
Abstract
Regenerative medicine is in demand of adult pluripotent stem cells (PSCs). The “Bonghan System (BHS)” was discovered and suggested to contain cells with regenerative capacity in the early 1960s. It had been ignored for a long time due to the lack of sufficient details of experiments, but about 37 years after the initial report, the BHS was rediscovered and named as the “primo vascular system.” Recently, we have discovered a similar structure, which contained a high level of hyaluronic acid, and hence, named the structure as hyaluronic acid-rich node and duct system (HAR-NDS). Here we discuss the HAR-NDS concept starting from the discovery of BHS, and findings pointing to its importance in regenerative medicine. This HAR-NDS contained adult PSCs, called node and duct stem cells (NDSCs), which appeared to circulate in it. We describe the evidence that NDSCs can differentiate into hemangioblasts that further produced differentiated blood cells. The NDSCs had a potential to differentiate into neuronal cells and hepatocytes; thus, NDSCs had a capability to become cells from all three germ layers. This system appears to be a promising alternative source of adult stem cells that can be easily delivered to their target tissues and participate in tissue regeneration.
Introduction
R
Several studies have provided evidence for the presence of adult stem cells in tissues or organs [6,7], such as mesenchymal stem cells (MSCs) [8], multipotent adult progenitor cells [9], marrow-isolated adult multilineage inducible cells [10], multipotent adult stem cells [11], endothelial progenitor cells [12], very small embryonic-like stem cells (VSEL) [13], endogenous plastic somatic cells [14], neural (N) stem cells, mammary stem cells, intestinal stem cells, inner ear stem cells, and testicular stem cells [15]. These cells exist at different levels of specification and may have broader differentiation potential [16].
In 1962, Kim discovered a new anatomical system, the Bonghan System (BHS) [17], and proposed the Bonghan theory [18]. In subsequent studies, he reported the presence of Sanal (meaning live tiny egg) with self-regenerating property. Interestingly, he reported that these sanals, while flowing in the BHS, divide to differentiate into several types of tissue-committed cells in vivo as well as in vitro. He also reported hematopoiesis and regeneration of damaged liver tissues by the sanals [19 –21]. Since the capacity to regenerate and the ability to differentiate into different types of cells are the essence of a stem cell, sanals are thought to be pluripotent, and the BHS to be a store of adult stem cells. Unfortunately, as the properties of sanals were not well characterized by Kim and the concept of stem cell/adult stem cell was not familiar at that time, the hunt for PSCs in the BHS remained unsucessful. Moreover, despite the efforts to confirm Kim's findings [22–23], the Bonghan Theory has been largely dismissed for about 37 years, due to the lack of a detailed procedure for the isolation/identification of sanals/BHS. Later, Soh and his research team confirmed the existence of BHS and it was renamed as the primo vascular system (PVS) [24]. Recently, we discovered a similar structure that might not be the same as BHS and therefore put forth a new name, hyaluronic acid-rich node and duct system (HAR-NDS), and demonstrated that the HAR-NDS contains hematopoietic progenitor cells (HPCs) and pluripotent stem cells [25,26]. We also reported the presence, in the HAR-NDS, of the node and duct stem cells (NDSCs), which appear to be a promising candidate for use in regenerative medicine [27].
In this review, we will describe the discovery of HAR-NDS in three main parts. In the first part, we describe Kim's findings related to discovery and characterization of BHS since most of them were published as book chapters in the Korean language. In the next section, we will discuss the findings of Soh et al. and others repeating and confirming Kim's observation, as well as elucidating the physiological role of PVS. Finally, we will discuss our studies relating to HAR-NDS characterization and the discovery of NDSCs. The alternative names used by different groups for similar structure in different parts of the BHS/PVS/HAR-NDS are shown in Table 1.
HAR-D, hyaluronic acid-rich duct; HAR-N, hyaluronic acid-rich node; HAR-NDS, hyaluronic acid-rich node and duct system; HAR-L, hyaluronic acid-rich liquor; NDSCs, node and duct stem cells.
The BHS: Discovery and Characterization of New Anatomical System (1963–1965)
In 1962, Kim reported his first work dealing with the discovery of new anatomical entities corresponding to the ancient acupuncture meridians (AM). In his report, for the first time, he made a potential connection between the physiological responses at the acupoints to an actual organ [17]. In his second report, Kim claimed the discovery in rabbit of a completely new system (BHS), consisting of node-like structures termed Bonghan corpuscles (BHC) at the acupoints and tube-like structures or Bonghan ducts (BHDs) on the skin, body surface, as well as in internal organs, including blood vessels and lymph vessels [18]. In 1965, he submitted another three reports that describe the anatomical and histological features of the BHS as well as its basic physiological aspects [19 –21]. He also reported that hematopoietic organs, such as bone marrow (BM), spleen, and lymph nodes, have a well-developed BHS that may function as novel hematopoietic organs [21].
In this section, we have included important findings of Kim's report “Kyungrak System and Theory of Sanal” published in the proceedings of the Academy of Kyungrak of the Democratic People's Republic of Korea in 1965, which is available in English. Kim's findings can be summed up in two parts: the Bonghan theory and the sanal theory.
The Bongahan Theory
According to Kim, the BHS is composed of two subsystems, BHDs and BHCs, and the Bonghan liquid flowing inside the BHDs and BHCs. All the BHCs are interconnected through BHDs. BHDs connect BHCs.
Kim demonstrated that the BHD consists of a bundle of very fine Bonghan ductules with a diameter of 5–10 μm. The wall of the ductules (0.1–0.2 μm) is formed of a single layer of endothelial cells, with rod-shaped nuclei with pointed ends (15–20 μm in length) and abundant homogeneous chromatin. The outer membrane of ductules consists of spindle-shaped smooth muscle-like cells with centrally located ellipsoid nuclei (13–27×4–5 μm). Fine chromatin granules are found in the nucleus, and the nuclear membrane is clearly distinguished. The outer membrane is further surrounded by fine, longitudinal, and circular fibers crossing each other. These ductules contain basophil granules of various shapes and small nucleus-like structures. Fibrous structure and amorphous substances exist between ductules. All the ductules are wrapped together into a bundle by a thin membrane (periductium, peri-BHD, Fig. 1A), bearing cells with spherical or oval-shaped nuclei (6–12 μm in diameter) and abundant cytoplasm. Cells in the periductium are very rare. Based on its location, BHDs are classified as internal/intravascular (IV), intraexternal/organ surface (OS), external/extravascular (EV), or neural (N) BHD.

The Bonghan System (BHS) and flow of fluid through the BHS.
