
Abstract
The hedgehog (Hh) signaling pathway plays fundamental roles during embryonic development and tumorigenesis. Previously, we have shown that ablation of the tumor suppressor and negative regulator, Suppressor of fused (Sufu), within this pathway causes embryonic lethality around E9.5 in the mouse. In this study, we examine how lack of Sufu influences early cell fate determination processes. We established embryonic stem cell (ESC) lines from preimplantation Sufu−/− and wild-type mouse embryos and show that these ESCs express the typical pluripotency markers, alkaline phosphatase, SSEA-1, Oct4, Sox2, and Nanog. We demonstrate that these ESCs express all core Hh pathway components and that glioma-associated protein (Gli)1 mRNA levels are increased in Sufu−/− ESCs. Upon spontaneous differentiation of Sufu−/− ESCs into embryoid bodies (EBs) in vitro, the Hh pathway is strongly upregulated as indicated by an increase in both Gli1 and patched1 (Ptch1) gene expression. Interestingly, developing Sufu−/− EBs were smaller than their wild-type counterparts and showed decreased expression of the ectodermal markers, Fgf5 and Sox1. In vivo teratoma formation revealed that Sufu−/− ESCs have a limited capacity for differentiation as the resulting tumors lacked the mesodermal derivatives, cartilage and bone. However, Sufu−/− ESCs were able to develop into chondrocytes and osteocytes in vitro, which suggests a differential response of ESCs compared with in vivo conditions. Our findings suggest a regulatory function of the Hh signaling pathway in early mesodermal cell fate determination and emphasize the role of Sufu as a key molecule in this process.
Introduction
T
The Hh ligands, Sonic hedgehog (Shh), Desert hedgehog (Dhh), and Indian hedgehog (Ihh), bind to and inhibit the receptor, Patched1 (Ptch1), an interaction that results in the activation of another transmembrane protein known as Smoothened (Smo). Activation of Smo causes dissociation of a complex formed between Suppressor of fused (Sufu) and the glioma-associated protein (Gli, Gli1–3) transcription factors, resulting in the translocation of activated Gli into the nucleus and the initiation of target gene transcription. In the absence of an Hh signal, Sufu interacts with Gli and regulates Gli protein activity.
Hh target genes include regulators of proliferation and differentiation, as well as genes for cell death and stem cell renewal. In addition, both Ptch1 and Gli1 are themselves Hh-activated target genes, likely playing a feedback role to control the level of Hh transcriptional activation. Thus, the expression levels of Ptch1 and Gli1 are often used to monitor Hh signaling activity.
Embryonic stem cells (ESCs) represent an important tool in studies of early mammalian development and differentiation. ESCs are self-renewing and pluripotent, meaning that they can differentiate into all cell types in the body [4]. Results on the role of Hh signaling in ESCs are partly contradictory. Undifferentiated human (huESCs) and mouse ESCs (mESCs) were shown to possess primary cilia, express Hh signaling pathway components, and display endogenous signaling activation [5 –8]. Although long-term treatment of undifferentiated mESCs with Shh stimulated proliferation and increased Gli1 mRNA expression [9], addition of Shh to huESCs did not affect pluripotency or proliferation [8].
Sufu is a negative regulator in the Hh pathway and Sufu-null mice exhibit an embryonic lethal phenotype displaying neural tube defects [10,11]. A recent study utilizing chondrocyte-specific knockout mice demonstrated that Sufu is highly expressed during the early stages of chondrocyte differentiation and has an essential function in endochondral bone formation, regulating chondrocyte proliferation and differentiation [12].
To study the effects of a loss of Sufu on early differentiation and lineage specification processes, we established ESCs from preimplantation Sufu−/− embryos. In the current study, we show that Sufu−/− ESCs can be maintained in an undifferentiated state, form embryoid bodies (EBs) in vitro and teratomas in vivo, and can be differentiated directly into chondrocytes and osteocytes in vitro. Furthermore, upon spontaneous differentiation, EBs formed from Sufu−/− ESCs displayed a strong increase in Ptch1 and Gli1 expression. Additionally, we observed that Sufu−/− EBs appeared smaller in size compared with their wild-type counterparts and displayed reduced ectodermal differentiation as suggested by reverse transcription quantitative real-time PCR (RT-qPCR) marker analysis.
