
Abstract
Increasing evidence suggests that multipotent stem cells are harbored within a vascular niche inside various organs. Although a precise phenotype of resident vascular stem cells (VSCs) that can function as multipotent stem cells remains unclear, accumulating evidence shows that multipotent VSCs are likely vascular pericytes (PCs) that localize within blood vessels. These PCs are multipotent, possessing the ability to differentiate into various cell types, including vascular lineage cells. In addition, brain PCs are unique: They are derived from neural crest and can differentiate into neural lineage cells. Because PCs in the central nervous system (CNS) can contribute to both neurogenesis and vasculogenesis, they may mediate the reparative process of neurovascular units that are constructed by neural and vascular cells. Here, we describe the activity of PCs when viewed as multipotent VSCs, primarily regarding their neurogenic and vasculogenic potential in the CNS. We also discuss similarities between PCs and other candidates for multipotent VSCs, including perivascular mesenchymal stem cells, neural crest-derived stem cells, adventitial progenitor cells, and adipose-derived stem cells.
Introduction
A
Although the precise phenotypes of multipotent VSCs remain unclear, vascular pericytes (PCs) are a strong candidate of VSCs [2]. In support of this view, we have recently demonstrated that brain vascular PCs acquire multipotent stem cell activity in response to ischemic injury and that these PCs generate neural lineages as well as vascular lineages [6,7]. Therefore, reactive PCs are likely multipotent VSCs, and in response to brain injury, they may aid in reconstructing the neurovascular unit (NVU) that comprises neural and vascular lineage cells.
In this article, we will refer to PCs as a possible source of multipotent VSCs, particularly focusing on their neurogenic and vasculogenic potential in the central nervous system (CNS). We will also compare the characteristics of PCs with those of other types of proposed multipotent VSCs, including perivascular mesenchymal stem cells (MSCs), neural crest-derived stem cells (NCSCs), adventitial progenitor cells (APCs), and adipose-derived stem cells (ADSCs).
What Is the Role of PCs, Particularly in the CNS?
Perivascular cells adjacent to the endothelium of capillaries were originally named as “Rouget” cells by the French scientist Charles-Marie Benjamin Rouget in 1873. However, because of their unique localization at perivascular regions, Rouget cells were renamed as PCs by Zimmerman in 1923. Recently, both PCs and vascular smooth muscle cells have been deemed as mural cells [8,9]. PCs extend long cell processes across the surface of endothelial cells (ECs) [10], a characteristic similar to that exhibited by telocytes [11]. Furthermore, PCs comprise a heterogeneous cell population that can be identified using various markers, such as platelet-derived growth factor receptor beta (PDGFRβ), neuronal/glial 2 (NG2), nestin, alpha smooth muscle actin (αSMA), CD146, and regulator of G protein signaling 5 (RGS5) [6,12 –18]. However, no marker can identify PCs exclusively.
Numerous studies have shown that both PCs and ECs exhibit synergistic effects [8,9,19,20]. For example, EC-secreted platelet-derived growth factor B (PDGFB) binds to PDGFRβ, which eventually activates multiple signals involved in cell proliferation, differentiation, and migration of PCs [21]. PCs construct the blood–brain barrier (BBB)/NVU along with neural [astrocytes (ACs) and neurons] and vascular cells (ECs). PCs regulate cerebral blood flow (CBF) and permeability of this NVU [22,23]. However, a loss of PCs causes the breakdown of this unit [24,25]. Thus, PCs are considered as playing an important role in maintaining the NVU [26]. In addition to their crucial role in maintaining the NVU, increasing evidence suggests that brain PCs exhibit similar activity as multipotent stem cells [6,7,12]. Therefore, in addition to providing a stem cell niche, brain PCs play diverse roles such as regulating CBF and maintaining homeostasis of the BBB/NVU.
Do Vascular PCs Function as Multipotent VSCs Throughout Development?
