
Abstract
Pluripotency is at the crossroads of stem cell research and biology of reproduction. The mature metaphase II oocyte contains the key factors for pluripotency induction and maintenance as assessed by its capacity to reprogram somatic nuclei. The cumulus cells (CCs) niche that surrounds the oocyte is crucial for its maturation and presumably for the oocyte to acquire its competence to confer pluripotency. In this study, we examined whether cells cultured from the human mature metaphase II oocyte CC niche (hCC) could be used as feeders for the propagation of human induced pluripotent stem cells. The induced pluripotent (iPS) cells cultured on hCC (hCC-iPS) were assessed for their pluripotency potential by their expression of pluripotency-associated genes such as Oct4, Nanog, and TRA1-60 and their competence to differentiate into the three germ layers in vitro (embryoid bodies) as well as in vivo (teratoma formation). We show that not only the hCC-iPS cells maintained their pluripotency potential, but they also exhibited much better self-renewal performance in terms of proliferation rate compared to the same cells cultured on human foreskin fibroblast (hFF) feeders (hFF-iPS). A comparative gene expression profile study of hCC and hFF revealed significant differences (P<0.05) in expression of cellular matrix components and an upregulation in hCC of genes known to be important players in cell proliferation such as interleukin 6 gene (IL6).
Introduction
O
To maintain their pluripotency, hiPS cells are typically cultivated on murine embryonic fibroblasts (MEF) as feeder layer cells [8,9]. However, MEFs are not recommended to support hPSC for therapy purpose [10,11]. Thus, various feeder cells have been proposed for maintenance of undifferentiated hPSC cells, including human foreskin fibroblasts (hFFs) [5,12 –14], amniotic epithelial cells [15], amniocytes [16], and mesenchymal stem cells [17]. Despite the development of various culture methods, including feeder-free conditions [18 –21], no standardized system has emerged and all of those abovementioned cells are rarely used as feeder layers for clinical application. Most importantly, the factors produced by the feeder that are crucial for self-renewal and pluripotency maintenance still remain unclear.
Cumulus cells (CCs) are specialized granulosa cells that are in close contact with the oocyte. They communicate with the oocyte through specialized gap junctions allowing transport of metabolic molecules and signal exchange. This intimate association is essential for the oocyte to acquire competence [22,23], a stepwise process that includes meiotic maturation up to the metaphase II stage, and the capacity to undergo normal fertilization and support embryonic development [24].
In the present article, we sought to analyze the capacity of cells derived from the CCs to support propagation and pluripotency of hiPS cells. We hypothesized that cultured human cumulus cells (hCCs) could prove valuable as a support for pluripotent cells growth. In this study, we evaluated their capacity to support pluripotency of hiPS cells, and using transcriptomic approaches we characterized the molecular portrait that may account for their properties as feeder layers in comparison with the classically used hFF.
Materials and Methods
CC isolation and establishment of human primary CC culture (hCC)
Cumulus–oocyte complexes were retrieved after signed written informed consent by the couples. The local Institutional Review Board approved the study. The CCs were mechanically separated from the oocytes using needles. They were then transferred to dish coated with a matrix of type I–III human collagen (supplied with SPE-IV medium) at 200 μg/mL in animal-free serum-free defined medium (SPE-IV/EBM-500; ABCell-Bio). After 2 h of culture, the cells began to adhere to the plate surface to form cytoplasmic extensions. On the first day, these extensions increased in number. From the third day onward, cells occupied larger areas of the plate. At confluence, cells were detached with TrypLE™ Select (3436D; Life Technologies) treatment for 5 min at 37°C. They were then seeded onto culture dishes treated with human collagen I–III (supplied with SPE-IV medium) at 100 μg/mL, for expansion.
Preparation of the feeder cells and culture of hiPS cells
The hiPS cells (M4C7) were obtained by reprogramming the human fibroblast foreskin hFF cell line using lentiviral vectors to express human OCT4/POU5F1, SOX2, NANOG, and LIN28 as described [25]. The hiPS cells were cultured in 35-mm wells on hFF feeder or on hCC feeder. For mitotic inactivation, both feeders were treated with 10 mg/mL of mitomycin C (Sigma-Aldrich) for 2 h at 37°C. They were then washed three times with phosphate-buffered saline (PBS) before plating on type I–III human collagen-coated culture dishes at a density of 3.5×104 cells/cm2. The original hCC medium (SPE-IV) was changed to stem cell medium just before hiPS cells were added. Stem cell medium consisted of 80% knockout Dulbecco's modified Eagle's medium (KO-DMEM; Invitrogen), 20% knockout serum replacement (Invitrogen), 0.1 mM non-essential amino acids, 2 mM
Immunocytochemical analyses
hiPS cells and differentiated cells were fixed for 20 min in 4% paraformaldehyde in PBS and washed three times in PBS. For immunostaining, cells were permeabilized with 0.1% Triton X-100 (Sigma). After blocking at room temperature for 60 min with 5% donkey serum (Chemicon International) in PBS, cells were incubated for 1 h at room temperature with primary antibody diluted in PBS with 5% donkey serum: Calcium channel, voltage-dependent (CAV1.2) (1:100; NeuroMab), Connexin 43 (1:300; Santa Cruz), and Alpha actin (1:500; Sigma). Cells were then washed three times in PBS and incubated for 1 h at room temperature with the secondary antibodies: anti-rabbit fluorescein isothiocyanate (FITC, 1:1,000; Molecular Probes) and anti-mouse Alexa Fluor 568 (1:1,000; Jackson ImmunoResearch). Unbound antibodies were removed by three washes in PBS. Cell nuclei were detected with Hoechst staining (5 μg/mL; Sigma-Aldrich). Alkaline phosphatase (ALP) staining was performed using the Vector Red Alkaline Phosphatase Substrate Kit I (SK-5100; Vector Laboratories) according to the manufacturer's protocol.
