenotypes observed in the freshly isolated SVF, including CD34+ and CD31+ cells. Together, these results suggest that the CD45−FIB−CD34− cells within the SVF of human adipose tissue function as mesodermal precursors of mesenchymal and endothelial cells.
Introduction
I
These findings raise an interesting and exciting possibility that the SVF contains mesodermal precursors that are more primitive than previously identified. In support of this hypothesis, we previously demonstrated the presence of small KDR+CD34+/− cells with hemangioblastic properties [18]. Here, we report that most of the nonendothelial CD34+ cells, contained in the SVF express the fibroblast (FIB) antigen, are plastic-adherent and are the source of origin for the fibroblastoid stromal cells. Alternatively, by depleting the SVF fraction of CD45+, FIB+, and CD34+ cells, we could show that the CD45−FIB−CD34− cells gave rise in culture to endothelial colonies and mesenchymal colonies.
Materials and Methods
Subjects
Subcutaneous abdominal adipose tissue was obtained from 12 female patients undergoing abdominoplasty after informed consent was obtained. The age of the women ranged from 25 to 50 years (42.8±9.9 years), the body mass index (BMI) from 23.2 to 44.3 kg/m2 (31.3±6.0 kg/m2), and the systolic and diastolic blood pressure from 104 to 155 mm Hg (127±14 mm Hg) and from 50 to 88 mm Hg (70±11 mm Hg), respectively; the subjects were free of metabolic or endocrine diseases. The study was approved by the Clinical Research Ethics Committee of the Valencia University General Hospital.
Cell isolation and culture
The SVF from adipose tissue was obtained after digestion of the tissue with collagenase as previously described [18]. The SVF cells were magnetically labeled with CD45 microbeads and then separated using the MACS separation system (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany). The CD45-negative fraction was then incubated with antifibroblast (FIB)- or anti-CD34-microbeads, and the CD45−FIB−, CD45−FIB+, CD45−CD34−, and CD45−CD34+ cell populations were collected. Finally, the CD45−FIB− fraction, which was free of magnetic beads, was labeled with CD34-microbeads and then separated into CD45−FIB−CD34+ and CD45−FIB−CD34− cells (Supplementary Fig. S1; Supplementary Data are available online at
Proliferation of CD45− cells
Freshly isolated CD45−FIB− and CD45−FIB+ cells were plated at a density of 30,000 cells per cm2 in endothelial growth medium (EGM) composed of endothelial basal medium (EBM-2) supplemented with EGM-2 MV SingleQuots containing vascular endothelial growth factor, basic fibroblast growth factor, insulin-like growth factor-1, epidermal growth factor, and 5% fetal bovine serum (Lonza Walkersville, Inc., Walkersville, MD). After 24 h of culture, the nonadherent CD45−FIB− cells were removed, and the adherent cells continued to grow. Upon reaching ∼75% confluence, the primary cell monolayers were detached by treatment with 0.25% trypsin-ethylenediaminetetraacetic acid (EDTA) solution (Lonza Walkersville, Inc.), further subcultured as P1 at a density of 5,000 cells per cm2, and expanded up to passage 5. The cultures were maintained at 37°C in a humidified atmosphere of 5% CO2. The population doubling level (PDL) was calculated using the formula PDL=log (total viable cells at the end of the incubation time/total viable cells at the beginning of the incubation time)/log2. The population doubling time (PDT) was calculated as T/PDL, where T is the incubation time in days.
Generation and culture of EPCs
Freshly isolated CD45−FIB−CD34+ cells were plated at (5, 10, 20, 50, or 100)×103 cells/cm2 in EGM in six-well plates. After 1 day of culture, the adherent cells were discarded, and the nonadherent cells were reseeded into replicate plates. After 7 days of culture, cells that did not adhere to plastic tissue culture plates were reseeded in replicate plates coated with fibronectin (10 μg/mL) and type I collagen (30 μg/mL) from rat tail (BD Biosciences, Erembodegem, Belgium). Adherent and nonadherent cells were grown in EGM. The CD45−FIB−CD34− cells were seeded at the same densities and in the same conditions, except that the cells only underwent one cycle of 24 h of adherence to the plastic tissue culture plates. Half the volume of the culture medium was replaced every 3 days. The cell cultures were examined daily for endothelial colony formation, and the frequency of colony formation was estimated. The cells were detached by trypsinization and plated in 6-well plates or tissue culture flasks in EGM at 5×103 cells/cm2. Subconfluent cells were expanded for five passages, and the PDL and PDT were calculated. All cultures were maintained at 37°C and 5% CO2 in a humidified atmosphere.
Culture of CD45−FIB−CD34− cells in defined embryonic stem cell expansion medium
CD45−FIB−CD34− cells were seeded in six-well plates coated with Matrigel hESC-qualified matrix (BD Biosciences) in mTeSR1, a serum-free medium containing bovine serum albumin, lithium chloride, pipecolic acid, γ-aminobutiric acid, and high levels of human recombinant transforming growth factor β and human recombinant basic fibroblast growth factor, or in mTeSR2, a xeno-free version of mTeSR1 (StemCell Technologies, Grenoble, France) at 5×103 or 10×103 cells/cm2. The cell cultures were incubated at 37°C in a humidified atmosphere containing 5% CO2 and either 20% O2 (normoxia) or 5% O2 (hypoxia).
Culture of CD45− cell populations in semisolid medium
CD45−FIB− or CD45−FIB−CD34− cells that did not adhere to the plastic surface were seeded at 1.5×104 or 2.5×104 cells/cm2 in Petri dishes coated with fibronectin and collagen in a methylcellulose-based media with or without serum (Methocult GF H4435 or Methocult SF H4436; StemCell Technologies). The cultures were incubated at 37°C in a humidified atmosphere containing 5% CO2. Images were acquired using a Leica EC3 camera and a Leica DMIRE2 inverted microscope.
Flow cytometry analysis
Freshly isolated SVF cells, cultured SVF cells, and endothelial cells were analyzed for surface marker expression using the FACSCanto II and FACSDiva software (BD Biosciences). The cells were resuspended in phosphate-buffered saline containing 1 mM EDTA and 1% bovine serum albumin, and they were incubated with human Fc block for 5 min to inhibit nonspecific binding. The cells were then incubated with primary antibody (Table 1). Monoclonal antibodies were conjugated to fluorescein-isothiocyanate (FITC), phycoerythrin (PE), allophycocyanin (APC), PE-cyanin7 (PE-Cy7), or APC-cyanin7 (APC-Cy7). The cells were incubated with specific antibodies or isotype-matched controls for 30 min at 4°C in the dark. For removal of nonviable cells in the analysis 7-Amino-actinomycin D (7-AAD; Sigma-Aldrich, Madrid, Spain) was used.
Immunocytochemistry and acetylated LDL cell uptake
Early-passage (P2 or P3) endothelial cells were seeded on glass coverslips. The cells were fixed, blocked, and incubated with rabbit anti-human von Willebrand factor (vWF, 1/100 dilution; Sigma-Aldrich) or a rabbit polyclonal antibody against endothelial nitric oxide synthase (eNOS 1/100 dilution; Abcam, Cambridge, United Kingdom) and subsequently incubated with goat anti-rabbit Alexa Fluor 555 or goat anti-rabbit Alexa Fluor 488 (Jackson ImmunoResearch Laboratories, West Grove, PA). The cells were washed and counterstained with 4′,6-diamidino-2-phenylindole dihydrochloride (DAPI; Vector Laboratories, Burlingame, CA). The incorporation of DiI-acetylated low-density lipoprotein (DiI-Ac-LDL; Molecular Probes, Life Technologies, Foster City, CA) was evaluated by fluorescence microscopy after incubation with 10 μg/mL DiI-Ac-LDL for 4 h at 37°C.