Vascular PCs surrounding blood vessels reside in multiple organs. These PCs are multipotent stem cells with multilineage differentiation potential, including adipocytes, chondrocytes, osteoblasts, neural cells, and vascular cells [6,12,13,16 –18,27 –30]. Although these studies show that PCs possess the characteristics of multipotent stem cells both in vivo and in vitro, it remains unclear whether they can retain their stemness throughout development. Indeed, in adult mice, we have shown that under normal conditions, brain PCs lack stemness both in vivo and in vitro [6]. Other groups describe that brain PCs can be isolated from adult mouse [31] and rat [12] even under normal conditions and that such “naïve” PCs exhibit the potential of stem cells in vitro [12]. However, it is possible for them to acquire stemness after altering their phenotype during in vitro treatment (eg, specific chemical treatment, repeated passages) because increasing evidence, including our own, indicates that transformation is essential for PCs to develop the stemness and/or to differentiate into non-PC lineage cells, such as neural cells [7,17].
Although we propose that adult-stage brain PCs lack the activity of multipotent VSCs under normal conditions, embryonic brain PCs express high levels of the stem cell marker, nestin, suggesting that they may have stemness even under normal conditions. It remains unclear whether nestin is expressed specifically in neural cells, such as neural stem cells (NSCs) [32], or in vascular lineage cells, such as ECs [33,34] and vascular PCs [12,13]. However, nestin is expressed in multipotent PCs, which can differentiate to both neural and vascular cells [7]. In a previous study using transgenic nestin-green fluorescence protein (GFP) mice, GFP was highly expressed in the brain's embryonic stages but the expression weakened during the postnatal stage and disappeared during adulthood, except in specific regions [35]. Nestin-positive (nestin+) cells decrease during development in a parallel pattern as they lose their multipotency [36]. These findings suggest that PCs gradually lose stemness during development. Thus, we should consider the possibility that stemness of brain PCs differs within the time course of development (Fig. 1A).
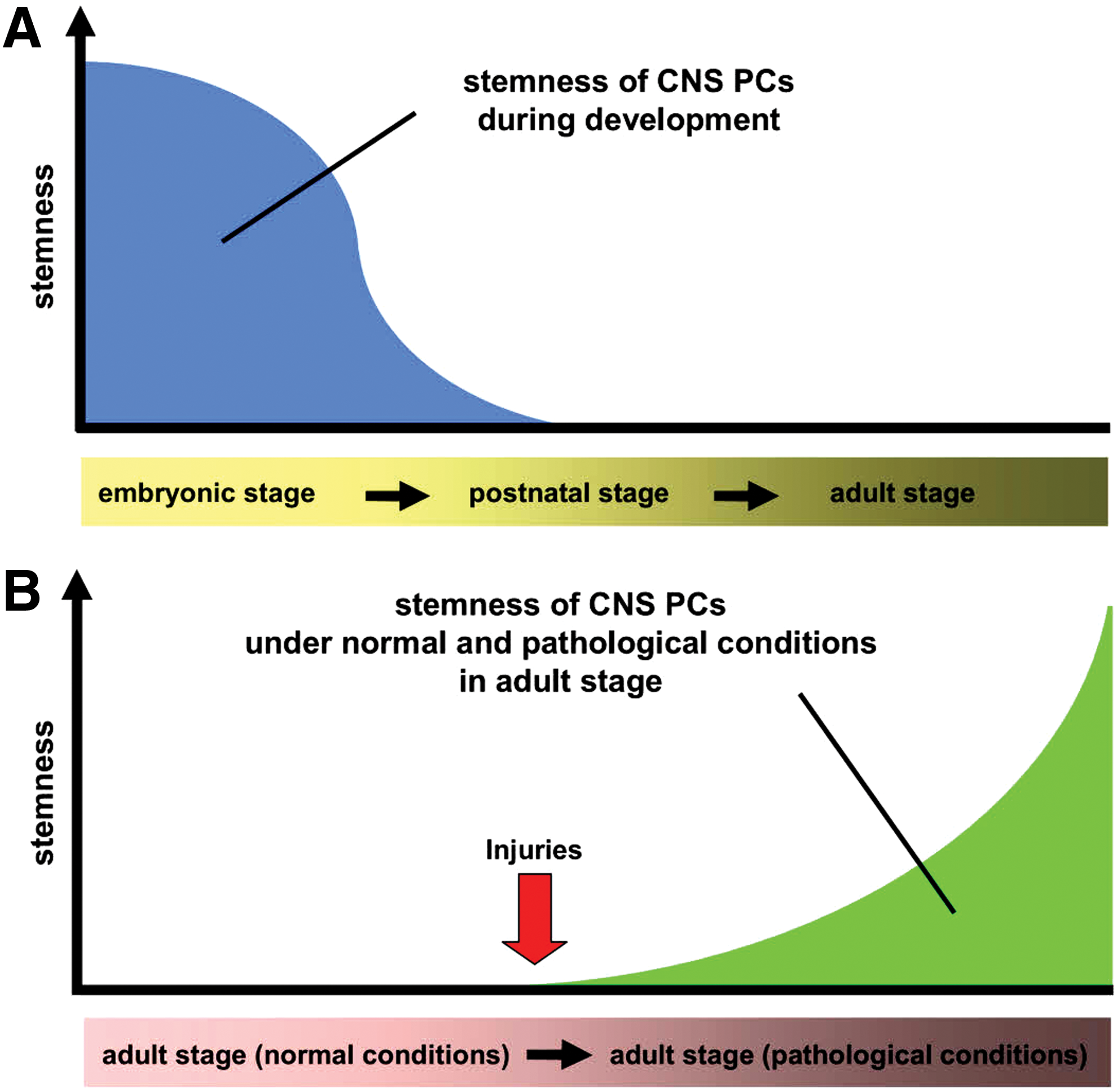