Flow cytometry analysis
hiPS cells were dissociated with TrypLE at 37°C for 10 min. Cell surface pluripotency markers were identified using a cocktail of four antibodies against SSEA4-PERCP (R&D), TRA-1-60-FITC (BD), TRA-1-81-APC (BD), and CD24-PB (EXBIO) and after PBS washes, cells were suspended in FACSFlow (Becton Dickinson;
RNA extraction
The RNeasy Micro Kit (ref. 74004; Qiagen) was used to extract total RNA from each cumulus sample and the RNeasy Mini Kit (ref. 74104; Qiagen) was used to extract total RNA from hCC and hiPS samples, respectively, according to the manufacturers' recommended protocols. The quantity and purity of the total RNAs were determined by using a NanoDrop ND-1000 spectrophotometer (NanoDrop ND, Thermo Fisher Scientific) and their integrity by using the Agilent 2100 Bioanalyzer (Agilent Technologies;
Transcriptomic data analysis and visualization
Total RNA (200 ng) was used to prepare cRNA (one cycle of amplification) using the Affymetrix 3′ IVT express protocol as described [26]. After fragmentation, the labeled antisense aRNA (15 μg) was hybridized to HG-U133 Plus 2.0 GeneChip pan-genomic oligonucleotide arrays (Affymetrix). Microarray experiments were performed in the IRMB (Montpellier University Hospital) DNA microarray platform. After image processing with the Affymetrix GeneChip Operating 1.4 software (GCOS), the CEL files were analyzed using the Affymetrix Expression Console Software v1.3.1 and normalized with the MAS5.0 algorithm by scaling each array to a target value (TGT) of 100 using the global scaling method. Gene annotation was performed using NetAffx (
In vitro differentiation
For endoderm induction, a protocol modified from previously published studies on human embryonic stem cells (hESCs) was applied [30]. hiPS colonies were mechanically disaggregated and replated onto a Matrigel-coated plate (40 μg/cm2) in hiPS medium conditioned by hCC for 48 h. When cells reached 50% confluence, the medium was replaced by an endoderm medium for 5 days consisting of DMEM with 5.5 mM glucose, 2 mM
Teratoma formation and histological analysis
NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ (NSG) mice (Charles River) were used for the teratoma induction. Animal experiments were performed in accordance with the European and National guidelines (Council Directive 2010/63/EU) and the institutional and regional ethics committees (permit number CEAA-LR-12157). Briefly, teratoma development was assessed by injecting hiPS cells as described [25]. Approximately 7 weeks after the graft, teratomas were dissected and fixed with 3.7% formaldehyde in PBS. Paraffin-embedded tissue sections were then generated and stained with Hematoxylin–Eosin–Safranin staining.
Reverse transcription–quantitative polymerase chain reaction analysis
Total RNA was isolated from cells using the RNeasy Mini Kit (Qiagen). The RNA extraction was performed according to the manufacturer's instructions. The concentration and quality of isolated RNA were determined using a ND-1000 Spectrophotometer (NanoDrop Technologies). Five hundred nanograms of total RNA was used for reverse transcription in a final volume of 20 μL with the SuperScript® II (Invitrogen) according to the manufacturer's instructions. Aliquots of the RT reaction were diluted in water to a 1/20 final dilution. Quantitative polymerase chain reaction (PCR) was performed using a LightCycler ® 480 Real-Time PCR System (Roche Diagnostics) with the LC480 SYBR Green I Master Kit (Roche Diagnostics) containing 2 μL cDNA and 0.5 μM forward and reverse primers in a total volume of 10 μL. Sequences of primers are given in Supplementary Table S1 (Supplementary Data are available online at
Enzyme linked immunosorbent assay
Culture medium conditioned by hCC for 48 h were stored at −20°C until analyzed. The level of IL6 cytokine (DY206-05) in the culture supernatants was analyzed using commercially available DuoSet ELISA Kits (R&D). The reaction intensity was assessed by measuring the OD450nm in an ELISA plate reader (PR 3100; Bio-Rad).
Statistical analysis
Each experiment was performed as least three times and data are shown as the mean±standard error of the mean. The differences were evaluated using Student's t-tests. The probability of <0.05 was considered to be statistically significant.
Results
Cells derived from the human cumulus as a substrate for expansion of PSCs
In the present work, we were able to maintain hCC in culture until passage 14, after which their growth slowed down. Moreover, hCC were successfully recovered after cryopreservation. These properties are of particular interest for a possible use of hCC as a feeder layer for the expansion and maintenance of PSCs. To address these issues and gain insight into the molecular basis that underlies the differences between hCC and hFF, a comprehensive analysis was performed as outlined in the workflow presented in Fig. 1.