CD45−Fib−CD34− cells cultured in stem cell medium were stained at P1 with antibodies against SSEA-3, Sox2, Tra-1-60, Tra-1-81, Oct4 (hES/iPS Cell Characterization Kit; Applied Stem Cell, CA), or anti-CD31 (clone JC70A; Dako, Glostrup, Denmark) and were counterstained with DAPI.
In vitro and in vivo Matrigel assays
The capillary tube formation assay was performed by plating endothelial cells (P2 or P3) in wells coated with Matrigel basement matrix (BD Biosciences) at a density of 10×103 or 20×103 cells per well. When capillary formation was complete (24 h), the cells were fixed and stained with mouse anti-human CD31 (Dako) followed by Alexa Fluor 488 goat anti-mouse (Jackson ImmunoResearch Laboratories). For the in vivo Matrigel assay, a total of 1×106 endothelial cells were mixed with Matrigel growth factor reduced (BD Biosciences) and subcutaneously implanted into the dorsal flank of 8- to 12-week-old female Hsd:athymic nude-Foxn1nu mice (Harlan Laboratories, Udine, Italy). After 14 days, the Matrigel plugs were removed and processed for hematoxylin and eosin (H&E) staining or for immunofluorescence as previously described [19]. The sections were immunostained with rat anti-mouse CD31 (1/25 dilution, clone MEC13.3; BD Biosciences) or biotinylated Ulex europaeus agglutinin 1 (UEA-1, 1/100 dilution; Vector Laboratories) and were then incubated with Alexa Fluor 555 goat anti-rat or fluorescein (DTAF)-conjugated streptavidin (Jackson ImmunoResearch Laboratories). Images were acquired using a Leica DFC480 camera and Leica DM6000 B microscope with the Leica Application Suite (LAS), Version 3.6.0.
Gene expression analysis
Isolated cell populations from the SVF, endothelial cells, and CD45−FIB−CD34− cells that were nonadherent to fibronectin- and collagen-coated plates were collected and kept frozen at −80°C until RNA extraction. Total cellular RNA was prepared using TRIzol Reagent, and the RNA was treated with DNase to avoid genomic DNA contamination. RNAs were reverse-transcribed using TaqMan Reverse Transcription Reagents. The assays were selected from the TaqMan Gene Expression Assay library: SCL/TAL1: Hs001097987_m1, GATA2: Hs00231119_m1, KDR: Hs00911700_m1, CD34: Hs00156373_m1, PECAM1 (CD31): Hs00169777_m1, ENG: Hs00923996_m1, TIE2: Hs00945146_m1, NANOG: Hs 02387400-g1, POU5F1 (OCT4): Hs00742896_s1, SOX2: Hs01053049_s1. Gene expression was normalized to the endogenous housekeeping gene glyceraldehyde phosphate dehydrogenase (GAPDH) or 18S rRNA. All polymerase chain reaction reactions were performed in duplicate. The results were analyzed using the ABI PRISM 7900HT Sequence Detection System. The 2−ΔCt method was used to calculate the relative gene expression levels, and the 2−ΔΔCt method was used to calculate the relative changes in gene expression levels. All reagents were purchased from Applied Biosystems (Life Technologies).
Statistical analysis
The results are expressed as the mean±standard deviation. The data were analyzed using GraphPad Prism Software 5.0 (GraphPad Software, Inc., La Jolla, CA). The comparisons among groups were analyzed by Student's t-test. Differences were considered to be statistically significant at P<0.05.
Results
Flow cytometric characterization of CD34+ cells from the SVF
Compared to bone marrow, the SVF of human adipose tissue is very rich in CD34+ cells, although the proportion of these cells has been reported to be highly variable [12,18 –22], and also donor-specific [23]. Nonetheless, given that the enumeration of CD34+ cells is performed by flow cytometry, it is possible that the high variability in the content of CD34+ cells is in part due to the use of different threshold settings for sample acquisition. After setting the threshold value of forward scatter (FSC) for the analysis of CD34+ hematopoietic progenitor cells (HPCs) in hematopoietic tissues, leukocytes, which were identified as CD45+ cells, accounted for 23%±15% of the SVF cells (values ranging between 2.3% and 56.6%; n=12). As shown in Fig. 1A, the leukocytes exhibited lower FSC and side scatter (SSC) properties than CD45− cells. Under these conditions, the proportion of CD34+ cells in the CD45− cell fraction was 83%±20% (ranging from 20.5% to 97.1%; n=12) (Fig. 1A). However, after setting the threshold value for FSC to acquire the small viable cells (Supplementary Fig. S2), the proportion of CD34+ cells within the CD45− cell subset was 36%±20% (range 6% to 63%; n=12) (Fig. 1B). These FACS analyses allowed us to demonstrate that the SVF CD45− cell fraction is a complex population containing cells with different FSC and SSC properties (Fig. 1). As previously described [24] BMI was positively correlated to percentage of SVF CD45+ cells (r 2=0.568), and thus, taking into consideration the number of stromal cells obtained after tissue digestion, the number of SVF CD45− cells was 0.873×106±0.594×106 cells/g of tissue (ranging from 0.224×106 to 1.732×106 cells/g; n=12) and that of SVF CD45−CD34+ cells was 0.326×106±0.291×106 cells/g of tissue (ranging from 0.039×106 to 0.858×106 cells/g; n=12).