The proposed fate of CNS PCs during development and pathological conditions. It is likely that stemness of CNS PCs is gradually decreased during development
When Do Vascular PCs Change into Multipotent VSCs in the Adult CNS?
PCs are quiescent and slow cycling during their resting stage. However, on stimulation by events, such as hypoxia, PCs become reactive and rapidly proliferate, migrate, and differentiate into various cells [6,18,37] after activation by various mediators, including growth factors (eg, PDGFB, vascular endothelial growth factors, transforming growth factors), transcription factors (eg, hypoxia-inducible factors), and stemness-related factors [38,39]. In addition, although the stem cell marker nestin is not normally expressed postnatally [35,36], it is expressed in brain PCs after ischemia (iPCs) in adult mice, and these nestin+ PCs can be easily isolated from pathological brains [6,40]. This suggests that PCs alter their traits under pathological conditions. Furthermore, using iPCs extracted from the mouse brains and PCs from human brain cultured under oxygen/glucose deprivation (OGD), we have shown that PCs can be reprogrammed into multipotent stem cells in response to ischemia/hypoxia. These iPCs express various stem/undifferentiated cell markers, including nestin, c-myc, Klf4, and Sox2 both in vivo and in vitro, whereas these markers are rarely observed in normal PCs [7]. These findings show that the properties of PCs highly differ between normal and pathological conditions and that reactive PCs (iPCs) are more likely to be multipotent VSCs than are quiescent PCs in the adult CNS (Fig. 1B).
Why does ischemia promote the stemness of PC? Ischemia in vivo is a complex event that cannot be perfectly replicated by OGD alone. However, OGD promotes PC stemness [7], indicating that ischemia/hypoxia reliably induces iPCs. We could not obtain iPCs from nonischemic areas, whereas iPCs were abundant in ischemic areas [6,7,18,41,42]. Furthermore, we found that environmental factors (eg, EC-derived trophic factors) around PCs in addition to OGD stimuli also influenced the fate of iPCs while developing the stemness [7]. Thus, multiple factors, including OGD, induce iPCs after ischemic stroke, although further studies are needed to clarify the critical factors and signaling pathways involved.
Are Perivascular MSCs Vascular PCs?