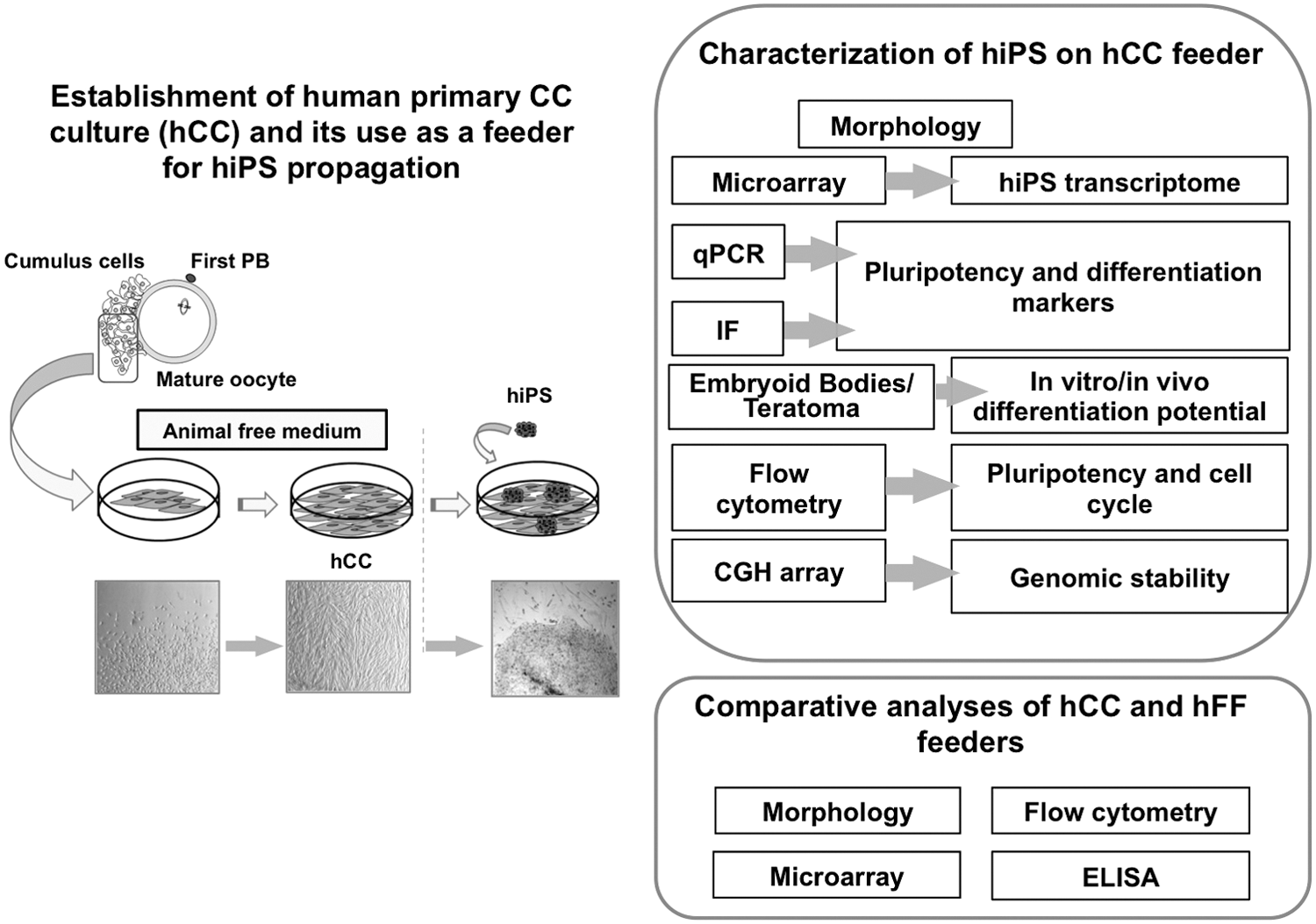
Overview of the workflow.
Morphological and molecular characterization of hCC compared to hFF
Adherent hCC cultures seem to be morphologically similar to fibroblasts (hFF) (Fig. 2A). To better understand to which extent these cells compare, we performed a nonsupervised analysis of their gene expression profile along with an unpassaged cumulus expression profile. The hierarchical clustering approach indicated that hCC and hFF cluster together (Fig. 2B). This is accounted for by the correlation matrix that demonstrated correlation coefficient of 0.851 between hCC and hFF (Supplementary Fig. S1). However, the heat map showed significant differences between hCC and hFF profiles (Fig. 2B). The 0.845 correlation coefficient between hCC and cumulus suggests that hCC have retained cumulus features (Supplementary Fig. S1). The analysis of specific markers using flow cytometry revealed that hCC expressed vimentin mesenchymal marker. Epithelial markers (as expressed in HT-29 cell line), such as EpCam and cytokeratin (KRT18), were either absent (EpCam) or poorly expressed (KRT18) in hCC (Fig. 2C). The transcriptomic analysis revealed that neither of the three markers, vimentin, EpCam, and KRT18 mRNAs were expressed in the unpassaged cumulus (Fig. 2D) underlining that hCC acquired new properties. It is noteworthy that the expression of vimentin was lower in hCC compared to fibroblasts.

Characterization of human cumulus cell (hCC).