According to Kim, BHCs may be oval, spindle-shaped, or some other shape (Fig. 1B) and their size varies from 0.1 to 3.0 mm in diameter. They are surrounded by loose connective tissue and by a dense network of blood vessels. The outer layer consists of a thin outer circular layer and thick inner longitudinal layer of smooth muscle-like cells. The BHC is mainly formed by dilated, ramified, and anastomosed ductules (sinus), and the main structural components of the BHC are the reticular elements of the outer membrane, interstitial substances of the ductules, and the periductium. The BHCs contain cell elements, chromaffin granules, and basophil structures, as well as small structures resembling nuclei. BHCs are classified as superficial (skin) or profound. The profound BHCs are further classified as internal/intravascular (IV), intraexternal/organ surface (OS), external/extravascular (EV), or neural (N) and intraorganic BHCs.
According to Kim Bonghan, liquor contains abundant DNA and RNA, protein, lipid, reduced sugar, free amino acids, mononucleotides, hyaluronic acid, adrenocortical hormone, adrenomedullary hormone, and sex hormone (estrogen) (Table 2). The DNA of Bonghan liquor is AT rich. Free mononucleotides present in Bonghan liquor are cytosine monophosphate, nicotinadenine dinucleotide, adenosine monophosphate, inosine monophosphate, guanosine monophosphate, cytosine diphosphate, uridine monophosphate, adenosine diphosphate (ADP), ADP ribose, cytosine triphosphate, uridine diphosphate (UDP), UDP-hexosamine, guanosine diphosphate, adenosine triphosphate, guanosine triphosphate, and uridine triphosphate. Besides these, it also contains spherical-shaped (occasionally oval) microgranules, which Kim calls “sanals” (Fig. 1C).
Taken from Kim's report “Kyungrak System and Theory of Sanal” published in the proceedings of the Academy of Kyungrak of the Democratic People's Republic of Korea in 1965.
Overall, the BHS is said to consist of the following: (1) The internal/intravascular (IV) BHS: This comprises IV-BHDs and -BHCs. It is free floating and distributed systemically in all the blood and lymphatic vessels and, thus, follows the entire path of the blood/lymphatic vessels and is delivered to the organs (Fig. 2A). Each blood vessel has one or more IV-BHD. Interestingly, the IV-BHDs are claimed to pass in and out of the blood/lymphatic vessels through their walls. They are extremely soft and glutinous and their periductium and interstitial substance are poorly developed. The IV-BHCs have a structure remarkably resembling that of a hematopoietic organ, and myeloid as well as lymphoid cells are found in the network of reticular tissues, suggesting that hematopoiesis is one of the important functions of the internal BHS. (2) The intraexternal/organ surface (OS) BHS: This consists of the OS-BHDs and OS-BHCs. It runs independently of the blood and lymphatic vessels and is distributed all over the body: on the surfaces of internal organs and in the thoracic and abdominal cavity, and it spreads reticulately, sending out branches to various organs (Fig. 2B). Interstitial substances and periductium are more developed in the OS-BHD than in the IV-BHD. The BHD sinus of the OS-BHC contains cells with a pale cytoplasm as well as basophilic structures. (3) The external/extravascular (EV) BHS: This mainly runs along the walls of blood and lymphatic vessels. The EV-BHDs sometimes run independently of blood vessels or along nerves (Fig. 2C) and occassionally contain the nucleus-like structure. The EV-BHDs connecting the superficial BHCs are called superficial BHDs. The periductium of the superficial BHDs is described as composed of connective tissue in which there are numerous blood capillaries. Many chromaffin granules are seen in the lumens of the BHDs and inside the BHCs. (4) The neural (N) BHS: This part of the BHS is composed of N-BHDs and N-BHCs that are distributed throughout the central and peripheral nervous system (Fig. 2D). In the central nervous system, they are said to be present in the brain and spinal cord, running in a free state through the central canal, cerebral ventricles, and subarachnoid space along the route of circulation of the cerebrospinal liquor. The N-BHD, consisting of two to four ductules, is semitransparent, faintly milky white, and of very soft consistency. (5) The intraorgan (IO) BHS: Inside the organs there are IO-BHCs, terminal BHCs, and terminal BHDs (Fig. 2E). These are part of the IV, EV, and N-BHDs. The IO-BHCs are located in the internal and other organs, measure 0.1–0.5 mm, and appear more compact than the surrounding tissue. They are described as the smallest of the BHCs and have the simplest structure containing granular and filiform basophilic structures. The BHDs leaving IO-BHCs ramify and reach a number of minute IO-BHCs called terminal BHCs. There are numerous terminal BHCs in the organs. A fine duct leads out of each terminal BHC and is called terminal BHD. The terminal BHD is thinner and reported to be filled with Feulgen-positive granules that enter the cytoplasm, penetrating the cell membrane and the karyoplasm through the nuclear membrane. Finally, Kim said that the BHD is connected directly with the nucleus where it seems to form a network.
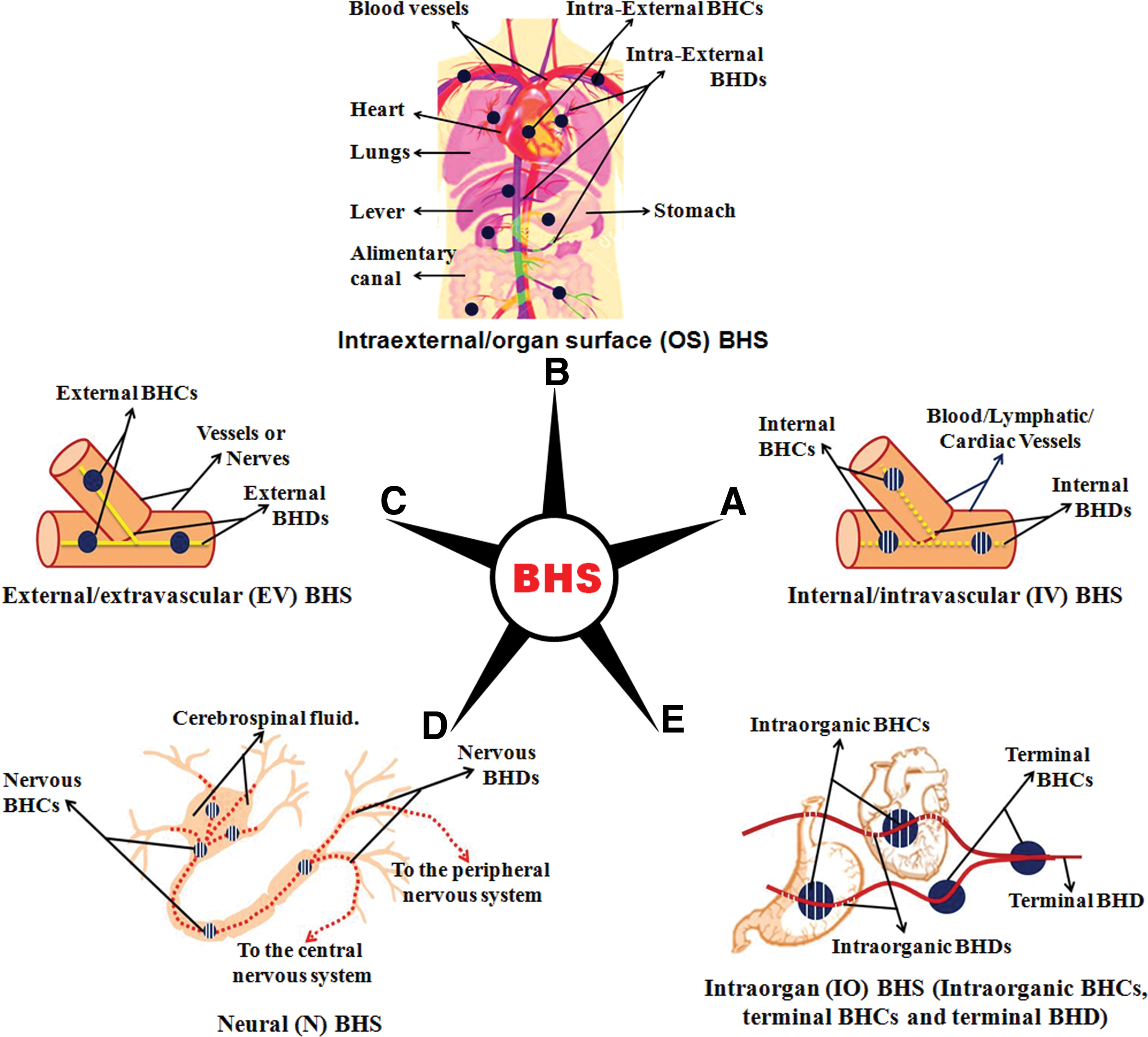
As described by Kim, the different subclasses of BHS are linked with each other, and all the cells are connected to BHS. Moreover, all the tissue cells are provided with fine terminal ductules penetrating into the cell nuclei and exiting from them. These ductules are linked with IO-BHCs, which in turn are linked only to tissues within a certain range. The IO-BHCs are linked with all the BHS, and the IV-BHS is linked to the OS-BHS through the vessel walls and to the EV-BHS through the EV-BHCs. The OS-BHS is connected to the EV-BHS and the N-BHS.