Both wild-type and Sufu−/− ESCs were able to form teratomas in vivo, but strikingly, the ablation of Sufu resulted in the complete absence of the mesodermal derivatives, cartilage and bone. These results demonstrate that Sufu plays an important role in regulating mesodermal differentiation in vivo.
Materials and Methods
Mouse strains and genotyping
The Sufu+/− mice (strain B6.129X1/SvJ-Sufutm1Rto ) in this study were between the 14th (N14) and 17th (N17) generation of backcrossing to C57BL/6J when they were intercrossed to obtain preimplantation embryos. Genotyping of mice and ESCs was performed as described previously (11). BALB/c nude spontaneous mutant mice [strain C.Cg/AnBomTac-Foxn1nu N20 (Charles River)] were used for the teratoma experiments. The mice were housed in a specific pathogen-free barrier facility according to local and national regulations, and the study was approved by the Stockholm South Animal Ethics Committee.
ESC derivation and culturing
For the derivation of ESCs, two previously published protocols were combined and modified [13,14]. Time-mated Sufu+/− females were sacrificed at embryonic day 3.5 (E3.5) by cervical dislocation. Uteri were dissected and blastocysts were flushed from the uterine horns using Dulbecco's modified Eagle's medium (DMEM) containing 10 mM HEPES buffer solution (Invitrogen) under a dissection microscope.
Blastocysts were collected and transferred individually to wells containing a mitotically inactivated monolayer of mouse embryonic fibroblasts (MEFs) in KSRES medium [KnockOut-DMEM (K-DMEM), 20% KnockOut serum replacement, 2 mM
Blastocysts were allowed to attach to the MEFs and hatch spontaneously. After 5–6 days, the inner cell masses (ICMs) were picked under a microscope and trypsinized by incubation in TrypLE Express (Invitrogen) for 5 min at 37°C. The ICMs were dissociated mechanically and transferred to wells containing mitotically inactivated MEFs and KSRES medium + PD98059. Several days later, typical ESC colonies could be detected in ∼40% of the wells. ESCs were fed daily and passaged approximately every second to third day. After the second passage, the inhibitor, PD98059, was omitted from the growth media. Mycoplasma testing was performed on all cell lines using the MycoAlert® Mycoplasma Detection Kit (Lonza Rockland).
MEFs were isolated essentially as described [15] and were mitotically inactivated, either by exposure to 30 Gy γ-irradiation before seeding or by treatment with 10 μg/mL mitomycin-C (Sigma-Aldrich) for 2–6 h after seeding.
ESC pluripotency was determined using the Alkaline Phosphatase Detection Kit (Merck Millipore) according to the manufacturer's instructions. Karyotype analysis of the ESC lines was performed essentially as described previously [15]. Chromosome counts were determined from at least 20 metaphase spreads of each cell line, and sex determination was performed using polymerase chain reaction (PCR) on ESC DNA [16].
Immunocytochemistry
For immunocytochemistry, ESCs were grown on irradiated MEFs seeded onto Lab-Tek™ Permanox™ Chamber Slides™ (ThermoScientific), followed by culturing for 2–3 days until the ESC clones reached 70% confluency. ESCs were fixed in 4% paraformaldehyde (PFA) in phosphate-buffered saline (PBS) for 15 min and washed twice with 1× PBS. After permeabilization with 0.1% Triton X-100 for 10 min, 4% serum [goat or rabbit serum (Sigma-Aldrich), depending on the secondary antibody] in PBS was applied to the cells for 30 min. Thereafter, cells were incubated at 4°C overnight with the following primary antibodies diluted to the indicated concentrations in 4% serum/PBS: anti-SSEA-1 (1:50; Abcam); anti-Oct4 (1:200; Santa Cruz); anti-Sox2 (1:500; Abcam); and anti-Nanog (1:200; Abcam).
After washing the cells with PBS, the following Alexa Fluor 488-conjugated (Life Technologies) secondary antibodies, diluted 1:400 before use, were applied: anti-mouse IgM (SSEA1), anti-mouse IgG2b (Oct3/4), and anti-rabbit IgG (Sox2, Nanog), all from Invitrogen, and the cells were incubated for 1 h at room temperature.
For nuclear staining, cells were incubated for 3 min at 37°C with 5 μM DRAQ5 (Alexis Biochemicals), followed by mounting with ProLong Gold antifade reagent (Invitrogen). The slides were examined using a Zeiss LSM510 confocal microscope with lasers set at 488 nm for Alexa Fluor 488 and at 633 nm for DRAQ5.