hCC feeder layer maintains hiPS cells in an undifferentiated state and promotes their proliferation
We first evaluated the ability of hCC to support growth of hiPS cells in an undifferentiated state. At first sight, the colonies of hiPS cells cultured on hCC feeder (hiPS-hCC) appeared rounded with obvious boundaries with hCC feeder, a morphology that is consistent with an undifferentiated state. Then, we investigated the expression of several pluripotency markers by hiPS-hCC. As shown in Fig. 3A, the hiPS-hCC colonies displayed a strong ALP activity. The expression of OCT3/4 and NANOG pluripotency nuclear markers was assessed by quantitative reverse transcription–polymerase chain reaction analysis (qRT-PCR) experiments and the expression of pluripotency surface markers (CD24, SSEA4, TRA-1-81, and TRA-1-60) was evaluated by flow cytometry analysis. The resulting data demonstrated that hiPS-hCC retained stemness up to at least passage 10 (Fig. 3B). As an additional test of the hiPS-hCC pluripotency, we performed Affymetrix oligonucleotide microarray to analyze the genome-wide gene expression profile. Unsupervised hierarchical clustering of the transcriptomic data of hiPS-hCC, PSCs (hiPSC and hESC), and different tissue samples representative of the three germ layers (cerebellum, heart, and liver) showed that hiPS-hCC were grouped with the PSCs and were clearly distinct from the differentiated cells (Fig. 3C). In addition, using the array CGH analyses, no subchromosomal aberrations were detected, confirming the genomic stability of the hiPS-hCC after seven continuous passages (Fig. 3D). Finally, the only difference that could be observed between hiPS-hCC and hiPS-hFF was the slightly bigger diameters of hiPS-hCC colonies (Fig. 3E) that resulted from an improved growth of the hiPS-hCC cells (Supplementary Fig. S2). To gain further understanding we performed flow cytometry analyses of the cell cycle of hiPS cells grown on the two types of support. These analyses revealed a higher proportion of hiPS cells in S-phase when cultivated on hCC in comparison to the same cells cultivated on hFF feeder (Fig. 3F).
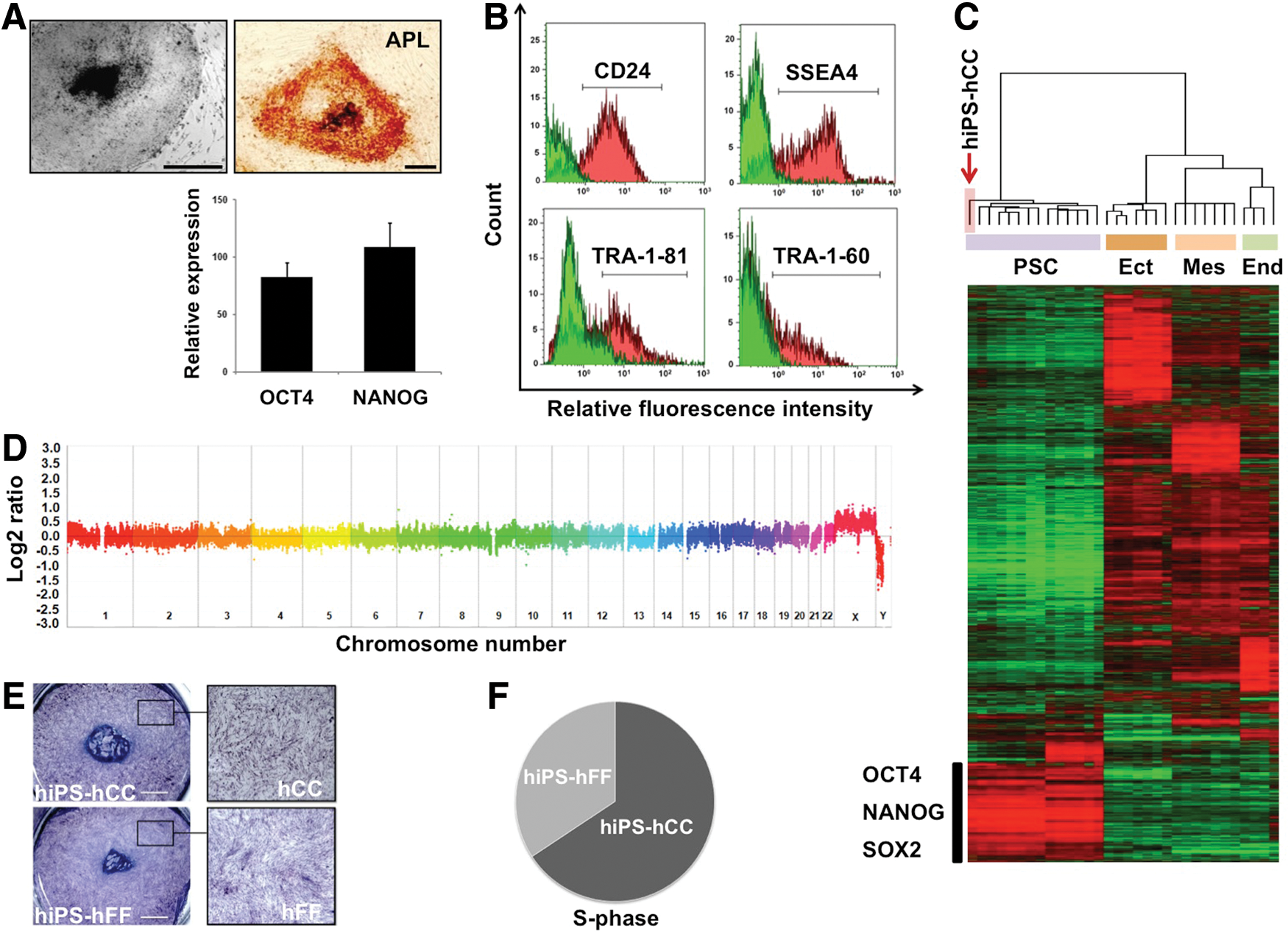
Human induced pluripotent stem (hiPS) cells cultured on hCC maintain pluripotency and genetic stability.