The BHS is said to possess bioelectrical activities, excitatory conductivity, and mechanical motility. The electrical changes in the BHDs are very slow and when they are stimulated the electric processes form a variety of patterns as they travel along the BHD. The BHDs also make automatic movements (continuous or periodic, longitudinal or vibrating transversely) that are propagated and change when the BHD is stimulated. Stimulation of the BHDs is described as affecting the heart beat and forcing cardiac contraction, as well as changes in the movements of the intestines. Cutting the BHDs causes prominent changes such as karyolysis, apoptosis, and reduced excitability of nerves and muscle movement.
Kim asserts that the BHS is capable of actively circulating the Bonghan liquor. There are several interlinked but independent circulatory pathways for Bonghan liquor. By injecting P32 into various parts of the BHS, Kim observed that the Bonghan liquor flowed from various organs and tissues toward the superficial BHCs, and starting from superficial BHCs it circulated only in one direction from the superficial to the profound BHCs. The Bonghan liquor flowing from the profound BHCs then reaches tissue cells through the IO-BHCs. Moreover, the circulations of the Bonghan liquor in the IV-BHD and the EV-BHD are said to be linked. Thus, the liquor circulates only along specific pathways, however, it can cross to another pathway through connections between two pathways (Fig. 1D).
The differentiation and development of the BHS are said to precede those of the vascular, nervous, and other organs. Thus, Kim suggested that the BHS played an important role during the embryonic development of organisms. The BHS exists in all vertebrates and invertebrates, as well as in plants, that is, in all multicellular organisms.
Thus, according to the Bonghan theory, the BHS is made up of multiple, independent circulating systems (subsystems), which are interconnected and form a single coherent system. All organs of living organisms are connected to and believed to be controlled by the BHS.
The Sanal Theory
Kim claimed that unique 0.8–2.4 μm, spherical (occasionally oval shaped) microgranules “Bonghan sanals” circulate in the BHS of all living organisms and plants. These “sanals,” are the key functional components of the BHS and are said to consist of sanalosome (containing abundant DNA), sanaloplasm (containing RNA), and an outer thin sanal membrane. Sanals perform peculiar rotating movements and are sensitive to environmental factors such as temperature and pH. The sanalosome also move ceaselessly in the sanaloplasm. Kim reported that while moving through the BHS, the sanals grow into cells and the cells transform into scores of sanals through sanalization. This process is repeated in a continuous cycle called the “Bonghan sanal-cell cycle” (Fig. 3A) and occurs in the tissues of all living organisms. Cells are also formed by cell division, a specific form of the Bonghan sanal cell cycle called the “intracellular Bonghan sanal cell cycle,” in which the sanalization of a cell precedes its division (Fig. 3B). At each stage of this process (fusion and dissociation), the sanal undergoes ceaseless changes. A sanal inside the cell nucleus is said to differ from those outside the cell, because while moving within the nucleus it absorbs something of the karyoplasms; it then exits the cell through the cytoplasm, absorbing substances from the cytoplasm. In this way, it acquires all features of a cell.