Similar to PCs, MSCs can be isolated from various organs, including the bone marrow. The mesoderm-derived multipotent stem cells MSCs [43] (Fig. 2) can differentiate into cells such as adipocytes, chondrocytes, and osteoblasts [44]. In addition, certain MSCs reside in a perivascular niche and share similar characteristics with PCs [45 –48], exhibiting activity of multipotent stem cells and expressing markers of both MSCs (eg, CD90, CD105, and CD13) and PCs (eg, PDGFRβ, NG2, αSMA, and RGS5).
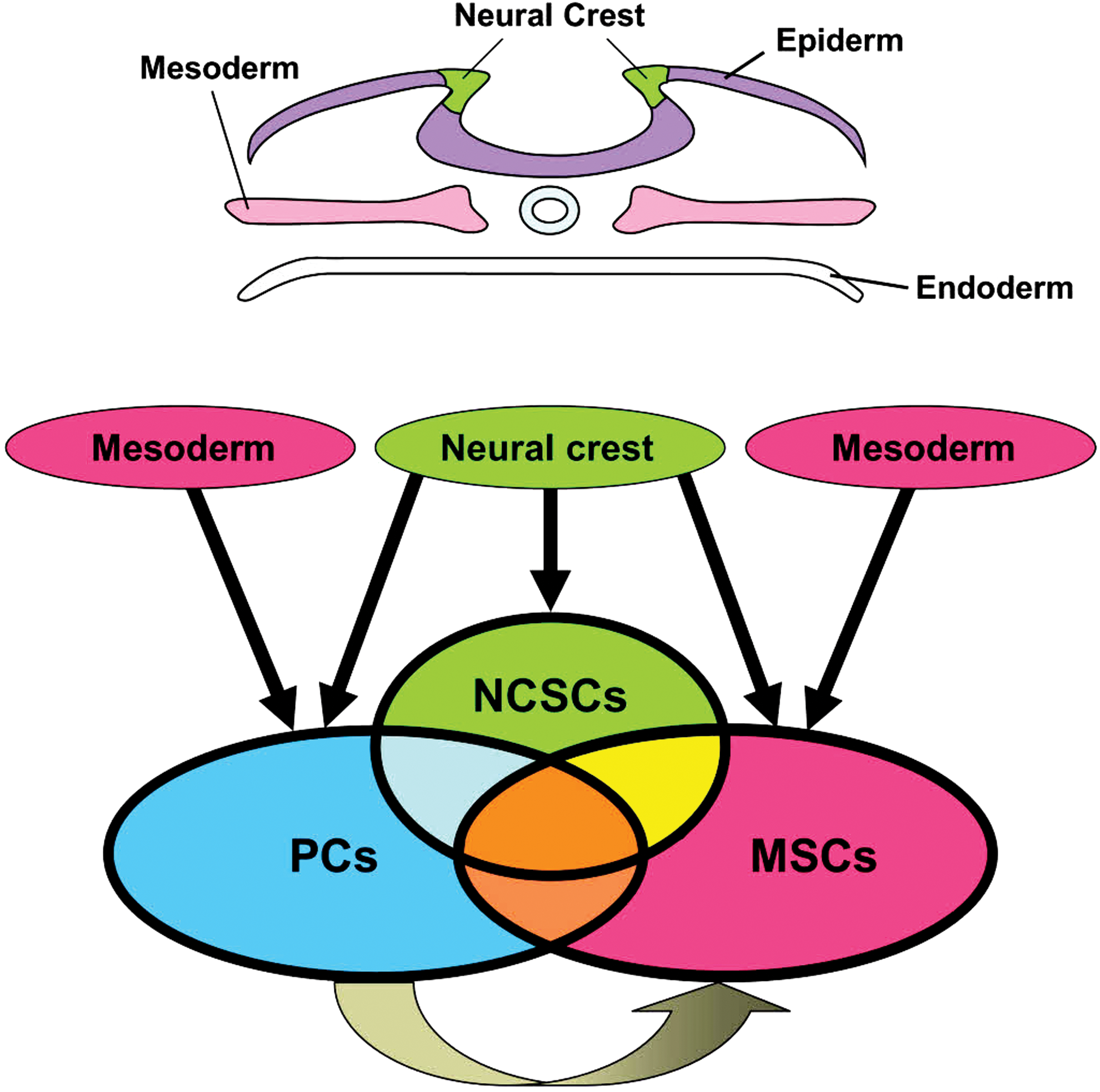
The proposed lineage and similarity among PCs, NCSCs, and MSCs. PCs or MSCs may be derived from the mesoderm and/or neural crest and PCs may produce MSCs. PCs, NCSCs, and MSCs also have overlapping markers and similar phenotypes. MSCs, mesenchymal stem cells; NCSCs, neural crest-derived stem cells. Color images available online at
Although the precise phenotype of perivascular MSCs remains unclear, Kuroda et al. show that subtypes of MSCs contain multilineage-differentiating stress-enduring cells (muse cells) [49,50]. Muse cells are multipotent stem cells of mesenchymal lineage expressing stage-specific embryonic antigen-3 (SSEA-3) and CD105. Muse cells can be isolated from various tissues (eg, bone marrow, skin, and testes). CD105 (endoglin) is known as a marker of both MSCs and muse cells [49,50]. However, PCs with multipotentcy also express CD105 [7,29]. These results suggest that CD105 may be a useful marker of multipotent stem cells, including PCs, MSCs, and muse cells, although CD105 is expressed in nonstem cell populations, including ECs [51]. Furthermore, CD146 (melanoma cell adhesion molecule) is also expressed in MSCs [52,53] and PCs [13,29]. The relationship between perivascular MSCs and PCs remains unknown. However, evidence that they have similar traits and overlapping markers [45 –47] supports the hypothesis that PCs are ancestors of MSCs [54 –56] (Fig. 2).
Why Do Vascular PCs Have Neurogenic Potential?