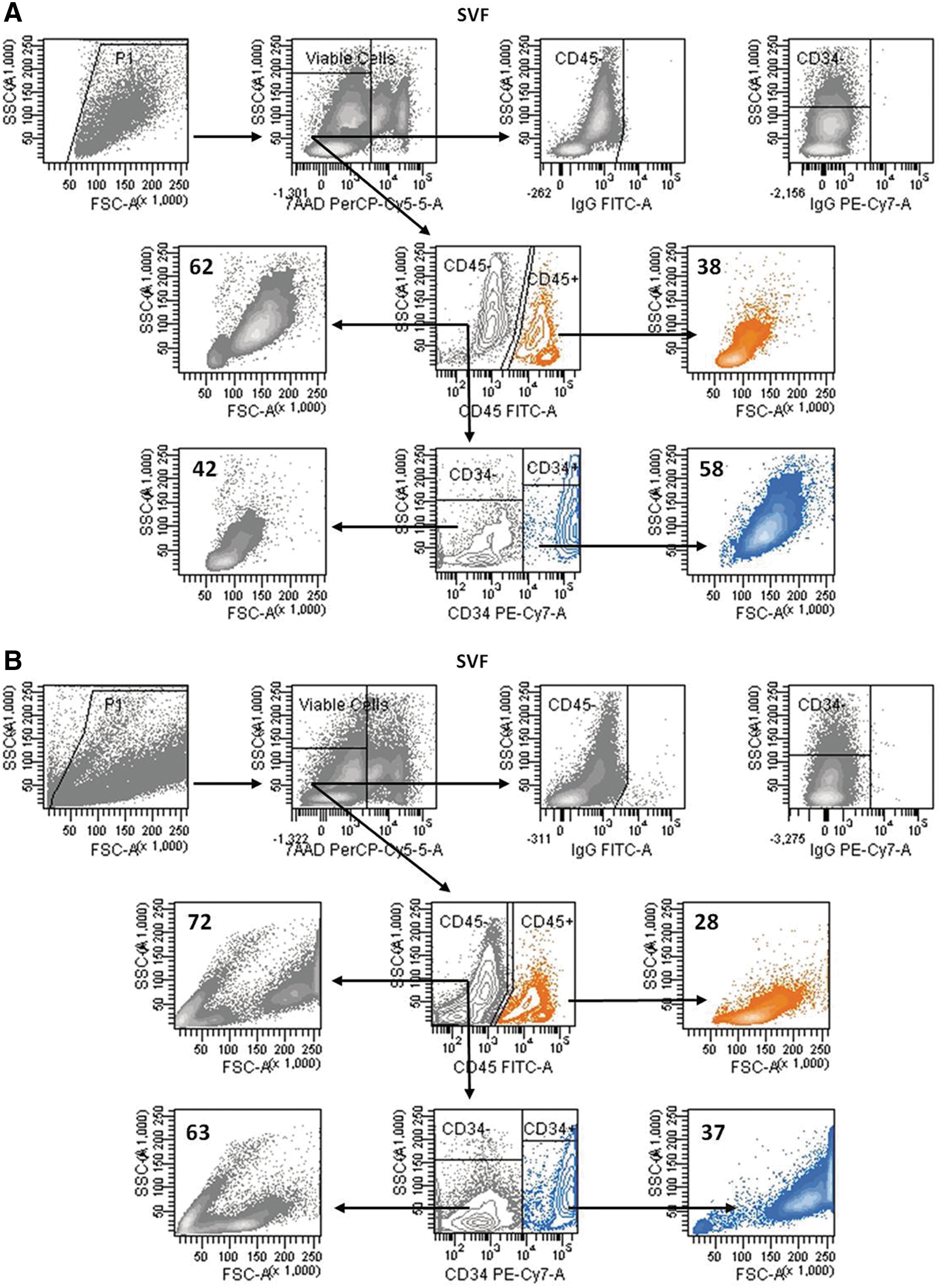
FSC and SSC FACS plots of the SVF cells from human adipose tissue. Freshly isolated SVF cells were stained with CD45 (FITC) and CD34 (PE-Cy7) antibodies and then acquired with a high FSC threshold value
The antigenic profiles of the two CD45−CD34+ and CD45−CD34− cell populations differed greatly. Nearly all of the CD34+ cells expressed the well-recognized markers of adult stem cells, including CD90, CD9, and CD13, whereas only 10%–15% of the CD34− cells were positive for these antigens. The expression of the MSC antigens CD143/BB9, MSCA-1, and CD105 and the endothelial markers CD31 and CD144 was restricted to CD34+ cells. Moreover, CD146, a perivascular antigen, was detected mainly in the CD34+ cells. CD117, Tie-2, KDR, and Flt-1, which identify HPCs and EPCs, were expressed by a very small proportion of cells in either the CD34+ or CD34− cell subsets, whereas CD133 expression was not detected in any of the cell subsets (Table 2).
Phenotype of freshly isolated CD34+ and CD34− cells within the SVF CD45− cell subset as detected by flow cytometry. Values indicate the percentage of cells expressing the selected antigens. Results are expressed as the mean±SD from 12 independent experiments with SVF cells derived from 12 different donors. Significant difference, CD45−CD34+ cells versus CD45−CD34+ cells, b P<0.02, a P<0.000001.
nd, not detected; SVF, stromal vascular fraction.
Most of the CD45−CD34+ cells comprising the SVF express the fibroblast antigen
To further characterize the SVF CD45− cells, we took advantage of the fibroblast antigen, which is expressed in MSCs from human bone marrow [25,26]. Freshly isolated CD45− cells were separated into fibroblast-positive (FIB+) and fibroblast-negative (FIB−) cells, (purity of CD45− and CD45−FIB− cells was always greater than 95%, Supplementary Fig. S3). In terms of total cell numbers, the CD45− cell fraction represented 81%±7% of the CD45+ and CD45− cells collected (the recovery was 75%±15%; n=12). Notably, the CD45−FIB+ cell fraction accounted for 84%±9% of the CD45−FIB+ and CD45−FIB− cells collected (75%±25% of recovery; n=8). Alternatively, SVF cells were first separated into FIB+ and FIB− cells, and then the FIB− cells were separated into FIB−CD45− and FIB−CD45+ cells. The FIB+ cell fraction accounted for 67%±10% of the total FIB+ plus FIB− cells recovered (n=8), and as expected, CD45+ cells were barely detected in this fraction (data not shown). The FIB−CD45− cell fraction represented 25%±20% of the FIB−CD45− and FIB−CD45+ cells collected. Therefore, regardless of the strategy used for cell isolation, the data show that ∼70% of the SVF cells correspond to CD45− cells expressing the fibroblast antigen. As shown in Fig. 2A most of the cells (70%±18%; n=5) contained in the CD45−FIB+ cell subset had high FSC values; by contrast, the majority of cells (76%±17%; n=5) comprising the complementary CD45−FIB− cell subset corresponded to small cells (low FSC). FACS analyses showed that the CD45−FIB+ cell subset was more enriched for CD34+ cells than its complementary CD45−FIB− cell fraction (59%±22% versus 23%±20%; P<0.02; n=5), although the CD34+ cells in both cell subsets showed a similar pattern of CD105, CD90, CD9, and CD13 expression (data not shown). Moreover, the CD143/BB9 and MSCA1 antigens, which identify mesodermal progenitors and MSCs, respectively [27 –29], were mostly expressed in the CD45−FIB+ cell subset, whereas almost all of the CD34+CD31+ endothelial cells were contained in the CD45−FIB− cell fraction (Fig. 2A). As expected, the CD45−FIB+ cells were adherent, and the CD45−FIB− cells were shown to be nonadherent or loosely adherent to tissue culture plastic. Consequently, ∼70% of the cells remained rounded and floating after 24 h of culture. Next, the proliferation kinetics of the CD45−FIB+ and the adherent CD45−FIB− cells was determined. As shown in Fig. 2B, the CD45−FIB+ cells grew faster (average PDT of 5 days) than the CD45−FIB− cells in the primary cultures (average PDT of 10 days, P<0.001). However, during the subsequent passages, no significant differences in the PDT values were observed. Moreover, both of the CD45− cell subsets (FIB+ and FIB−) completed ∼15 PDLs after the fifth passage (Fig. 2B). As expected, adherent cells derived from the FIB+ and FIB− subpopulations gave rise to typical fibroblastoid cells that uniformly expressed CD90, CD105, CD13, and CD9 (data not shown). Furthermore, as previously described [30] the CD34 antigen expression disappeared when the SVF cells were passaged in culture. We observed that the expression of CD34 within the CD45−FIB+ cultured cells decreased dramatically (from ∼75% of cells expressing CD34 at the beginning of the culture to 3.6% and 0.3% after establishment of the primary culture and at the first passage, respectively), whereas in adherent CD45−FIB− cultured cells, the expression of CD34 decreased more gradually (Fig. 2C). Indeed, the CD34+ cells reached three PDLs by the first passage. CD31 exhibited a pattern of expression similar to that of CD34 (Fig. 2C), although no net expansion of the CD34+CD31+ cells was observed. This finding suggests that the adherent endothelial cells have little proliferative capacity or that the higher rate of proliferation of the other accompanying cells in the culture prevents their growth.