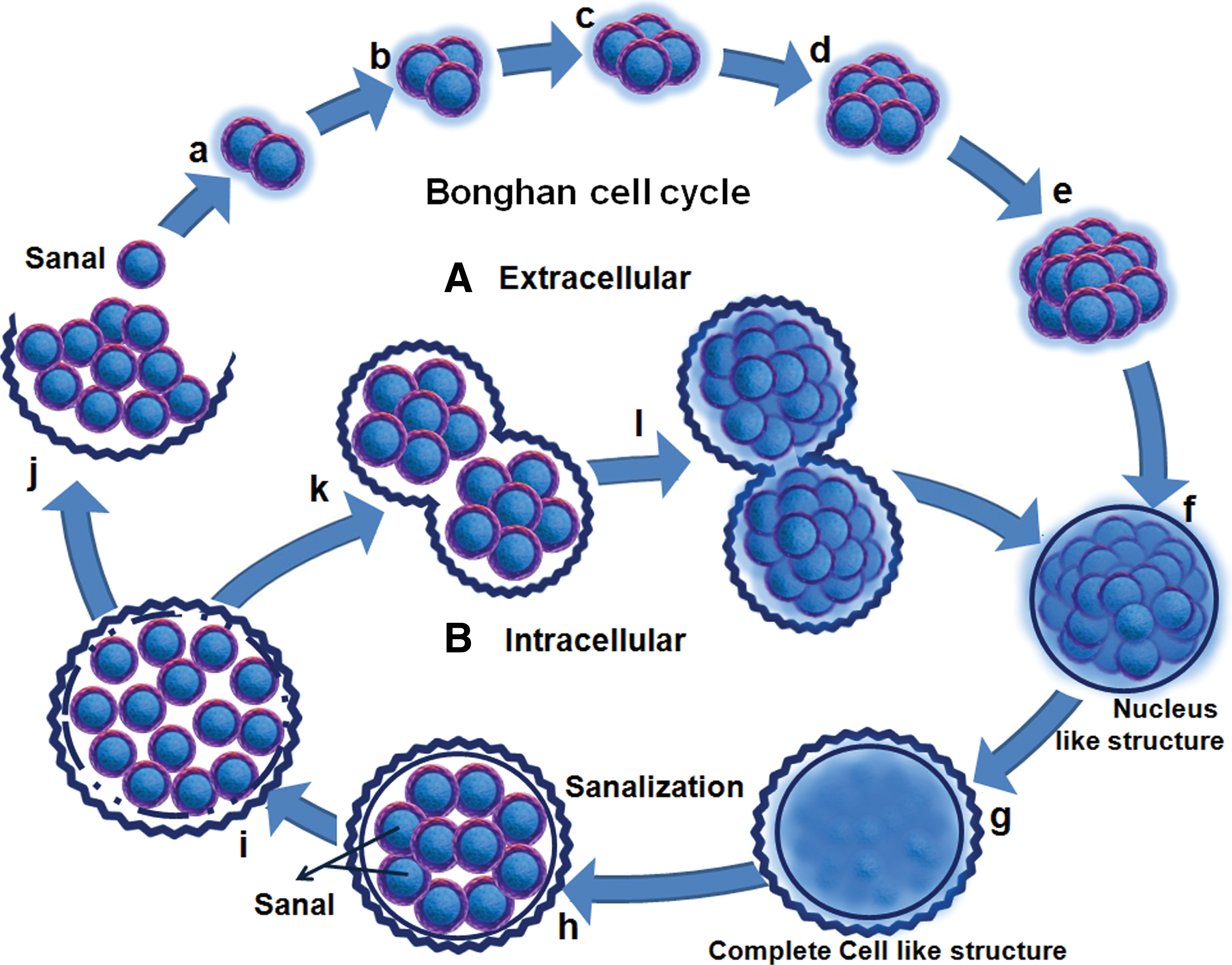
According to Kim, the cell is a specific stage of the cyclic movement of the sanal. While cell division is underway, the sanalosome appears in the form referred to as a chromosome. The quantity of DNA carried in one sanalosome is almost equal to that of DNA in one chromosome, and when sanalization takes place within the nucleus the number of sanalosomes is equal to the number of chromosomes. The picture of stained chromosomes in the metaphase of cell division is the same as that of the sanalosomes of stained sanals.
Kim further developed his view of sanals as follows: in the organism, sanals at different stages of maturity exist only in the BHDs and BHCs and are absent from blood, lymph, and tissue fluid. The Bonghan liquor contains biochemical substances necessary for the growth of sanals. Sanals grow easily into cells when they are cultivated in Bonghan liquor or in culture media of similar composition, but do not grow into cells in the blood, lymph, or tissue fluid. Sanals travel through the circuit of the Bonghan liquor, mature in it, and grow into the tissue cells of organs (lung, liver, kidney, etc.). Sanals of different stages of maturity can be seen when they are isolated from different parts of the circuit of the Bonghan liquor. When sanals from superficial BHCs are cultured, they grow into the corresponding cells. The network of BHDs in the outer layer of superficial BHCs containing abundant sanals is connected with the epidermis and the sanals circulate through the superficial BHCs. Light has an important influence on sanal numbers and movements.
Steady self-renovation takes place in all tissue cells not only at the molecular level but also at the cellular level. In other words, in an organism, there takes place unremitting metabolism as well as uninterrupted self-renewal of morphological constituents. This process can also be seen clearly in cultures of sanals. A comparison of histological preparations of normal tissue with living specimens apparently convinced Kim that many processes of the Bonghan sanal cell cycle go on normally and actively in tissue cells. The sanalization of tissue cells (2%–4% of the cells of a rabbit liver) proceeds side by side with the formation of cells from sanals (1%–3% of the cells of a rabbit liver). The process of the Bonghan sanal cell cycle can be observed more clearly in the course of regeneration of the injured tissue in which vigorous renovation takes place.
Based on his observations, Kim proposed a Theory of sanal. According to this theory
(1) All the constituent structures of the organism are steadily renewed.
(2) This self-renovation takes the form of the Bongahan Sanal-Cell Cycle.
(3) The self-renovation of the organism is performed by the BHS.
Thus, movements of sanals underlie all the phenomena of life.
The PVS: Rediscovery of BHS and Investigation of Its Physiological Role (2002 to Till Date)
The research group of Soh at Seoul National University, South Korea, has performed panoptic studies on the BHS. They showed BHS to be a semitranslucent thread-like structure (diameter of 0.1 mm) and renamed it as the PVS in 2010. The term “primo” refers to the formation of the PVS before the blood vessels or nerves. In the past 12 years, the PVS has been studied by various research groups, viewing it as a new anatomic system, separate from the blood and lymphatic systems. A variety of functions have been proposed for the PVS, based on the elements found within it.
The PVS has been observed inside blood vessels [28 –30] and lymphatic vessels [31 –36] and on the surfaces of internal organs, such as the stomach, liver, large and small intestines, bladder [37 –41], and in the brains of rabbits, rats, and mice [42 –46]. It was reported in the heart epicardium [47], mesentery [48], and spine of the rat [49]. Among the larger animals, it was confirmed in bovine heart chambers [50], mesentery surrounding the small intestine of a dog [51], and the spine of the pig [52]. Recently, human PVS was discovered on both the epithelial fascia and inside the blood vessels of the umbilical cord [53].
This system was also detected in adipose tissues, suggesting it to be a niche for MSCs, which can differentiate into adipocytes and pathways for macrophages involved in adipogenesis [54]. Studies have shown BHDs along blood vessels, lymph vessels, and nerve bundles connecting xenografted tumor tissue [39,55 –57]. In addition, intraperitoneally seeded and fluorescently labeled cancer cells were observed to infiltrate the PVS [58] and migrate through it from the primary to secondary sites during cancer progression [59]. The endogenous formation of a PVS in blood vessels has also been reported [60]. Conversely, Islam et al. reported the presence of a tumor-derived PVS in a murine xenograft. The cells obtained from this PVS yielded a striking predominance of human gene signatures and expressed human stem cell-specific transcription factors (KLF4), suggesting that the primo vessels associated with the xenografted tumor may provide a safe haven for a select population of cancer stem cells [61]. Since primo vessels (PVs) are known to constitute a circulatory system corresponding to AM or collaterals, the PVS has been suggested to be a novel pathway for metastasis in addition to blood or lymph vessels [57,62,63]. Furthermore, the PVs in the mesentery were shown to have a structure transitional between the endothelium and mesenchymal, which again supports its role in cancer [64]. Hong et al. have suggested a role for the PVS in oxygen transport [65].