EB formation
EBs were produced using the hanging drop method [17] in KSRES medium without leukemia inhibitory factor (LIF). Essentially, cultures were enriched for ESCs as described for the RT-qPCR analysis, then 500 ESCs in 30 μL drops of medium were placed on the lid of a Petri dish, inverted, and cultured at 37°C in a cell culture incubator. EBs were picked after 2 days (d2) in hanging drops and were thereafter maintained separately in suspension culture in 96-well, ultra-low attachment culture dishes (Corning) until they were again collected for analyses after 3 (d2 + 3) and 12 (d2 + 12) days in suspension.
For size measurements and quantitation of ATP present, the littermate ESC lines, STE671-8B and STE671-8D, derived from the female STE671-8 were compared as were the littermate pair of ESC lines, STE670-3G and STE670-3D, derived from the female STE670-3.
Statistical analysis was performed using Student's t-test. Relative ATP levels were assessed using the CellTiter-Glo® Luminescent Cell Viability Assay (Promega) following the manufacturer's instructions.
For histology, EBs were fixed in 4% PFA in PBS for 1–2 h, followed by dehydration in 70% ethanol and embedding in paraffin. The paraffin blocks were then cut into 4-μm serial sections and characterized using hematoxylin–eosin (H&E) staining for general morphology. To quantify the number of phospho-histone H3-positive cells, two serial sections per EB were analyzed and values were averaged. At least seven EBs per genotype and time point were evaluated.
RNA isolation and RT-qPCR analysis
ESCs, growing exponentially on a feeder layer of irradiated MEFs, were harvested from the culture dish using the recombinant enzyme, TrypLE Express (Invitrogen). Subsequent enrichment for ESCs was performed for 15–20 min at 37°C on a 0.1% gelatin (Sigma-Aldrich)-coated Petri dish, which allowed the MEFs to attach. Following attachment, the ESCs were washed in 1× PBS (Invitrogen). For each cell line and each time point (d2 + 3, d2 + 12), 96 EBs were collected and washed in 1× PBS (Invitrogen) and thereafter snap-frozen and stored at −80°C.
Total RNA was prepared using either RNeasy spin columns (Qiagen) with on-column DNaseI treatment for 15 min at room temperature or the RNAqueous micro kit (Ambion) with subsequent DNAseI treatment for 20 min at 37°C. The RNA was quantified using an Infinite 200 NanoQuant microplate reader (Tecan), and up to 1 μg RNA was used for cDNA synthesis with SuperScript III and Oligo(dT)12–18 primers (both Invitrogen) according to the manufacturer's instructions.
Real-time PCR reactions were performed using FastStart Universal SYBR Green Master (Roche). Analysis was conducted using a 7500 Fast Real-Time PCR system (Applied Biosystems). Primer pair sequences are listed in Supplementary Table S1 (Supplementary Data are available online at
Teratoma formation
For the teratoma experiments, ESCs, growing exponentially on irradiated MEFs, were harvested from the culture dish using TrypLE Express (Invitrogen) and were enriched as described above for the RT-qPCR. The ESCs were washed in 1× PBS (Invitrogen) and the desired number of cells, in a total volume of 100 μL PBS containing 30% BD Matrigel™ (BD Biosciences), were injected subcutaneously into the flanks of 8-week-old nude mice using a 30-gauge U-100 insulin syringe (BD Medical). Four to five mice for each genotype and cell concentration (5,000; 10,000; 100,000; 500,000; 1 million ESCs) were used.
Teratoma growth was determined by palpation and measured using a caliper on a weekly basis. Palpable teratomas started to develop approximately 2–3 weeks after injection and were collected when they had reached the set maximum size of 1 cm in diameter or, at the latest, after 12 weeks postinjection. All animals were sacrificed by cervical dislocation and tumors, together with the surrounding tissue, were dissected and fixed in 4% PFA in PBS overnight, followed by dehydration in 70% ethanol.
Teratomas were divided into parts and all tissue embedded in paraffin in such a way as to ensure sectioning at various levels and directions. The paraffin blocks were then cut into 4-μm serial sections and characterized using H&E staining for general morphology, periodic acid-Schiff diastase (PAS-D) staining to visualize goblet cells, and Alcian blue solution to visualize acidic polysaccharides found in chondroid matrix and mucous-producing cells.