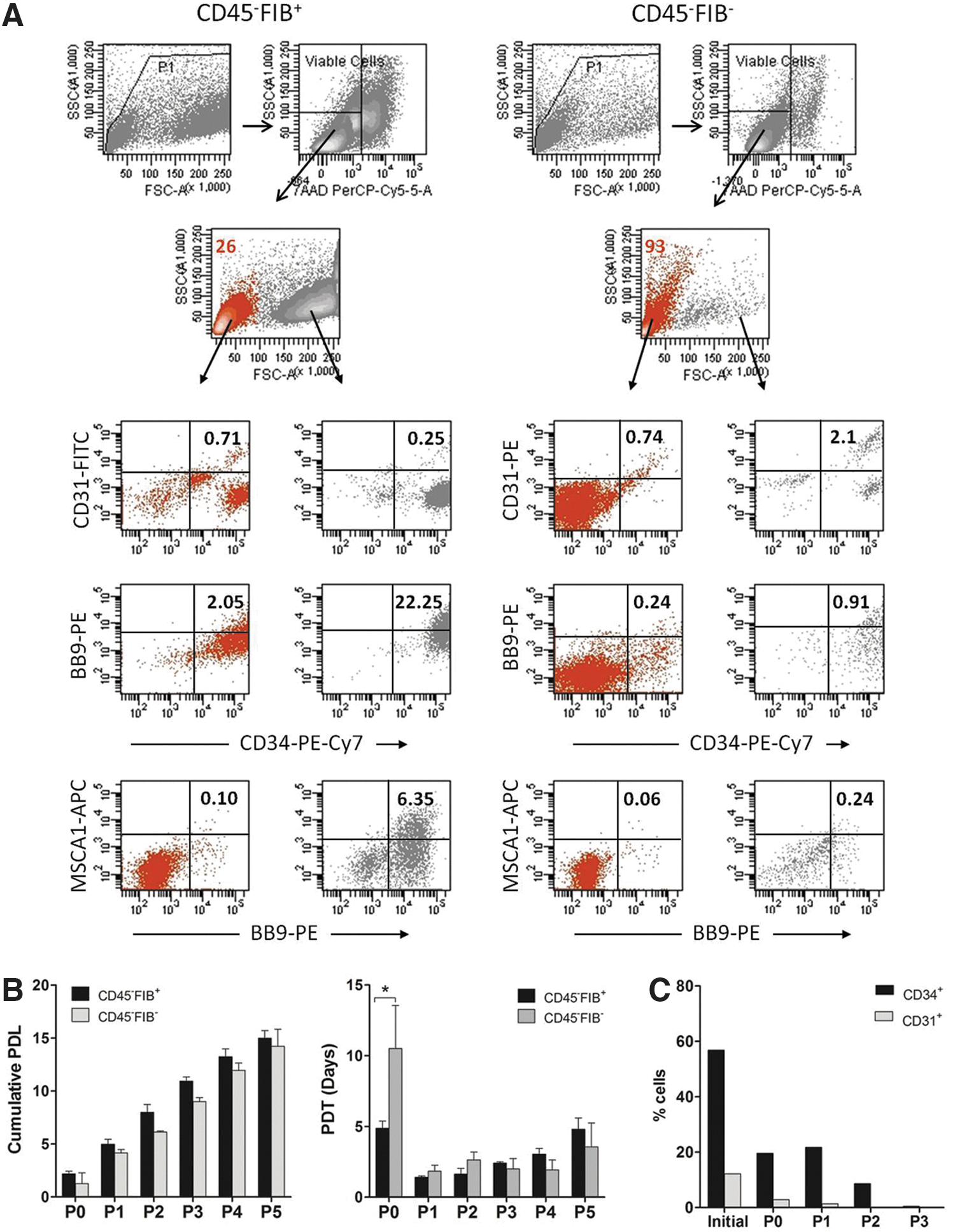
FACS analysis and growth kinetics of isolated CD45−FIB+ and CD45−FIB− cells. Freshly isolated CD45−FIB+ and CD45−FIB− cells were stained with CD31-FITC or CD31-PE, CD143/BB9-PE, CD34-PE-Cy7, and MSCA1-APC antibodies and analyzed by flow cytometry
The nonadherent CD45−FIB− cell population contains endothelial cells with proliferative capacity
Most of the CD45−FIB− cells that did not adhere to the plastic after 7 days in culture were small, and ∼90% of these cells lacked CD34 expression; however, ∼50% of the CD34-expressing cells expressed CD31 (Fig. 3A). To assess the ability of these endothelial cells to proliferate, the nonadherent CD45−FIB− cells were cultured in semisolid medium on fibronectin- and collagen-coated dishes. After 13 days in culture, the total number of cells increased by approximately 5-fold (4.6±2.3-fold; n=3), but the number of CD34+CD31+ endothelial cells increased 9-fold (9.3±3.2-fold; n=3). As previously reported [18,31], these culture conditions allowed the identification of HPCs residing in the SVF CD45− cell population, as evidenced by the generation of hematopoietic colony-forming units (Fig. 3B). Additionally, it is interesting to highlight that cells with endothelial-like morphology appeared as clonal-derived cells and, in many cases, were surrounded or circumscribed by stromal cells (Fig. 3C).