The presence of multiple sub-PVs within a PV has also been confirmed. The PV's outermost layer was shown to contain large pores, and the sub-PVs were observed to have a thin wall consisting of endothelial cells with characteristic rod-shaped nuclei [37,66,67]. The corpuscles were found to be cucumber shaped with an average length of about 2 mm and a thickness of about 400 micron, and to be surrounded by a thin membrane [68]. Transmission electron microscope (TEM) images disclosed disordered collagen fibers forming the extracellular matrix of the BHS and various types of immune cells such as macrophages, mast cells (MCs), and eosinophils [67 –69]. Lim et al. also reported that both PVs and primo nodes were densely filled with WBCs (90%), RBCs (5.9%), and putative MCs (3.8%) [70].
Several studies have established unidirectional flow of primo liquid (0.3±0.1 mm/s speed) for up to 12 cm through the PVS, due to the peristaltic motion of the PVS itself [69,71,72]. Proteomic analysis of tissue and liquid from BHDs on the surface of rabbit intestines showed the presence of various proteins (keratin 10 and epithelial marker protein; EMP-3 on the epithelial cell of the PVS’ outer surface) that are not normally found in blood, lymph, or blood vessels [73]. Kim confirmed that PVs develop earlier and differentiate more rapidly than blood vessels or nerves [74].
Using the Feulgen reaction (that specifically stains DNA), 1–2 μm-sized round DNA-containing extracellular vesicles called P-microcells/DNA vesicles (Bonghan granules) were seen in the primo lumen on the surface of rabbit liver [37]. Confocal laser scanning microscopy (CLSM) and TEM of these granules showed the presence of a small nucleus surrounded by a layer of cytoplasm and a trilaminar plasma membrane and cytoplasmic protrusions resembling pseudopodia [66]. The motion of these granules was studied by Lee [75] and was found to be influenced by light [66]. Furthermore, the microgranules or sanals were shown to grow in circular patterns to become cell-like structures representing the mitosis-free alternative pathway for cell formation [76]. As previously suggested by Kim, a budding process for cell proliferation was also shown [77,78].
Based on their characteristics, the Bonghan granules are suggested to be naturally generated microcells, a possible origin of adult stem cells, which can be transformed into undifferentiated cells [79]. The expression of stem cell biomarkers (Oct4, Nanog, CD133, Sox2, Stella, Rex1, and Klf4) on the cells isolated from the PVS confirmed them to be embryonic-like stem cells [80]. Proteomic analyses of the tissues and liquid from BHDs on rabbit organ surfaces indicated that the BHDs located on organ surfaces have roles as temporary depots of stem cells for tissue regeneration, especially MSCs, cancer cells, and differentiated myeloid cells [73,81]. The composition of extracellular matrices was also found to be consistent with the stem cell expression and BHC was suggested as a novel stem cell supplier to the organs through the connected BHD [82].
The HAR-NDS: Discovery of Adult Stem Cell (2012 to Till Date)
HAR-NDS was discovered by Kwon et al., a research team at the National Cancer Center in 2012. These workers (including the present authors) have published three articles and also have considerable unpublished data focusing on the biological function and utility of the system in human diseases and health.
In 2012, Kwon et al. [25] demonstrated the existence of a new anatomical location, microscopic node and duct system in rats, which appeared very similar to the Bonghan or PVS. We also examined the immunohistochemical properties of the node and duct system and the possible roles of resident cells in the nodes, especially those on the surfaces of internal organs. By injecting 1% alcian blue dye, we demonstrated BHC/BHD, with typical node and duct structure, inside lymphatics, between the abdominal wall and the surface of the liver and intestine. These are abundantly filled with hyaluronic acid. The nodes and ducts were covered by a layer of EMP-3-positive spindle-shaped epithelium (contrary to Kim's finding of smooth muscle-like cells) and a layer of vWF-positive, but CD31-negative, endothelium. The BHD and BHC did not stain for LYVE-1, a lymphatic endothelium marker, establishing them as a distinct system from lymphatic and blood vessls. They found that these nodes were filled with a variety of immune cells such as MCs (20%), eosinophils (16%), neutrophils (5%), histiocytes (53%), a few lymphocytes (1%), round immature cells (3%), and chromaffin cells (previously shown by Kim) producing epinephrine and norepinephrine, as well as other granule-containing cells. In culture condition, BHC-derived cells were positive for cytoplasmic Oct4 evoking the possibility of the presence of hematopoietic stem cells. Secretory granules originating from the MCs in the nodes were found to pass along ductules signifying a role for this system in the transport of secretory granules. Thus, these results confirmed the existence of a potentially closed circulatory system, whose anatomical structure and cellular content are distinct from the blood and lymph systems [25].
In the subsequent study [26], Hwang et al. renamed the node and duct system as HAR-NDS, since it is rich in hyaluronic acid and stained with alcian blue. The nodes and ducts were renamed hyaluronic acid-rich nodes (HAR-Ns) and hyaluronic acid-rich ducts (HAR-Ds), respectively (Fig. 4A). In view of the presence of pluripotent and committed stem cells within the population of round and immature cells (3%), Hwang et al. examined whether (1) hematopoietic stem and progenitor cells exist in the HAR-NDS of mice and (2) this system is capable of producing blood cells independently of the BM and blood system. Consistent with these possibilities we identified HPCs such as multipotential progenitors [colony-forming unit-granulocyte erythroid macrophage megakaryocyte (CFU-GEMM)], progenitors of granulocyte-macrophage [colony forming unit-granulocyte macrophage (CFU-GM)], erythroid cells [burst-forming unit-erythroid (BFU-E)], and mast cell progenitors (MCPs) among the cells isolated from the HAR-NDS on the intestinal surface and inside the vein. HAR-NDS inside lymphatics showed only CFU-GEMM and CFU-GM. In clonogenic assay, CFU-GEMM colonies showed basophil/megakaryocyte/eosinophil/erythroblast precursors, CFU-GM colonies showed neutrophil/macrophage precursors, BFU-E colonies showed erythroblast/erythrocyte precursors, and MCP colonies showed MC precursors (Fig. 4B, C). There were fewer of these progenitor cell colonies from HAR-NDS than from BM (Table 3, Fig. 4E). MCPs were the most abundant among the HPCs in HAR-Ns. The number of MCPs in the HAR-NDS (111±13.8/100,000) was ∼5-fold higher than in the BM (5.3±0.6/25,000) and 100-fold higher than in the spleen (1.0±1.4/100,000).