Immunohistochemistry
The following primary antibodies were used for immunohistochemistry: anti-Sox2 (1:1,000; Abcam); anti-synaptophysin (1:40; Abcam); anti-K5 (1:5,000; Covance); anti-Sma (1:500; Abcam); anti-FoxA2 (1:1,000; Millipore); anti-Sox9 (1:200; R&D); anti-phospho-histone H3 (1:1,000; Cell Signaling); anti-Ki67 (1:500, clone SP6; Thermo Scientific), and anti-caspase 3 (cleaved, Asp175) (1:200; Cell Signaling).
Antigen retrieval was performed either for 30 min in DIVA Decloaker (DV2004MX; Biocare Medical) or in 10 mM Na-citrate buffer (pH 6.0), both using the 2100-Retriever (Proteogenix). Biotinylated anti-rabbit or anti-goat IgG secondary antibodies (1:200; Vector Laboratories), streptavidin-conjugated peroxidase (Invitrogen), and diaminobenzidine (Life Technologies) were used to visualize the bound primary antibodies, followed by hematoxylin counterstaining and mounting in Aqua-Poly/Mount (Polysciences). Mouse primary antibodies were used in combination with the Vector® M.O.M.™ Immunodetection kit (Vector Laboratories).
Chondrocyte and osteocyte differentiation in vitro
ESCs were cultured in ESC medium containing K-DMEM supplemented with 2 mM GlutaMAX-I, 0.1 mM minimum essential medium (MEM) with nonessential amino acids, 55 μM β-mercaptoethanol (all from Life Technologies), 15% ESC-tested fetal bovine serum (FBS; Tissue Culture Biologicals), 50 μg/mL ascorbic acid, and 1,000 U/mL LIF (ESGRO; Millipore) on gelatinized tissue culture plates with mitomycin C (Sigma-Aldrich)-inactivated MEFs as a feeder layer.
The differentiation of ESCs into chondrocytes and osteocytes was performed as described previously [20,21] with modifications. Specifically, ESCs were detached from the feeder layer with 0.25% trypsin–ethylenediaminetetraacetic acid and feeder depleted for 1 h by culturing on a gelatinized plate. EBs were formed as hanging drops in ESC medium without LIF (∼500 cells per EB).
After 2 days as hanging drops, EBs were collected and cultured as a suspension culture in an ultra-low attachment plate in the presence of 100 nM all-trans retinoic acid (ATRA) in ESC medium without LIF to induce cell commitment toward the mesoderm. After 2 days of induction, the EBs were plated on gelatin and cultured in αMEM supplemented with 20% FBS and 1% penicillin/streptomycin for 6 days, changing the medium every other day. On day 6, the outgrown EB-derived cells were replated onto a fresh gelatinized plate in αMEM supplemented with 20% FBS and cultured for another 2 weeks; splitting if necessary.
To promote chondrocyte and osteocyte differentiation of EB-derived cells, the mouse mesenchymal stem cell (MSC) functional identification kit from R&D Systems (Catalog No. SC010) was employed. Chondrogenic micromasses were generated according to the manufacturer's instructions. After 21 days, the cell pellet was fixed in formalin and sectioned. To determine if the cells differentiated into a chondrocyte lineage, we used immunofluorescence staining with antibodies specific to aggrecan (Santa Cruz Biotechnology) and collagen 2a1 (Santa Cruz Biotechnology), proteins present in chondrocyte matrix. The secondary antibodies used for immunofluorescence were a goat anti-rabbit Alexa Fluor 594 and a donkey anti-goat Alexa Fluor 488-conjugated antibody (both Life Technologies).
To promote the differentiation to the osteocyte lineage, EB-derived cells were seeded at a high density (∼80% confluence) on BD BioCoat™ fibronectin-coated culture slides in αMEM supplemented with 20% FBS and 1% penicillin/streptomycin, followed by incubating with osteogenic differentiation medium, prepared according to the manufacturer's instructions. The medium was changed every 3 days. After 6 weeks, differentiated cells were fixed with 4% PFA in PBS. The differentiation was documented by Alizarin red staining (EMD Millipore) and by immunofluorescence staining with an antibody recognizing the osteocyte matrix protein, osteopontin (R&D Systems). The secondary antibody used for immunofluorescence was a donkey anti-goat Alexa Fluor 488-conjugated antibody (Life Technologies).