hiPS-hCC maintain their potential for differentiation
To evaluate the pluripotency of hiPS-hCC cells, they were assayed for their capacity to differentiate in vitro into lineages representative of the three embryonic germ layers (ectoderm, endoderm, and mesoderm). Conversion into ectoderm was assayed by the addition of N2B27 and bFGF to the culture medium. After approximately 10 days of culture of hiPS-hCC on matrigel, clusters of neural cells appeared and qRT-PCR analysis demonstrated that these hiPS-hCC-derived cells turned on NESTIN and PAX6 expressions (Fig. 4A). Endoderm differentiation was assayed by cultivating hiPS-hCC in the presence of WNT3A, Activin A, and FGF2. As revealed by the qRT-PCR analysis, HEX, FOXA2, CXCR4, and SOX17 endodermal markers were expressed (Fig. 4A). To assess the capacity of hiPS-hCC to differentiate into mesodermal derivatives, EBs were produced through their cultivation in suspension for 7 days followed by plating them onto collagen-coated dishes. Approximately 10 days after plating, attachment and further spontaneous differentiation, contracting cell aggregates were observed (Supplementary Video S1). qRT-PCR and immunofluorescence analyses of these aggregates revealed the expression of cardiac-specific markers, GATA-4, Nkx2.5, TBX5, CAV1.2, CX43, and α-actinin (Fig. 4A, B), indicating the presence of functional cardiomyocytes. All the data consistently showed that the differentiation markers were strongly induced while the expression of the OCT4 and NANOG pluripotency markers displayed a significant decrease during the differentiation process (Fig. 4A). A teratoma assay was performed to assess the in vivo differentiation capacity of hiPS-hCC. The transplantation of hiPS-hCC into immune-compromised mice resulted in the formation of teratomas, which contained tissues of the three germ layers (Fig. 4C–E).
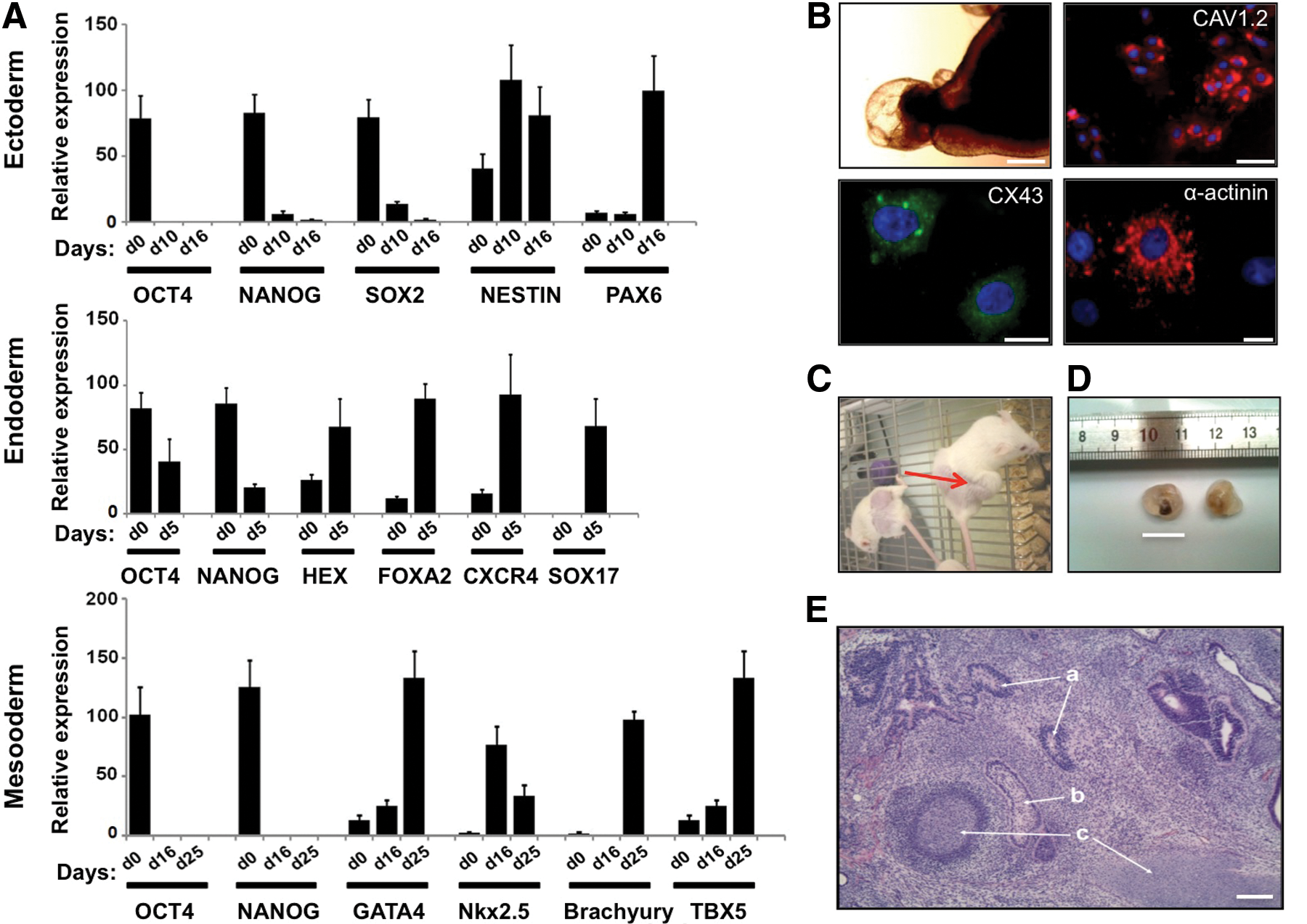
In vitro and in vivo differentiation capacity of hiPS grown on hCC feeder.