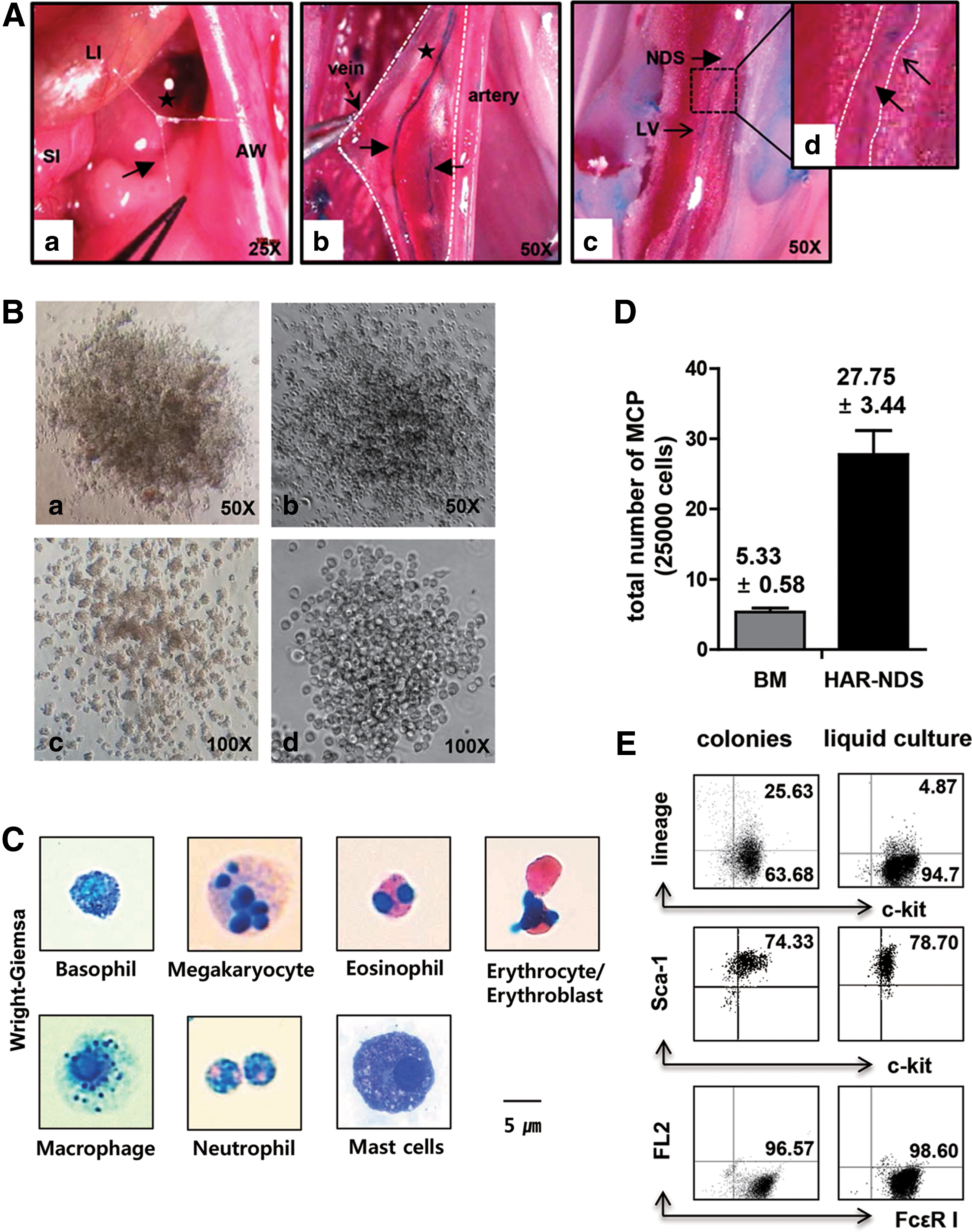
BFU-E, burst-forming unit-erythroid colonies; BM, bone marrow; CFU-GEMM, colony-forming unit-granulocyte erythroid macrophage megakaryocyte/multipotential progenitors; CFU-GM, colony-forming unit-granulocyte macrophage; MCPs; mast cell progenitors.
In addition, Hwang et al. [26] provided evidence that the HAR-NDS contains PSCs capable of generating hemangioblast-like cells on OP9 cells that subsequently generated phenotypical HSCs, functional HPCs, and differentiated blood cells. Bone marrow-derived cobblestone-area-forming cells (CAFCs) were found to form colonies during the first 7 days of culture and started to die, and no cells survived in the second passage. In contrast, CAFCs from HAR-NDS stayed undifferentiated till the fourth passage and started to differentiate on the fifth passage and to die after the fifth passage (Fig. 4D). These findings supported that CAFCs from HAR-NDS are different from the hematopoietic cells from BM. Since the HAR-NDS appears to be compactly spread throughout the body, on the surface of all internal organs, under the skin, inside the vessels, and along the nervous system, we hypothesized that this system could be a route for delivering PSCs, progenitor cells, and immune cells. However, we failed to demonstrate any long-term reconstituting or short-term radioprotective ability of HAR-ND cells on their own, perhaps due to transplantation of too few HAR-NDS-derived stem-like cells [26].
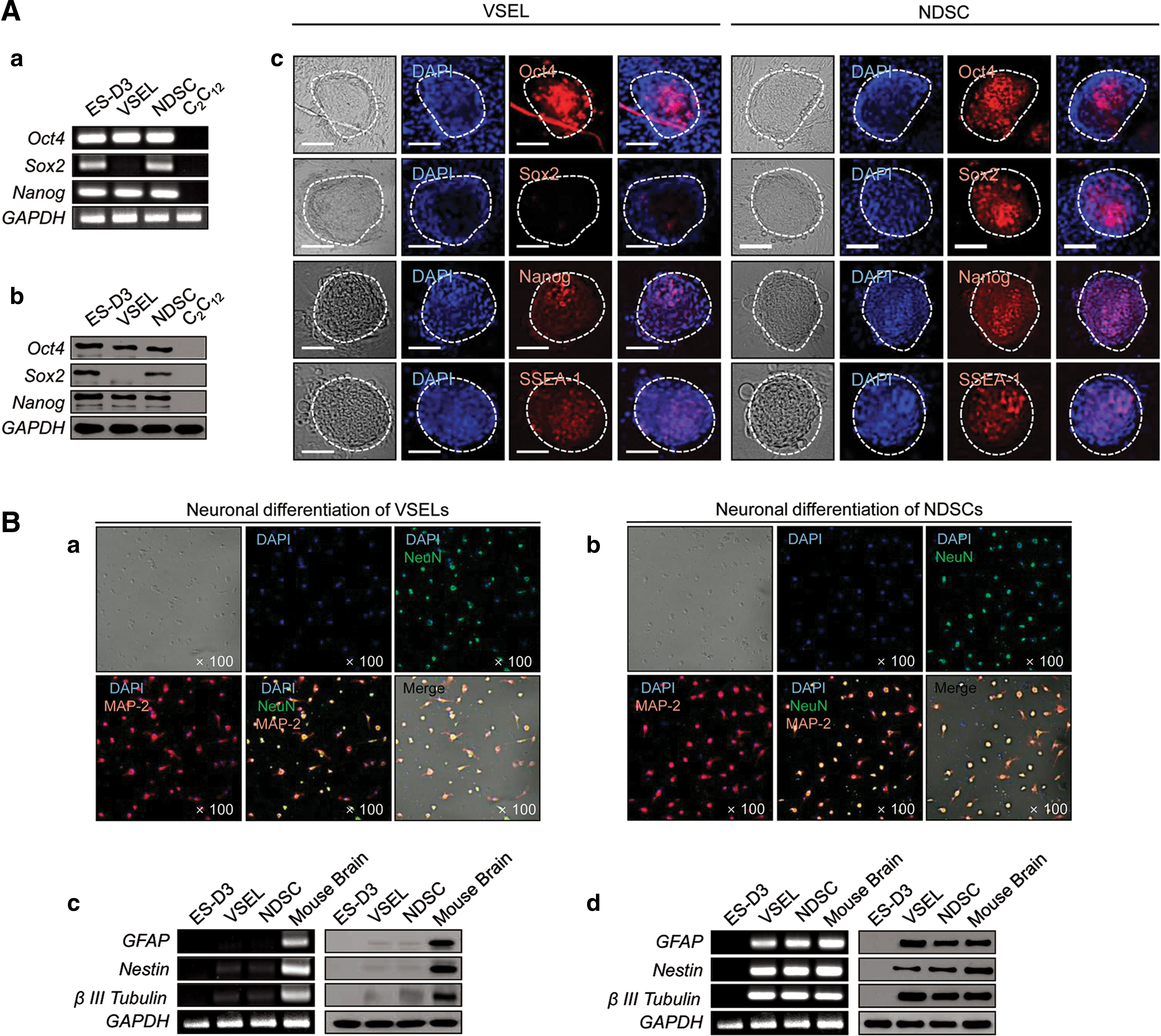
Recently, considering the very small size of sanals, we suspected the simultaneous presence of adult stem cells in HAR-NDS, similar to the VSELs previously described by Ratajczak and colleagues [13]. We were able to isolate VSEL-like cells from the HAR-NDS by employing a purification scheme commonly used to purify BM VSELs and we named them NDSCs (Fig. 5A) as they were derived from the HAR-NDS [27]. The intravascular HAR-NDS was found to contain ∼100-fold more NDSCs (∼2.32% of total HAR-NDS cells) than the BM (∼0.02% of total BM cells, Fig. 5B, C). Next, we performed a study comparing the general properties of VSELs from BM with NDSCs (Table 4). A scanning electron microscopy study showed that the NDSCs were round and small (3.5–4.5 μm in diameter). TEM confirmed the immature morphology of NDSCs (Fig. 5D). They contained a large