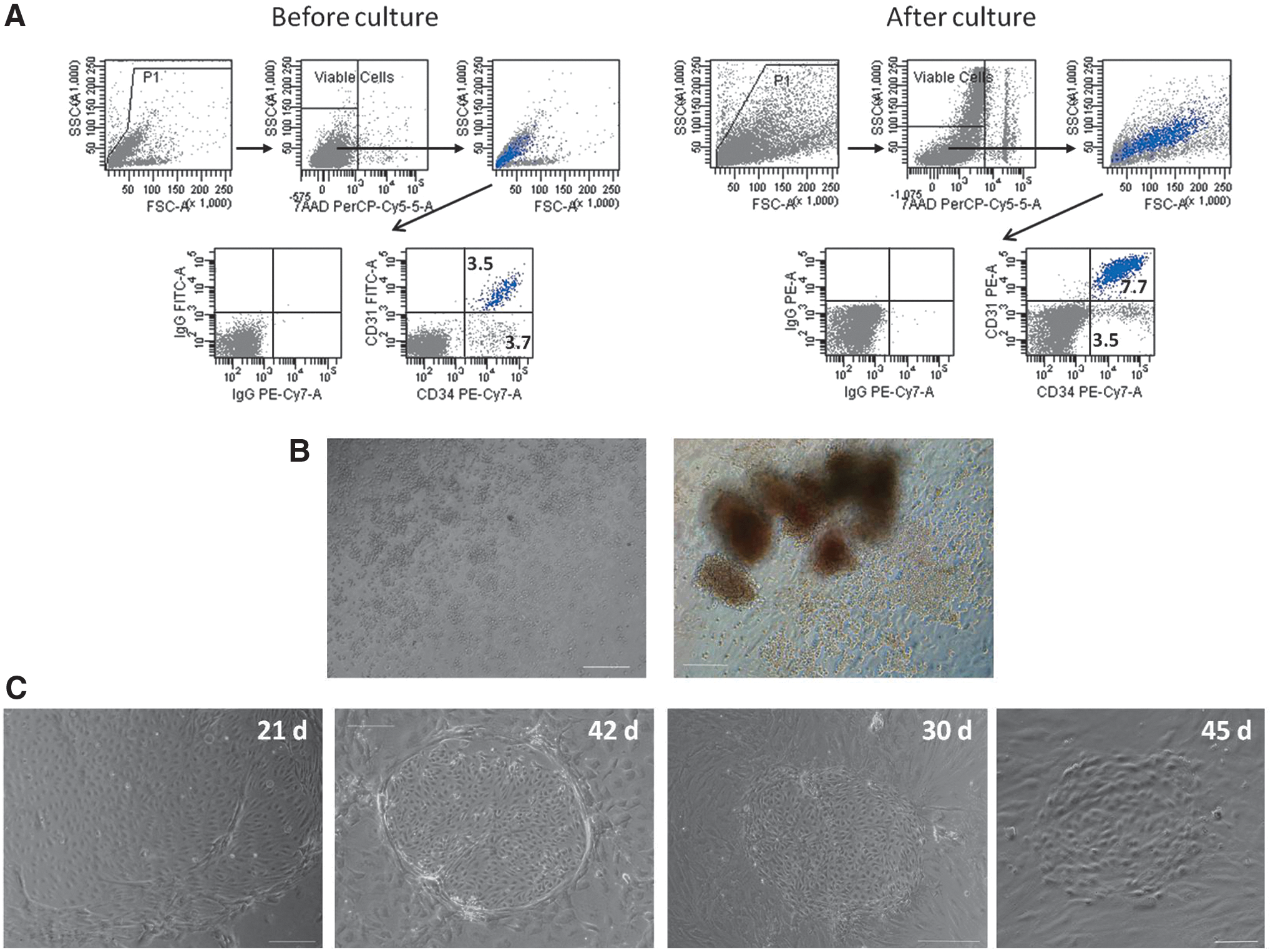
Proliferating endothelial cells reside in the small nonadherent CD45−FIB− cell population. After 7 days in culture, the nonadherent CD45−FIB− cells were collected and seeded in semisolid medium on fibronectin- and collagen-coated dishes.
CD45−FIB−CD34− cells from the SVF contain endothelial precursors
We previously described the existence of a rare population of small nonadherent CD45−KDR+CD34+/− cells in the SVF of human adipose tissue that displayed hemangioblastic properties in vitro [18]. Therefore, the possibility that endothelial precursors exist within the SVF must be considered. If this hypothesis is correct, one would expect that genes coding for transcription factors directly involved in hematopoietic and endothelial cell differentiation could be detected. The results show that KDR, SCL/TAL1, and GATA2 were expressed in the FIB−CD45+ and FIB−CD45− cell populations, although SCL/TAL1 expression was significantly higher in the FIB−CD45+ cells than in the FIB−CD45− cells (Fig. 4A), which was likely a result of the greater hematopoietic activity of the SVF CD45+ cells, as previously reported [31]. Recently, we reported that CD14+ monocytes residing in the SVF of human adipose tissue express the CD34 and KDR antigens [19]; accordingly, when FIB− cells were separated on the basis of CD34 expression, the level of KDR mRNA was significantly higher in the FIB−CD34+ cells than in the FIB−CD34− cells. Nevertheless, although CD34 is a well-recognized marker of HPCs and EPCs, the SCL/TAL1 and GATA2 mRNA levels were similar in the FIB−CD34+ and FIB−CD34− cell subsets, probably because both populations contain CD45+ hematopoietic cells. Finally, when FIB− cells were depleted of CD45+ cells and the resulting FIB−CD45− fraction was separated into FIB−CD45−CD34+ and FIB−CD45−CD34− cells, we observed that the CD34− cell fraction contained significantly higher levels of KDR and GATA2 mRNA than its complementary CD34+ fraction (Fig. 4A). Therefore, these results led us to hypothesize the existence of CD34− mesodermal precursors endowed with endothelial potential.

The SVF CD45−FIB−CD34− cells give rise to ECFCs.
FACS analysis showed that the purity of the isolated CD45−FIB−CD34− cells was always greater than 95% (Supplementary Fig. S3), and further that these cells lacked CD31 expression (Fig. 4B); additionally, CD45−FIB−CD34− and CD45−FIB−CD34+ cells expressed Tie2, KDR, and Flt1 at similar levels (∼1.0%, 0.5% and 5%, respectively), but CD9 and CD90 had higher expression levels in the CD34+ cell population (∼85% and 15%, respectively) than in its counterpart CD34− cell subset (∼30% and 2%, respectively). To assess the ability of these primitive CD34− cells to generate endothelial cells in vitro, freshly isolated CD45−FIB−CD34− cells were plated on uncoated 6-well plastic plates, and after 7 days, the nonadherent cells (Fig. 4C) were replated in replicate fibronectin/collagen-coated plates in EGM; in some experiments, the nonadherent cells were grown in semisolid medium. The CD45−FIB−CD34+ cells were used as a control, but to prevent the growth of the highly proliferative adherent CD34+ cells, CD45−FIB−CD34+ cells underwent two cycles of adherence to plastic. Typically, endothelial colonies appeared between days 12 and 25 and between days 18 and 36 in cultures of the adherent and nonadherent CD45−FIB−CD34+ cells, respectively. Importantly, the CD45−FIB−CD34− cells also gave rise to endothelial colonies that appeared in the culture at similar times, between days 12 and 25 in cultures of adherent cells and between days 18 and 50 in cultures of nonadherent cells (Fig. 4D). Given the late appearance of endothelial colonies, their cobblestone morphology, and the absence of CD45+ cells, we conclude that these colonies represent late-outgrowth endothelial progenitors that are also known as endothelial colony-forming cells (ECFCs) [32,33] (Fig. 4C). The ECFC frequency in the adherent cells was very similar in both cell populations, yielding an average of three or four ECFCs per 100,000 cells. In contrast, the nonadherent CD45−FIB−CD34+ cells were highly enriched in the ECFCs, with a mean value of ∼10 ECFCs per 100,000 cells, a frequency 11-fold higher than that exhibited by the nonadherent CD45−FIB−CD34− cells (Fig. 4D). Considering that in the process of immunomagnetic selection, the recovered CD45−FIB−CD34+ and CD45−FIB−CD34− cells represented ∼89% and 11%, respectively, of the CD45−FIB− labeled cells and that the number of nucleated cells per gram of adipose tissue was 1.27×106±0.55×106 cells, we estimated a yield of ∼19 ECFCs per gram of tissue, of which only 4.7% of the ECFCs are derived from CD45−FIB−CD34− cells.