In general, PCs in organs outside the CNS are less likely to differentiate into neural lineage cells, although they can differentiate into adipocytes, chondroblasts, and osteoblasts [27,28]. However, brain PCs can differentiate into both neural and non-neural lineage cells [6,7,12,17,18], consistent with the traits of perivascular MSCs localized in the brain [45,48]. Brain PCs originate from both the mesoderm [57] and neural crest [57,58] (Fig. 2). The neural crest is developmentally identified as a group of cells found between the neural tube and surface ectoderm in the vertebrate embryo. Similar to PCs, NCSCs have stemness [59] and can differentiate into both neurons and glia [60 –64]. Thus, the neural differentiation potential of brain PCs may be related to their neural crest origin. Furthermore, NCSCs and MSCs have a similar property [65] and the neural crest is considered a potential origin of MSCs [66] as well as that of PCs [57,58] (Fig. 2). The precise relationship between PCs, MSCs, and NCSCs remains unclear. However, if PCs and MSCs are actually derived from the neural crest, it is possible for PCs, MSCs, and NCSCs to have similar traits and overlapping markers.
Neurogenesis occurs near a vascular niche during developmental [67] and pathological conditions [42]. Multipotent PCs express markers of NSCs nestin [6,12,13,18], and nestin+ PCs directly differentiate into neuronal cells [13]. However, the mechanism by which neuronal cells are derived from nestin+ PCs remains unclear. Our recent research showed that nestin+ iPCs simultaneously express mesenchymal markers, such as αSMA, Sox9, Twist, Snail, and Slug, although these markers are gradually downregulated after incubation in the medium promoting NSC induction. In contrast to a decrease in mesenchymal markers, we found that neuroepithelial marker Sox2 was upregulated [7]. These findings indicate that brain PCs differentiate into neural lineage cells in a mesenchymal–epithelial transition-like manner.