nucleus with a narrow rim of cytoplasm (Fig. 5D-a). We also identified a prominent nucleolus (Fig. 5D-b), nuclear membrane (Fig. 5D-c), heterochromatin and euchromatin (Fig. 5D-d), mitochondria and vacuoles (Fig. 5D-e), and an endoplasmic reticulum with scattered ribosomes (Fig. 5D-f). On C2C12 murine myoblast feeder layers, the NDSCs formed spheres resembling embryoid bodies that were positive for fetal alkaline phosphatase (ALP) arguing for the presence of PSCs. Furthermore, NDSCs produced ∼176 spheres per 1,000 cells, compared with ∼14 spheres per 1,000 cells for the VSELs and the number of ALP-positive spheres originating from the NDSCs increased after repeated replating, while it remained almost constant for VSEL-derived spheres. These findings suggested better sphere-forming ability of NDSCs compared to VSELs.

Purification of VSELs from the BM and NDSCs from the HAR-NDS.
ALP, alkaline phosphatase; FACS, fluorescence-activated cell sorting; VSEL, very small embryonic-like stem cell.
In addition, we showed that NDSCs express both mRNA and protein corresponding to all three stem cell markers (Oct4, Sox2, Nanog) at similar levels to those found in a murine ESC line (ES-D3), and the NDSC spheres were also positive for Oct4, Sox2, Nanog, and SSEA-1 (Fig. 6A). Under in vitro conditions (under neuronal differentiation culture conditions for 25 days), the NDSCs differentiated into NeuN-positive, MAP-2-positive neuronal cells with an efficiency similar to that of VSELs (Fig. 6B). These NDSCs and VSELs derived neuronal cells expressed GFAP, nestin, and β-III tubulin markers (neuronal cell marker) at both mRNA and protein levels. After injecting CM-DiI-labeled NDSCs into the tail veins of mice with brain ischemia, the infarct volume was significantly reduced (15.8%) in the NDSC-injected group compared to control, and the CM-DiI-positive cells were found in the damaged hippocampus (Fig. 7). Furthermore, we also observed NDSC differentiation in the same regions of the hippocampus suggestive of a therapeutic value for NDSCs in brain injury. The number of NDSCs was increased with increasing age. The NDSCs also express EpiSC and germ line-related genes and can differentiate into hepatocytes, both in vivo and in vitro (unpublished data).
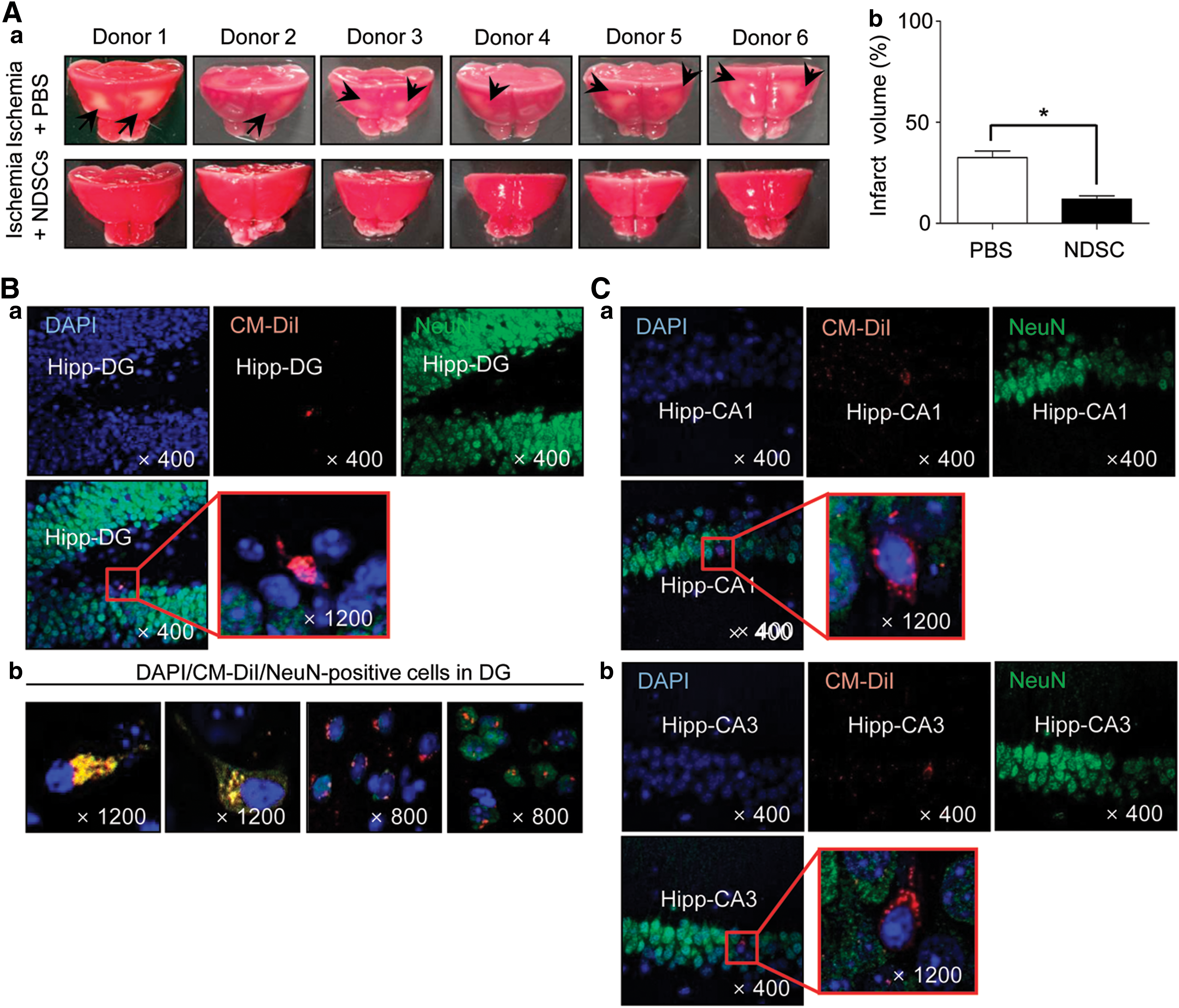
NDSC transplantation in a mouse ischemic brain injury model.