ECFC-derived endothelial cells are functional
To characterize the ECFC-derived endothelial cells generated in the CD45−FIB−CD34− cell cultures, when colonies grew to cover almost the entire surface of the wells, the cells were detached and subcultured on uncoated plastic culture dishes. The cells displayed the typical endothelial cell morphology, and they expressed CD34, CD31, KDR, Tie2, CD144, and CD146 (Fig. 5A). Then, for analysis at the molecular level, and specifically, to evaluate the progressive changes in the expression of genes involved in the differentiation of the endothelial lineage, the CD45−FIB−CD34− cells that were not adhered to the plastic were seeded on fibronectin/collagen-coated dishes. At different days of culture, the cells were harvested and processed for reverse transcription-polymerase chain reaction (RT-PCR), and the gene expression levels relative to the levels in primary endothelial cells derived from ECFCs arising in the CD45−FIB−CD34+ cell cultures were calculated. At the beginning of culture, the CD34, CD31, Tie2, and SCL/TAL1 mRNA levels were undetectable, whereas KDR and Endoglin expression was minimally detected. At 7 days of culture, the CD31, KDR, and Endoglin mRNA levels were already detected, and the transcript levels continued to increase, reaching levels similar to those exhibited by primary endothelial cells derived from CD45−FIB−CD34+ cells. Interestingly, after 28 days of culture, the KDR and SCL/TAL1 mRNA levels were elevated up to 3.5- and 7- fold, respectively (Fig. 5B), suggesting that endothelial cells derived from CD45−FIB−CD34− must be more immature than those derived from CD45−FIB−CD34+ cells.
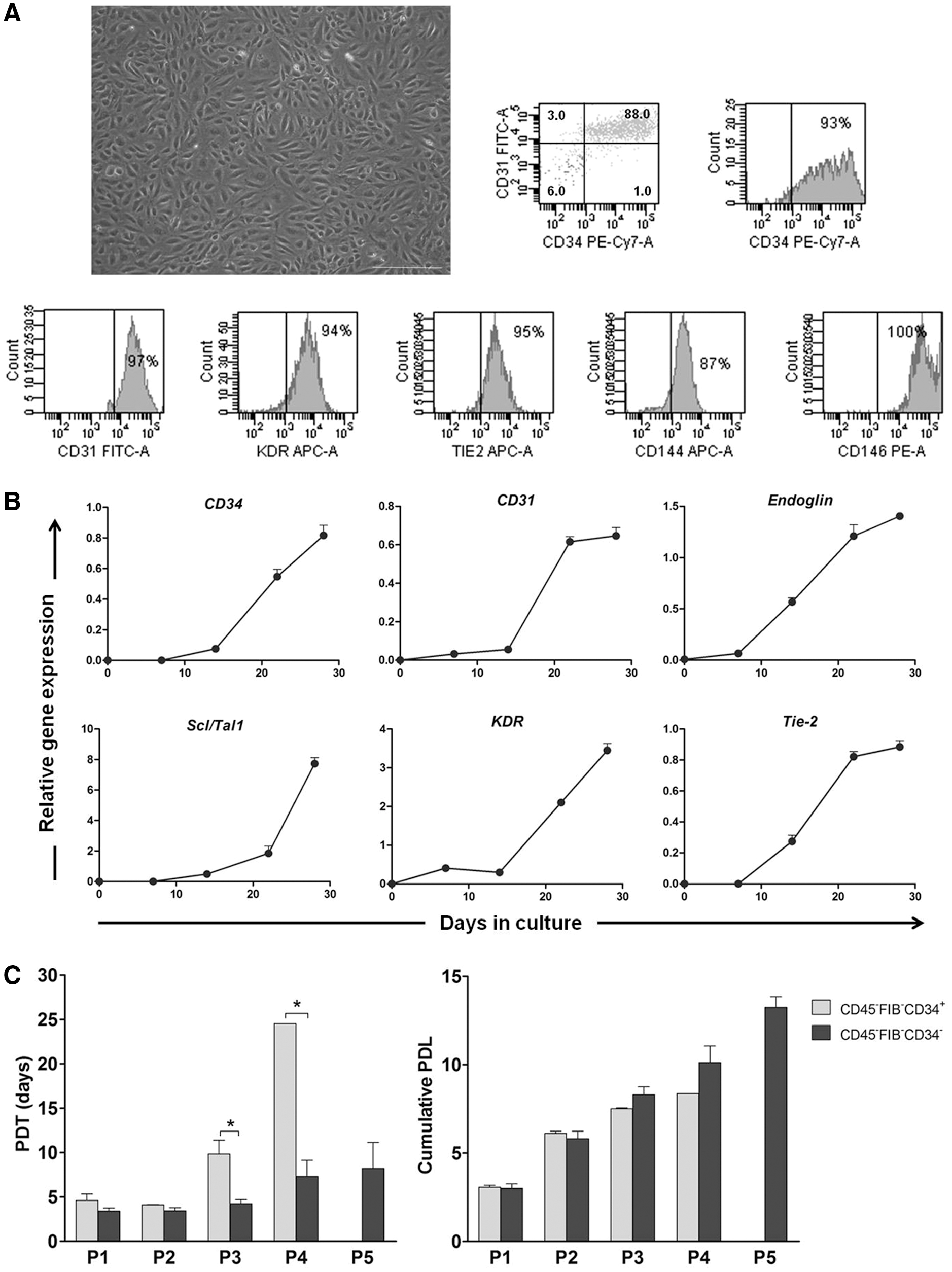
Phenotypic characteristics and kinetics of proliferation of ECFC-derived endothelial cells.
Next, the proliferation kinetics of ECFC-derived endothelial cells was determined. We observed that endothelial cells derived from the CD45−FIB−CD34+ cell population proliferated with a PDT of ∼4 days for the first and second passages, increasing up to 10 and 25 days by the third and fourth passages, respectively, and then reaching senescence. By contrast, endothelial cells derived from the complementary CD45−FIB−CD34− cell subset proliferated with a significantly lower PDT (∼4, 7 and 8 days for the third, fourth and fifth passages, respectively) (Fig. 5C). Nevertheless, endothelial cells derived from either SVF cell population completed ∼10 PDLs by the fourth passage (Fig. 5C). Again, these results suggest that ECFC-derived endothelial cells arising from the CD45−FIB−CD34− cell populations must be more immature than those developing from the complementary CD34+ cell subset. The functionality of endothelial cells derived from the CD45−FIB−CD34− cell population was also determined. As shown in Fig. 6A, when cells were seeded on Matrigel-coated dishes, they began to align into tube-like structures that were stable for several days. Additionally, the cells expressed markers that are characteristic of endothelial cells, vWF and e-NOS, and they also took up Dil-Ac-LDL (Fig. 6B). Moreover, in an in vivo system, such as the subcutaneous implantation of Matrigel containing ECFC-derived endothelial cells into nude mice, we observed the formation of functional vascular structures, as demonstrated by the presence of erythrocytes within the structures (Fig. 6C). To assess whether ECFC-derived endothelial cells contributed to the formation of new blood vessels, we used UEA-1 to stain the endothelial cells of human origin and rat anti-mouse CD31 that recognizes murine endothelial cells. Double immunofluorescence demonstrated the formation of vascular structures with human and murine endothelial cells co-localizing in the newly formed blood vessels (Fig. 6D).

In vitro and in vivo functionality of endothelial cells derived from ECFCs arising in CD45−FIB−CD34− cells.