We have also demonstrated that Sox2 trasduction in iPCs promotes the expression of several neuronal-related genes, such as MAP2, Neurogenin2, and Mash1, whereas small interfering RNA knockdown of Sox2 results in the inhibition of neuronal-related genes [7]. These findings support previous studies showing that Sox2 induces differentiation of neuronal lineage cells from PCs [17], neural crest cells [68], fibroblasts [69], and hematopoietic cells [70] and underscore the importance of Sox2 as an essential factor for brain PCs to develop traits consistent with neuronal lineage cells. In addition, the mesodermal and neural tubes are derived from common bipotential axial stem cells and Sox2 directs axial stem cells to develop into a neural lineage [71]. Thus, we speculate that multipotent PCs may differentiate predominantly into mesodermal lineage cells, such as adipocytes, chondroblasts, and osteoblasts, unless the Sox2 gene is sufficiently activated and causes them to differentiate into neural lineage cells.
Sox2 is frequently observed in PCs near ECs within ischemic areas after stroke [7], suggesting that a vascular niche, such as that of ECs, may secrete trophic factors after CNS injury. In support of this notion, we found that EC-derived trophic factors, such as leukemia inhibitory factor and fibroblast growth factor-basic [72,73], promoted Sox2 expression in cultured PCs under OGD and initiated their differentiation into neural lineage cells [7]. Thus, under pathological conditions, ECs may facilitate neurogenesis from multipotent PC (iPCs) through signaling, which affects the expression of Sox2 [41,42], although further evidence is necessary to support this hypothesis.
Why Do Vascular PCs Have Vasculogenic Potential?
Certain PCs exhibit vasculogenic traits [16,29] and immature PCs share angioblastic traits during an early stage of vasculogenesis [74,75]. Although under normal conditions, adult brain PCs lack angiogenic properties, after ischemia, iPCs acquire multipotency that reveals a complex phenotype in addition to their original mesenchymal properties, including angioblast expressing hematopoietic stem cell markers such as CD34 [7]. Similarly, reactive MSCs exhibit vasculogenic potential [76,77] not found in MSCs under normal conditions [78,79]. iPCs revealed enhanced expression of a pluripotent marker such as Klf4 [7], which can promote reprogramming into an angioblastic lineage cell [80]. Thus, ischemia/hypoxia may convert normal PCs into multipotent PCs (iPCs) of a mesenchymoangioblast (multipotential mesodermal precursors with angiogenic potential) nature that are typically observed during developmental stages [43]. Notably, a mesenchymoangioblast is considered the origin of MSCs [43,81]. This also supports the hypothesis that MSCs may be derived from certain PCs [54 –56] (Fig. 2). Furthermore, consistent with a phenotype of VSCs [5,82], iPCs coexpress αSMA and CD144 and differentiate into both smooth muscle and ECs that form a tube-like network [7], supporting the vasculogenic potential of iPCs. In addition, the evidence that iPCs have multipotency [7] supports the hypothesis that PCs are likely multipotent VSCs [2]. However, various other types of multipotent stem cells, such as NCSCs, APCs, and ADSCs, may also be multipotent VSCs.
NCSCs have both neural and mesenchymal phenotypes [64], expressing various mesenchymal markers, such as Snail, Slug, twist, and Sox9 [60]. Fate mapping analysis with the neural crest marker myelin protein zero (Po) showed that Po+ cells are present in resident vascular progenitors with endothelial differentiation potential [62], indicating that NCSCs also possess angioblastic properties. Furthermore, NCSCs function consistently with other multipotent PCs and differentiate into multilineage cells, including chondrocytes, osteoblasts, and smooth muscle as well as neural and vascular lineage cells [60 –64]. However, considering that the neural crest is an origin of PCs [57,58], it is reasonable to speculate that NCSCs and PCs have similar properties [6,7] (Fig. 2).