Results
Sufu−/− ESCs can be isolated from mouse blastocysts
To study the role of Sufu during the proliferation and differentiation of stem cells, we established mESC lines of the desired genotypes (wild-type and Sufu−/− ) from blastocysts of Sufu+/− mice intercrossings [11]. We obtained multiple lines with typical ESC morphology from blastocysts originating from six Sufu+/− female mice and performed Sufu genotyping of the ESC lines using PCR (data not shown). Karyotype analysis of metaphase spreads showed that the frequency of euploid cells in the various ESC lines ranged from 45% to 92%. There were no apparent differences in the growth rate (data not shown) and morphology relating to the Sufu genotype among the lines (Fig. 1A, upper panel).
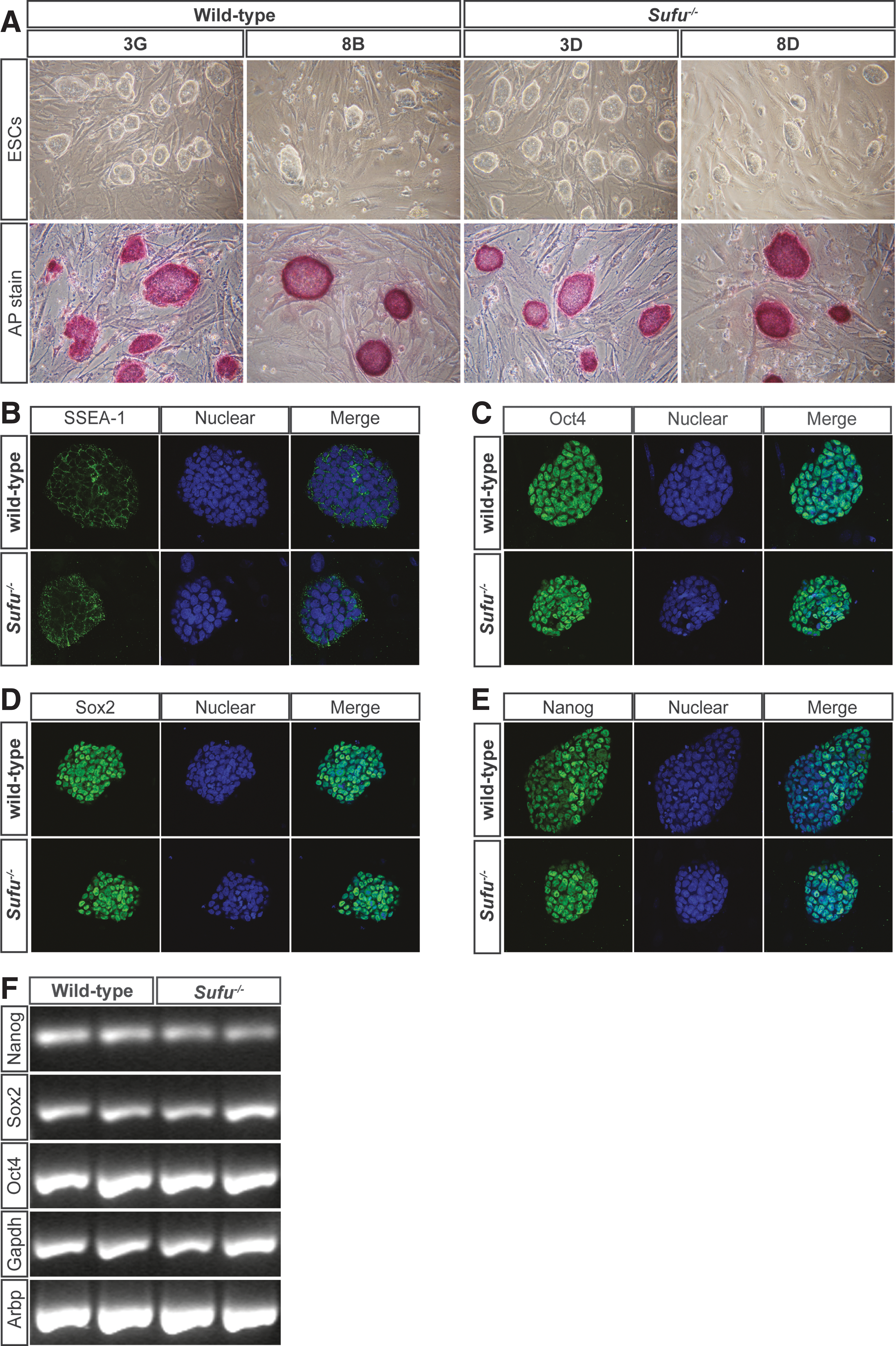
Loss of Suppressor of fused (Sufu) does not affect pluripotency.