CD45−FIB−CD34− cells in the SVF are enriched in mesodermal precursors
Daily observation of the cultures allowed us to determine that although most CD45−FIB−CD34− cells that were not adherent to the plastic exhibited adhesion to a coating of fibronectin and collagen, a small proportion of cells remained floating in the culture medium for the duration of the experiment. Gene expression analysis showed that these nonadherent cells lacked CD34, CD31, Endoglin, KDR, Tie2, and SCL/TAL1 expression (data not shown); however, after 22 days of culture, the Nanog, Oct-4, and Sox2 mRNA levels were increased relative to their levels in CD45−FIB−CD34− cells at day 0, demonstrating the stemness properties of these cells (Fig. 7A). Second, it is important to highlight the fact that in some cultures of adherent CD45−FIB−CD34− cells, in addition to the endothelial colonies, colonies of adherent spindle-shaped cells that resembled primitive stromal cells were also generated (Fig. 7B). Although we did not determine the date of emergence or the frequency of such colonies given their scarcity, we observed that their growth rate was similar to that of endothelial colonies. Taken together, these findings suggest that CD45−FIB−CD34− cells contain mesodermal precursors endowed with endothelial and stromal potential. To further assess this possibility, freshly isolated CD45−FIB−CD34− cells were cultivated using stem cell culture medium to prevent cell differentiation under normoxic (20% O2) or hypoxic (5% O2) conditions. Spindle cell colonies generated in culture (Fig. 7C) rapidly proliferated, and after 20 days of culture, the total number of cells was increased approximately by five-fold. No differences in morphology or proliferation rate were observed during the first three subcultures between cells that were grown in mTeSR1 or mTeSR2 medium under normoxic or hypoxic conditions. Immunocytochemical staining showed that the cultured cells expressed the human stem cell markers SSEA-3, Tra-1-60, Tra-1-81, Oct4, and Sox-2; furthermore, cells expressing CD31 were also detected (Fig. 8). FACS analysis showed that CD45−FIB−CD34− cells gave rise to two well-defined cell populations with different FSC and SSC characteristics that exhibited different phenotypes (Fig. 7D and Table 3). The smaller cells (low FSC, depicted in red in Fig. 7D) comprised 58.5%±11.0% of the total cells under normoxic conditions, but only 8.9%±5.5% under hypoxia. Regardless of the culture conditions, almost all the cells contained in the high FSC population expressed CD90, CD9, CD13, and CD105. Additionally, the expression of CD143/BB9, SSEA4, and CD146 was practically restricted or was predominantly restricted to cells with high FSC. By contrast, CD34 and CD31 expression was enriched or restricted, respectively, on cells with low FSC. Moreover, it is noteworthy that under normoxic conditions, only between 1% and 2% of cells with high or low FSC, respectively, expressed the Tie2 antigen, whereas under hypoxic conditions, the percentage of Tie2-expressing cells was 4% and 17%, respectively. Additionally, KDR expression was undetectable on cells cultured under normoxia, but it was minimally expressed under hypoxic conditions (Table 3). Moreover, under these culture conditions, no CD45+ cells were generated. Together, these results support the hypothesis that the SVF fraction contains FIB−CD34− mesodermal precursors with mesenchymal and endothelial potential.

The SVF CD45−FIB−CD34− cell population contains primitive mesodermal precursors.
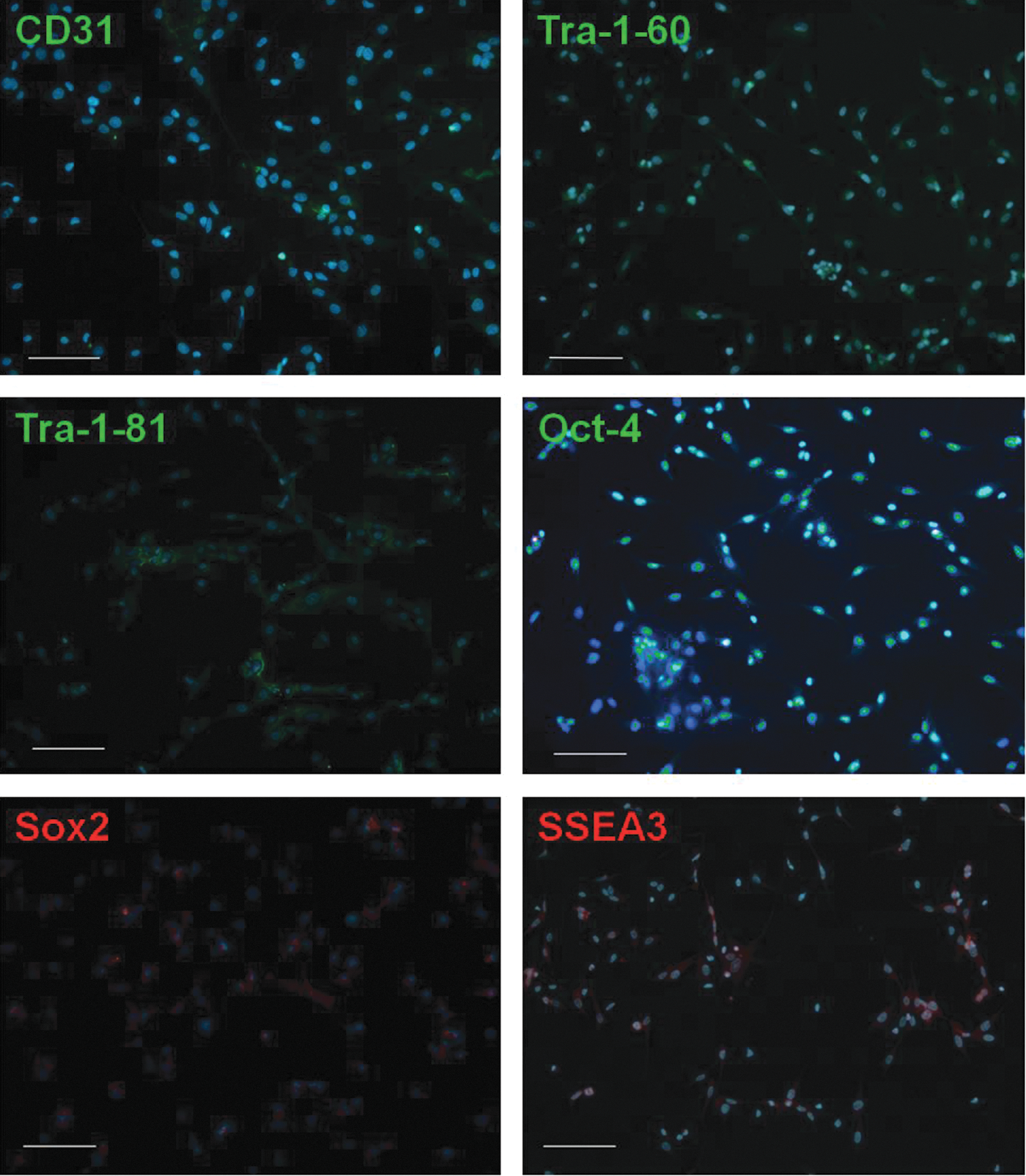
Immunocytochemistry of CD45−FIB−CD34− cells cultured in stem cell medium. The figure shows representative photomicrographs of cultured CD45−FIB−CD34− cells at P1 immunostained for the stem cell markers Tra-1-60, Tra-1-81, Oct-4 (green), Sox2, and SSEA3 (red) or for the endothelial marker CD31 (green) and then counterstained with DAPI (blue). Merged images are shown; scale bar: 100 μm. DAPI, 4′,6-diamidino-2-phenylindole dihydrochloride.