Akin to PCs around capillaries, APCs around larger vessels are also a strong candidate of VSCs [2]. APCs are Sca1+ multipotent stem/progenitor cells localized to the adventitia of blood vessels [83]. Similar to the properties of multipotent PCs [7], APCs are positive for nestin, PDGFRβ, NG2, and CD34 [84 –86] and contribute to vasculogenesis [84,86,87]. In addition, APCs have multilineage differentiation potential for adipocytes, osteocytes, chondrocytes, myocytes, and neural cells [84,87,88], suggesting that APCs have traits similar to those of multipotent PCs. However, the precise origin of APCs remains unclear. Hu et al. found no evidence suggesting that APCs are derived from bone marrow cells [85]. On the other hand, using fate mapping analysis with a neural crest marker, Wnt1, Passman et al. reported that Wnt1+ cells were not observed in APCs and thus they suspected that APCs may derive from the mesoderm [83]. Likewise, APCs are positive for CD44 and CD90 [87,89]. However, these markers are also expressed in MSCs [90]. Thus, MSCs are considered to be derived from APCs [56] other than PCs [54 –56] and NCSCs [66].
Adipose-derived stem cells (ADSCs) express various PC markers [91,92]. Although MSCs generally lack CD34, ADSCs express CD34 [91,93,94] and display endothelial differentiation potential [95], which is observed not in normal but in reactive PCs (iPCs) [7]. The exact origin of ADSCs in adipose tissue is unclear, but several reports show that ADSCs are present in a perivascular niche [96] in the adventitia of large vessels and in capillaries where they coexist with PCs [97]. Because ADSCs are multipotent and can differentiate into various lineage cells, including neural cells, ECs, adipocytes, chondroblasts, and osteoblasts [95,98 –101], they may be subtypes of multipotent PCs and/or VSCs [2,91,92].
The precise source, lineage, subtypes, and traits of multipotent VSCs with vasculogenic properties should be further investigated. However, evidence that PCs, MSCs, NCSCs, APCs, and ADSCs are of a mesenchymoangioblast nature strongly suggests that stem/progenitor cells in primitive stages share the traits of both the mesenchyme and angioblast. The traits of possible multipotent VSC sources, including PCs, MSCs (perivascular MSCs), NCSCs, APCs, and ADSCs, are summarized (Table 1).
ADSCs, adipose-derived stem cells; APCs, adventitial progenitor cells; MSCs, mesenchymal stem cells; NCSCs, neural crest-derived stem cells; PCs, pericytes.
Vascular PCs as a Possible Source of the NVU in the Damaged CNS
Brain vascular PCs are a perivascular element along with neural and vascular cells, constituting the BBB/NVU. Brain injury, such as a stroke, disrupts the BBB/NVU [102], which forms in the CNS as a minimal functional unit [26]. Thus, multipotent stem cells with neurovasculogenic potential can be targeted to repair the injured CNS.
Neurogenesis and vasculogenesis occur coordinately in the CNS during developmental [67] and pathological conditions [42]. Whether the same type of stem cells contributes to both neurogenesis and vasculogenesis remains controversial. However, increasing evidence shows that certain NSCs [103 –105], particularly those with a glioblastoma-like nature, differentiate into not only neural but also vascular lineage cells [106,107]. Thus, under pathological conditions such as ischemic stroke, NSCs/glial progenitors may be strong candidates mediating reconstruction of the NVU (Fig. 3).

The proposed lineage among PCs, NSCs, and glial lineage cells. Various types of stem/progenitor cells in the CNS, such as brain PCs, NSCs, and glial progenitors, have the potential to differentiate into both neural and vascular lineage cells, which can construct the NVU. The proposed relationship between these cells, based on previous reports, is also shown. NSCs, neural stem cells; NVU, neurovascular unit. Color images available online at
The PCs generally do not express glial markers, such as GFAP, aquaporin4, and Olig2 [6]. However, PCs and certain glial progenitors, such as oligodendrocyte precursor cells (OPCs), have common markers such as NG2 [6,12,108,109] and PDGFRα [30,110]. In addition, PCs produce glial lineage cells [6,108,111,112], whereas glia, in turn, generate PCs [113] (Fig. 3). However, MSCs are derived from glia [114] other than from PCs [54 –56]. The relationship between PCs and glia remains unclear, although the findings mentioned earlier suggest that PCs and glial lineage cells may be reciprocal in nature (Fig. 3).