Dissecting the molecular divergence between hCC and hFF feeders
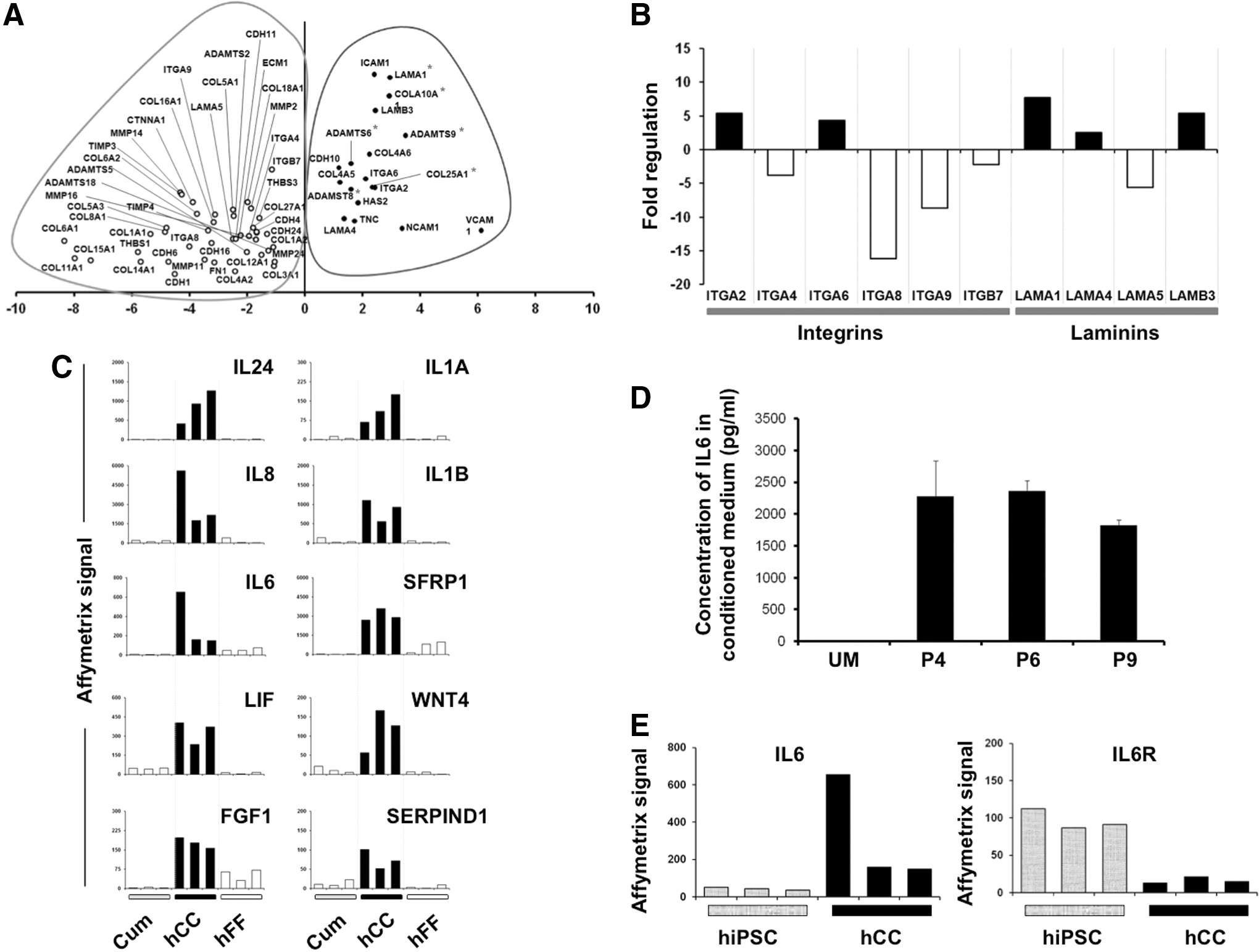
Next, we sought to dissect the molecular differences between the two feeder types. Comparison of the transcriptomic profiles of hCC and hFF using the SAM software with a two-FC cutoff and a FDR <5% showed that 1,860 and 6,748 transcripts were upregulated in hCC and hFF, respectively (Supplementary Tables S2 and S3). Some genes encoding functions that are essential in cell signaling showed higher expression levels in hCC (Supplementary Table S4). The list of the 50 genes whose FC varies between 15<FC<467 with a significant statistical value (FDR <5%) are presented in Supplementary Table S5. To identify a gene expression signature specific to hCC (expressed neither in hFF nor in Cum), we confronted hCC expression profiles to both hFF and cumulus transcriptomes. Using the SAM analysis, we found that 722 transcripts were overexpressed and 1,898 underexpressed in hCC specifically (Fig. 5A). The functional GO annotations of these two lists revealed that there were significantly (P<0.001) more genes involved in cytokine activity (GO:0005125), transmembrane transporter activity (GO:0003677), ion transport (GO:0006811), and membrane localization (GO:0044425) in hCC. Conversely, genes associated with localization in the nucleus (GO:0005634) and connected with transcription factor activity (GO:0003700) and regulation of transcription (GO:0045449) were underrepresented (Fig. 5B).