We selected the two wild-type [STE670-3G (63%) and STE671-8B (60%)] and Sufu−/− ESC lines [STE670-3D (73%) and STE671-8D (73%)] for further characterization and experiments. For EB and teratoma formation, all experiments were performed using the littermate ESC line pairs, STE670-3G (3G) with STE670-3D (3D) and STE671-8B (8B) with STE671-8D (8D); however, only the results for the representative pair, 3G and 3D, are described if not indicated otherwise.
Sufu−/− ESCs express typical pluripotency markers and can be kept in an undifferentiated state
To assess the pluripotency status of the different ESC lines, we stained for alkaline phosphatase activity, a marker for undifferentiated human and mESCs. As shown in Fig. 1A (lower panel), all ESC lines were alkaline phosphatase positive with a high frequency (>95%) of positive ESC colonies within each line. The pluripotent identity of the derived cell lines was supported by immunofluorescent staining with antibodies against the ESC markers, SSEA-1 (Fig. 1B), Oct4 (Fig. 1C), Sox2 (Fig. 1D), and Nanog (Fig. 1E), which were expressed in a similar manner in both wild-type and Sufu−/− ESCs among the different ESC lines.
Additionally, we confirmed the expression of Nanog, Sox2, and Oct4 by RT-PCR and found no apparent differences in expression between the wild-type and Sufu−/− ESC lines (Fig. 1F). Hence, our results clearly demonstrate that in the absence of Sufu, murine ESCs maintain their pluripotent character and can be kept in an undifferentiated state.
Sufu loss leads to increased Hh pathway activation in undifferentiated mESCs
Human ESCs have been shown to express Hh signaling components and display Hh pathway activation [8]. However, in mESC lines, expression of the core Hh pathway components seems to vary in different studies, probably due to differences in genetic background and the methods of analysis used [7,22,23]. Thus, first we used RT-qPCR to determine whether Hh signaling components were expressed in our ESCs. Both wild-type and Sufu−/− ESCs expressed Shh (Fig. 2A), Ihh (Fig. 2B), Dhh (Fig. 2C), Smo (Fig. 2E), Gli2 (Fig. 2G), and Gli3 (Fig. 2H) at comparable levels.
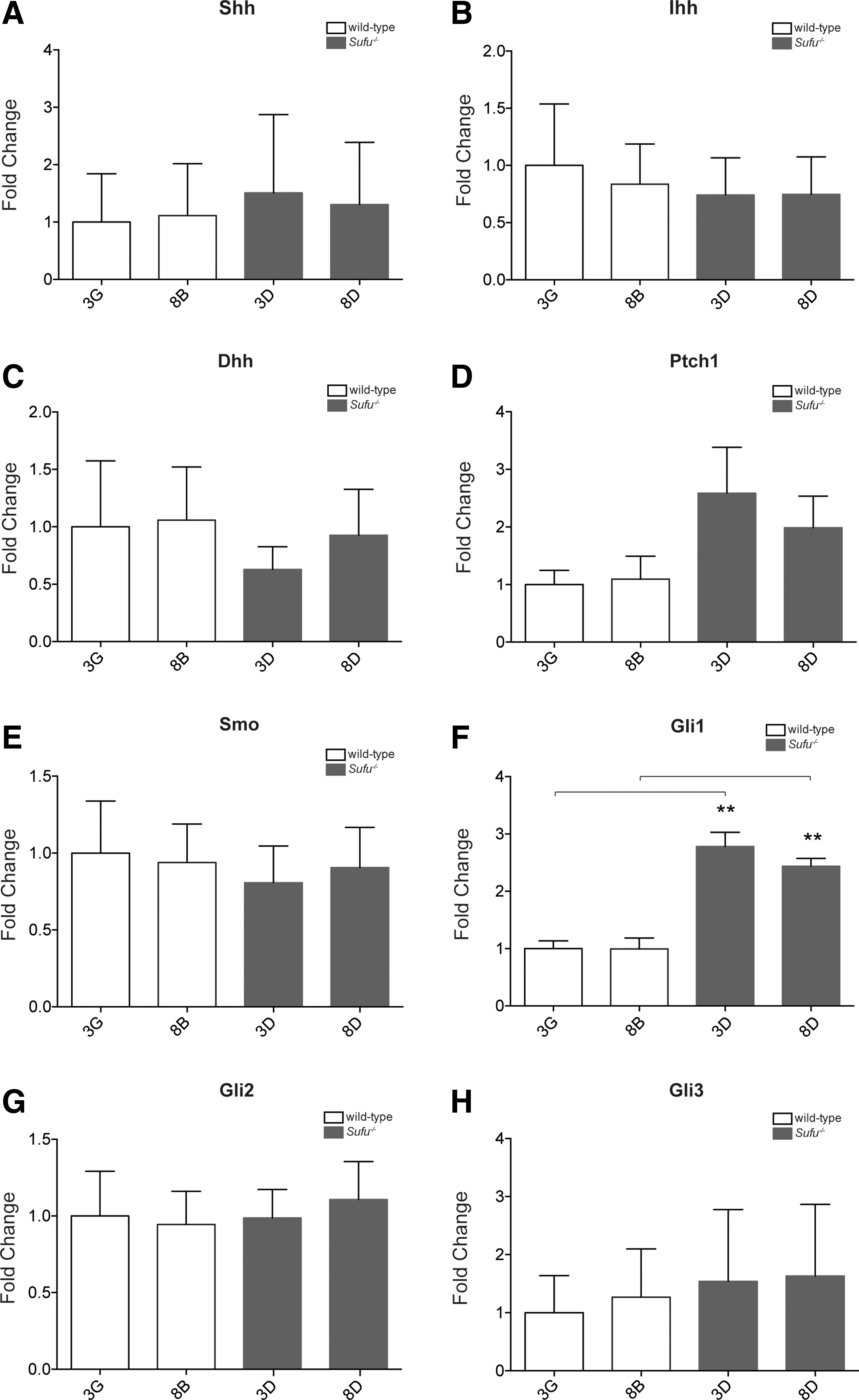
Sufu−/−
ESCs display increased hedgehog (Hh) pathway activity. Expression of Hh pathway genes was analyzed by RT-qPCR on total RNA from wild-type and Sufu−/−
ESCs.