It is well documented that NSCs reside in a perivascular niche [115] and can produce both neuronal and glial cells [116]. However, various types of glial lineage cells, such as radial glial cells [117], subventricular zone ACs [118], reactive ACs [119], resident glial cells [120,121], OPCs [122], and microglial cells [123,124], have been suggested as potential sources of NSCs (Fig. 3). Although the precise origin, identity, and subtypes of NSCs remain unclear, using a mouse model of ischemic stroke, we have demonstrated that iPCs are an origin of NSCs after CNS injury [6], consistent with a previous report showing that PCs are an origin of NSCs [12] (Fig. 3). The iPCs express the NSC marker nestin as well as PC markers PDGFRβ and NG2. Consistent with the phenotype of multipotent PCs [17,29], iPCs exhibit multipotency and differentiate into neural and vascular lineages in vitro [7]. Similarly, others show that nestin+/NG2+ PCs differentiate into both neural and vascular cells [13,16], suggesting that not only NSCs/glial progenitors but also multipotent PCs (iPCs) have the potential to reconstruct the NVU [7] (Fig. 3).
Whether iPCs can differentiate into neurovascular cells to reconstruct this unit in vivo is unclear, because there are multiple factors that influence the fate of iPCs [6,7,41,42,125,126]. We previously showed that although they predominantly differentiated into neuronal cells in vitro [6,41,127], nestin+ iPCs can differentiate into glial lineage cells rather than neuronal lineage in vivo. Consistent with this, fate mapping with the PC subpopulation marker Glast showed that Glast+ spinal cord forms a glial scar after injury [111]. Although the mechanism by which CNS PCs differentiate into glial lineages in vivo remains unclear, various chemical mediators and cytokines are produced at CNS injury sites. Among these, interleukin-6, ciliary neurotrophic factor, and bone morphogenic proteins promote differentiation into a glial lineage [128,129].
Because ECs [41,130] and ACs [131,132] can function as a niche for promoting neurogenesis via cell–cell contact or by activating signaling pathways, these cells also potentially affect the fate of reactive PCs after injury. In addition, PCs are localized at perivascular (Virchow-Robin) spaces in the CNS where immune cells such as macrophages and lymphocytes infiltrate [133,134]. Because PCs can regulate the fate of these inflammatory cells via the immune system [135,136], immune system regulation may be another key factor promoting CNS regeneration after injury. Our previous study showed that suppression of a subpopulation of lymphocytes promotes the survival of iPCs, thereby promoting neurogenesis in vivo [125,126].
Although it is likely that regulation of these factors, including chemical mediators, cytokines, niche cells, cell–cell contact, signaling pathways, and inflammatory cells, is needed for accomplishing iPC-derived neurogenesis in vivo after CNS injury, nestin+/NG2+ multipotent PCs have angiogenic potential in vivo [16]. Furthermore, increasing evidence suggests that exogenous cotransplantation of neural and vascular cells after CNS injury promotes CNS repair [41] by reconstructing the NVU [137]. Future experiments of labeling lineages of PCs through PDGFRβ, NG2, and αSMA will investigate such reparative capacity in vivo. We believe that by understanding the factors that control the fate of iPCs, CNS regeneration via reconstruction of the NVU may soon become a reality.
Concluding Remarks
In conclusion, we discussed the traits of PCs when viewed as multipotent VSCs, with particular focus on their neurogenic and vasculogenic potential. We also compared the traits of PCs with those of previously reported multipotent stem cells. Because several of these cells, including perivascular MSCs, NCSC, APCs, and ADSCs, display properties similar to those of PCs, we are possibly merely examining subsets of the same cells during different stages. In addition, the phenotype of multipotent stem cells may differ with age, organ, species, or stimulus. Thus, a further examination of their properties is required to elucidate appropriate clinical applications.
Footnotes
Acknowledgments
This work was partially supported by a Grant in Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science, and Technology (24650173) and by a Grant in Aid for researchers, Hyogo College of Medicine (2013).
Author Disclosure Statement
No competing financial interests exist.