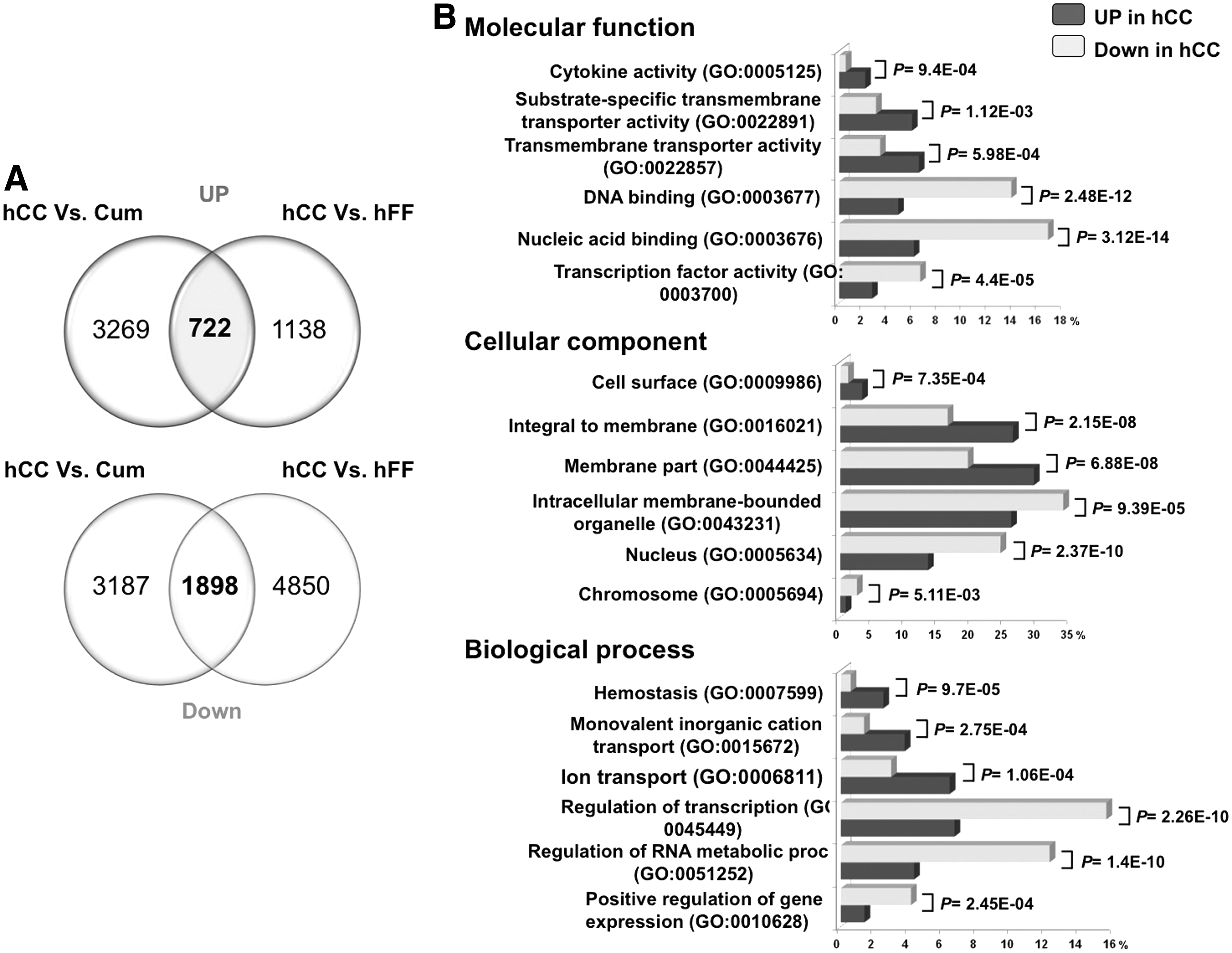
Comparative analysis of hCC versus cumulus (cum) and hFF transcriptomes.
Expression of genes encoding extracellular matrix and cytokines in hCC
Considering that cell-to-cell communication genes are a functional category that might play an essential role in the dialog between the feeder and hiPS cells, we focused on the comparison between hCC and hFF of expression profile of extracellular matrix (ECM) and cytokine-encoding genes. Sixty-one differentially expressed ECM and ECM-related protein-coding genes (FC >2, FDR <5%) were extracted and listed in Supplementary Table S8, of which 18/61 were overexpressed in hCC, whereas 43/61 were underexpressed (Fig. 6A). Interestingly, integrins (ITGA2, ITGA6) and laminins (LAMA1, LAMA4, and LAMB3) were among the overexpressed genes in hCC (Fig. 6B). Among the differentially expressed cytokine-encoding genes, 14/25 [for instance, interleukin 6 (IL6)] were overexpressed in hCC, whereas 11 of them were underexpressed (see Supplementary Table S9). Interestingly, some were also overexpressed in hCC in comparison to unpassaged CCs suggesting that these cells have acquired new signaling properties upon cultivation (Fig. 6C). The role of IL6 in cell proliferation is particularly well documented [32,33]. We, therefore, evaluated the secretion of IL6 by hCC feeder into the culture medium after 48 h of incubation, using enzyme-linked immunosorbent assay (ELISA). The analysis of the conditioned culture medium revealed the presence of IL6 at an average concentration of 2 ng/mL (Fig. 6D). In an effort to link the expression of IL6-secreted factor by hCC, we analyzed the expression of its receptor by hiPS cells. The histograms drawn after our micorarray data revealed the expression of IL6 receptor (IL6R) gene in hiPS cells (Fig. 6E). While on the one hand IL6R gene expression was much lower in hCC than in hiPS, on the other hand IL6 gene expression was higher in hCC, suggesting a paracrine signaling from hCC toward hiPS.
Crosstalk between hiPS and hCC.