Freshly isolated CD45−FIB−CD34− cells from the SVF of human adipose tissue were cultured under normoxia (20% O2) or hypoxia (5% O2) in mTeSR1 or mTeSR2 media. Semi-confluent cells were harvested by mild trypsinization, and then they were analyzed by flow cytometry. The values indicate the percentage of cells expressing the selected antigen within the gate (Low FSC or High FSC). The results are expressed as the mean±SD from four independent experiments with SVF cells derived from four different donors. For comparison, the values obtained for cells with low FSC cultured in normoxia were compared with those obtained in hypoxia, and values for cells with high FSC cultured in normoxia were compared with those obtained in hypoxic conditions.
Significant difference, a P<0.01.
Nd, not determined.
Discussion
In the present study, we show that ∼85% of the SVF CD45− cells were bound to fibroblast microbeads, and this positive cell fraction contained 90%–95% of the nonhematopoietic CD34+ cells. Similar to FIB+ cells from the bone marrow [25,26], the CD45−FIB+ cells from the SVF were adherent to plastic and gave rise to fibroblastoid cells. Depletion of FIB+ cells allowed us to study the nonstromal CD34+ and CD34− cells. Consequently, we observed that the plastic nonadherent CD45−FIB−CD34+ cells were highly enriched for ECFCs. These results are consistent with a recent study [34] that demonstrated the existence of ECFCs among isolated CD34+CD31+ cells from the SVF of human adipose tissue.
It has been reported that the hemangioblast and the hemogenic endothelium are derived from an Flk1/KDR mesodermal precursor [35 –37]. We previously reported the existence of a rare population of CD45−KDR+CD34+/− cells within the SVF of human adipose tissue that exhibited hemangioblastic properties in vitro [18]. Here, we show that the CD45−FIB−CD34− and CD45−FIB−CD34+ cell populations contained KDR+ cells, although at very low levels. Additionally, genes associated with hemangioblast induction and differentiation [37 –39] were expressed in both cell populations, but unexpectedly, the KDR and GATA-2 mRNA levels were significantly higher in the CD34− cell compartment. Similar to the CD45−FIB−CD34+ cells, the CD45−FIB−CD34− cells when cultured in EGM, a medium that favors and supports endothelial cell growth, also gave rise to ECFCs, and the ECFC-derived endothelial cells were incorporated into new blood vessels in a model of in vivo angiogenesis. We eliminated the possibility that the endothelial potential exhibited by FIB−CD45−CD34− cells was due to contamination by CD34+ or CD31+ cells because no CD34 or CD31 expression was detected by flow cytometry or RT-PCR analysis. However, ∼10%–20% of the small CD34− cells were always collected in the CD45−FIB−CD34+ cell fraction. A rare population of CD34−Lin−CD45−CD133− cells endowed with the ability to differentiate into hematopoietic and endothelial cells has been reported to exist in human peripheral blood after long-term coculture with murine stromal cells [40]. However, using culture conditions similar to those employed in this study, these cells did not exhibit endothelial potential [40], and therefore are expected to be different from those present in the SVF of adipose tissue. It has also been reported that CD34− cells, isolated from cord blood, generated endothelial cells, but this cell population contained CD45+ and CD31+ cells [41]. Plasticity of adipose-derived MSCs has been widely described in the literature [42,43], but from our point of view, it is important to make a distinction between the endothelial potential of the CD45−FIB−CD34− cells described in this study and plasticity toward endothelial cells obtained by the differentiation of adipose-derived MSCs [44,45]. Moreover, the results show that a hierarchy of endothelial cells, comprising mesodermal precursors with endothelial potential, ECFCs and mature endothelial cells, exists in the adipose tissue.
Additionally, when cultured in EGM, the plastic-adherent CD45−FIB−CD34− cells also gave rise to stromal colonies, suggesting the existence of a mesodermal precursor of stromal cells in this cell population. It has been proposed that pericytes are the ancestors of MSCs in postnatal tissues [9,46] because pericytes isolated from human fetal and adult tissues, including white adipose tissue, give rise to MSCs in culture; however, these perivascular cells were identified and isolated by high CD146 expression and a lack of CD45 and CD34. Given that the SVF CD45−FIB−CD34− cells, unlike pericytes, lacked the expression of CD146, they identify a distinct mesodermal precursor. Reyes et al. [47] identified multipotent adult progenitor cells (MAPCs) in human bone marrow in the CD45−GlyA− cells that exhibited differentiation into multiple mesodermal lineages, including endothelial cells [48], when cultured in the appropriate culture medium. Our results do not allow us to ascertain whether mesodermal precursors within the CD45−FIB−CD34− cell population that give rise to stromal colonies represent a population similar to MAPCs, but the possibility that there is a relationship with this lineage cannot be excluded.
Interestingly, under stem cell culture conditions, comprising the use of a qualified extracellular matrix, and a culture medium highly enriched in growth factors that promote and support the maintenance and long-term culture of human embryonic and pluripotent stem cells, CD45−FIB−CD34− cells gave rise to all the phenotypic heterogeneity present in freshly isolated SVF cells, including CD34 and CD31 expression. Thus, these results demonstrate the existence of different stem and progenitor cells residing in the CD45−FIB−CD34− cell population. In addition, cells arising in culture expressed some of the well-known stem/pluripotency markers suggesting that CD45−FIB−CD34− cells represent a very primitive cell population, or at least contain very primitive cells, as evidenced by the relatively high expression of Nanog, Oct-4, and Sox-2 in the nonadherent cell population. Recently, a new population of pluripotent stem cells termed Multilineage Differentiating Stress-Enduring (Muse) cells has been identified in human bone marrow, fibroblasts [49], and adipose tissue [50]. These cells are isolated using severe cellular stress conditions, are positive for the mesenchymal marker CD105 and express pluripotency markers including SSEA3, TR-1-60, Oct3/4, Nanog, and Sox2. Moreover, they differentiate into cells of the three germ layers [49,50]. Although the ability of the CD45−FIB−CD34− cell population to differentiate into cells of the different lineages has not been determined, the results presented in this study support the notion that Muse-adipose cells must be contained therein.
Using human embryonic stem cells directed toward mesendodermal differentiation, Vodyanik et al. [51] identified two different colonies, blast colony-forming cells, or hemangioblast colonies, and mesenchymal colony-forming cells, which they termed mesenchymoangioblasts, as they exhibited mesenchymal and endothelial potential. It is not known whether the mesenchymoangioblasts that occur with hemangioblasts give rise to mesenchymal and endothelial precursors at a later stage of differentiation. However, pending further progress in the knowledge of the adipose CD45−FIB−CD34− cells; it is very tempting to hypothesize, according to the theory of Bianco and Cossu [52] that these precursors or perhaps other primitive mesodermal precursors could persist in adult life. Because adipose tissue is the only organ that can grow throughout life and angiogenesis is required for expanding adipose tissue, these mesodermal precursors could meet the needs of the tissue.
Conclusions
In this study, we identify, for the first time, a cell population within the SVF of adipose tissue that contains mesodermal precursors of mesenchymal and endothelial cells. These findings offer an opportunity to investigate the cellular and molecular pathways for the development of endothelial cells in the adult. Furthermore, this cell population could provide a cell source for tissue engineering and regenerative medicine applications.
Footnotes
Acknowledgments
This work was supported by grants from Ministerio de Ciencia e Innovación (PI11/01242) and Ministerio de Sanidad y Consumo (EMER07/005). We thank personnel from the Immunohematology Laboratory of Centro de Transfusiones, and Elena Alvarez of the Regenerative Medicine Laboratory for their technical assistance.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.