Although, the HAR-NDS forms a network throughout the body and is 100-fold more enriched with NDSCs than with VSELs in the BM, we suspect that the VSELs are NDSCs residing in the HAR-NDS of those organs and they perform their biological functions mainly within the HAR-NDS. Thus, this may be the system responsible for transporting VSEL-like adult stem cells to their target tissues [27].
Limitations and Leverage
There have been some reviews focusing on the concept of BHS or PVS. However, most of the studies have presented a brief summary of Kim's and Soh's findings as well as recent discoveries have related to the role of BHS/PVS in cancer metastasis or acupuncture [24,83 –85]. To the best of our knowledge, this review is the first to describe the discovery, characterization, and HAR-NDS. Moreover, for the first time, we have provided evidence for the discovery of adult stem cell from HAR-NDS and its role in tissue regeneration. Our studies may deal with the anatomical structure that Kim observed, which needs confirmation. We, however, have focused on the role of the system in immune network, hematopoiesis, adult stem cells, and pathways for cells and subcellular structures.
Although many of the Kim's findings have been confirmed, there are several points that cannot be accepted or needs to be reconfirmed. In this section, we will discuss the limitations of previous studies that need to be solved to make it easy to be accepted in the classic biology society.
Although the structural constituents of BHS have been confirmed by several researchers, to date only the existence of the IV (in large caliber blood and lymphatic vessels), OS, and N-BHS has been confirmed (as discussed in the section “the PVS”) and the existence of the other two classes (EV and IO-BHS) still needs to be verified. Furthermore, modern histology cannot accept independent channels (terminal BHD) coming out of the nucleus and connecting cell nuclei to each other. In fact, very complex systems exist within cells that provide communication with neighboring cells; these involve hormonal and nerve signaling, cell junctions (anchoring junctions, communicating junction, and occluding junctions), as well as nuclear pore complexes for the entry and exit of large molecules such as RNA and proteins.
Kim argued that the superficial BHDs have a muscular layer and various cells inside, and their structure is different from that of the deep BHDs. This description is quite similar to that of glands. Moreover, the presence of extracellular DNA and the idea that the DNA of one sanal is equivalent to one chromosome as well as the behavior of sanals such as floating and interacting to generate and degenerate cell-like structures in response to biological conditions at the subcellular level are quite questionable in the light of modern science. The production of new cells from sanals and their mode of division (extracellular and intracellular Bonghan cell cycle) are not acceptable. In the multicellular organism, only two distinct type of cell divisions are found, that is, mitosis and meiosis. Although these findings seem to be interesting, they are likely to be only fantasy resulting from the absence of detail methodology.
Despite explosion of staining methods, there is still an urgent need to develop a better, simple, and more specific imaging technique for HAR-NDS. Classical staining methods, involving alcian blue and trypan blue dye, are simple to use, but are limited in terms of specificity and in vivo imaging ability to elucidate the cytology and anatomical structure of the HAR-NDS. Moreover, modern molecular imaging methods, involving light and electron microscopy, CLSM and X-ray microscopy, require long processing time and/or sophisticated instruments. Development of antibody- or peptide-based fluorescence or radiological imaging for HAR-NDS will facilitate more rapid growth of this field.
Infact, the formation of the BHS/HAR-NDS at an early stage of development, expression of stem cell marker on cells, fluid of this system, and the regenerative properties of NDSC suggest that it is a potential niche of stem cells. Since NDSCs do not represent any ethical problems like ESC or a possible genetic problem like iPSC or an immunorejection problem, they may contribute to the regeneration of cells and tissues. Furthermore, NDSCs seem to be more potential in terms of number of cells, colony forming ability, and survival (till fifth passage) than VSEL. However, the limited knowledge regarding the molecular and functional characteristics of NDSCs needs to be explored more. To rigorously prove the existence of PSCs inside HAR-NDS, reliable subculture of colony must be performed. In addition, the potential of NDSCs to repair damaged tissues, as well as its migration in response to various chemoattractants necessitated further investigation. Furthermore, the authentic, reproducible, robust, and scalable sampling technique to get enough number of cells must be developed.
Thus, there is still a long way to go to check if it could be employed in the clinical advancement for treatments, and, of course, human study is more urgent before getting into clinical care. Therefore, adipose tissue or skin seems to be of interest for HAR-NDS identification in humans.
Conclusions
The HAR-NDS is a new integrated anatomical functional circulatory system, separate from the blood and lymph systems. Since HAR-NDS contains abundant hyaluronic acid, which is a potent regulator of stem cell proliferation and differentiation, it is thought to serve as a store of stem cells that can participate in the regeneration of cells and tissues. In this study, we also describe evidence for the existence of adult PSCs, such as HSCs and NDSCs. Since NDSCs can differentiate into cell lineages of three germ layers such as neurons, hepatocytes and blood cells, and have better colony forming efficiencies than VSELs, they may have an important role in tissue regeneration. Therefore, during pathological conditions, this system is envisioned to circulate the stem cells and other substances, throughout the body, to aid in the recovery of damaged organs. Further studies are required to confirm NDSCs as promising pluripotent adult stem cells and the HAR-NDS as an important conduit for the movement of stem cells to regions of damaged tissue.
Footnotes
Acknowledgments
This work was supported by grants from the Korean Research Foundation (KRF-2005-084-E00001), International Collaborative Project (KIAT-N0000901), Korea Drug Development Fund (KDDF 201408-11) and the National Cancer Center, Korea (NCC-1310430-10).
Author Disclosure Statement
No competing financial interests exist.