Discussion
The capacity of hiPS cells to differentiate into various specialized cell types holds promise for the generation of an unlimited supply of cells for regenerative medicine [34,35]. However, the mechanisms that underlie the stemness status are complex and appear to be the result of intrinsic cell potential, cell/matrix, and cell/cell interactions. In this investigation, we report the use of cells derived from the human cumulus (hCC), the in vivo physical support, and microenvironment of the developing oocyte, as feeder to maintain the undifferentiated form of hiPS cells. The rationale for using hCC as feeder is that, during in vitro fertilization procedure, the coculture of human embryos with autologous cumulus improves embryo quality, blastocyst formation and pregnancy rate [36,37]. In this study, we show that during their propagation on hCC feeder, the hiPS cells maintain the expression of the typical pluripotency markers and the ability to differentiate in vitro into many cell types, including those with contracting myocardial tissue. In addition, hiPS cells cultured on hCC retain their ability to differentiate in vivo as revealed by the teratoma formation. The mRNA expression profiles produced by global gene microarray analyses comparing hiPS-hCC and other PSC revealed an overall high similarity in profiles. Moreover hiPS-hCC retained chromosomal stability. All these results indicate that hCC layers are suitable supports for hiPS expansion. Most importantly, hCC feeder impacts the proliferation of the hiPS-hCC. Indeed the diameters of hiPS undifferentiated colonies were bigger when hiPS were grown on hCC than on hFF. Moreover, the flow cytometry analysis showed that the number of cells entering S-phase in hiPS cells grown on hCC was higher than that of cells grown on hFF. Interestingly, the regulation of the cell cycle has been recognized as a key factor in pluripotency maintenance [38,39] and involves a niche environment [40]. ECM or cytokines and growth factors released by the niche are essential in this process. This suggests that hCC express a myriad of factors that could be involved in both the increase of hiPS cells proliferation rate and the maintenance of hiPS cells' essential properties.
Our transcriptome analysis shows that the genes overexpressed specifically in hCC are annotated as either membrane or extracellular, demonstrating a strong bias toward genes involved in cell to cell communication processes. We focused our analysis on gene expression profile of ECM and cytokines with the postulate that they could be important for the hCC feeder performance. Indeed, various ECM proteins such as laminins, fibronectins, collagens, integrins, and heparan sulfate proteoglycans have been reported as key factors provided by the PSC-supportive feeder cells [41 –44]. In this study, we show that some ECM genes such as genes coding for laminins (LAMA1, LAMA4, and LAMB3) are highly overexpressed in hCC in comparison with hFF. Various laminin recombinant isoforms have been shown to support hESC proliferation in an undifferentiated state [45 –48]. Interestingly in a human blastocyst, the natural source of hPSC, LAMA1, LAMA5, and LAMC1 genes are mainly expressed in the trophectoderm [49,50]. It is noteworthy that LAMA1 is overexpressed both by trophectoderm and hCC. However, the impact of LAMA1 on the physiology of hCC-iPS cells and the embryo inner cell mass remains to be examined.
As stated above, numerous cytokines including IL6, IL1B, IL8, IL24, LIF, and FGF1 are strongly expressed by hCC. In addition to high mRNA expression of IL6 by hCC and IL6 receptor by hiPS cells, ELISA assays revealed a substantially higher level of IL6 in the hCC-conditioned medium. These observations have to be considered in light of the essential role played by IL6 in the propagation of hiPS cells in suspension culture [51,52] and in derivation and self-renewal of mESC [53]. The secretion of IL6 by hCC could explain its property to boost proliferation and sustain self-renewal of hiPS cells. In our attempt to compare trophectoderm and hCC, it is worth noting that IL6 is also expressed in trophectoderm [49], suggesting that IL6 pathway is operative in blastocyst. More investigations are required to identify and study the role of the cytokines secreted by hCC in successful maintenance and higher proliferation of hiPS cells. Our system may be further exploited to identify novel factors that support iPS cells pluripotency and proliferation, which may ultimately help to design a completely defined and animal-free medium that is compatible with therapeutic applications. Before that, some issues must be addressed such as whether the hCC can help to support hiPS cells for long term and if hCC-conditioned media can preserve pluripotency using feeder-free culture methods. Beyond molecular insights into the support of hiPS cells, this new culture system will have other interesting applications in routine in vitro fertilization procedures.
Conclusion
In summary, this study demonstrates that the hCC system is convenient to sustain hiPS cells. There are several advantages for using cumulus-derived cells as feeders: (i) Cumulus can easily be obtained as medical wastes and be used as an abundant source to generate feeder layers. (ii) The process is inexpensive, very rapid, and non-invasive for the oocyte. (iii) Most hiPSC lines are derived from hFF and are therefore XY. Thus, any contamination can be easily assessed when using cells of maternal origin (XX) such as hCC as feeder. Finally, this culture system can be regarded as a biological model for the investigation of oocyte–niche interactions.
Footnotes
Acknowledgments
The authors thank the direction of the University Hospital of Montpellier for their support. The authors thank Sevrine Kremmer for technical support with ELISA, Catherine Panabières, Guilhem Requirand, and Laure Cayrefourcq for excellent technical assistance with cytometry analysis, and Dr. Béatrice Orsetti for technical support with CGH array analysis. They acknowledge Dr. Catherine Panabières for providing HT-29 cells and Dr. Hassan Boukhaddaoui for help with cell imaging (Montpellier RIO Imaging). Last but not the least, the authors would like to warmly thank the ART Department team. Ferring Pharmaceuticals partially contributed to the Affymetrix chips used for the cumulus analysis.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.