Since Hh signaling increases dramatically in Sufu−/− MEFs [11], we were particularly interested in the effect of the loss of Sufu on Hh pathway activity in our ESCs. The most widely used indicator of Hh pathway activation is the expression of Hh target genes, of which the pathway components, Ptch1 and Gli1, are the best defined. We detected a significant increase in Gli1 expression in Sufu−/− ESCs (Fig. 2F), while Ptch1 expression showed an upward trend (Fig. 2D) compared with wild-type ESCs. However, the increase in Gli1 mRNA levels in Sufu−/− ESCs was modest compared with the dramatic expression observed in Sufu−/− MEFs (Supplementary Fig. S1). Thus, our data suggest that the ablation of Sufu in ESCs does lead to changes in Hh pathway activity, but is not sufficient to reach such high levels as seen in Sufu−/− MEFs.
Sufu loss does not impede EB formation in vitro, but does constrain their size
When grown in suspension without LIF, ESCs spontaneously form EBs, cell aggregates that contain cells of all three germ layers [24]. Diminished Hh signaling in ESCs was shown to lead to a reduction in EB size [23], thus we were interested in whether the absence of Sufu would affect EB formation and growth.
We cultured a defined number of ESCs without LIF in hanging drops and transferred them to 96-well plates after 2 days, where they continued to grow in suspension culture for up to 18 days. After 2 days in hanging drops, the Sufu−/− ESCs had formed aggregates comparable with their wild-type counterparts. After another 3 days in suspension culture (d2 + 3), all EBs appeared small, round, and regularly shaped, and there was no apparent difference between wild-type and Sufu−/− EBs (Fig. 3A). Interestingly, from d2 + 6 and onward, EBs formed from Sufu−/− ESCs appeared smaller in size (Fig. 3A). Measurements at the various time points confirmed a significant reduction in size for the Sufu−/− EBs at later stages (Fig. 3B). To further quantify this difference, we measured the amount of ATP produced per EB using a cell viability assay and found a significant reduction in Sufu−/− EBs at d2 + 12 (Supplementary Fig. S2). To understand if histological disparities could account for the size reduction, we analyzed H&E-stained 4-μm paraffin sections, but could not detect any obvious differences (data not shown). Thus, we performed immunohistochemical staining for the mitotic marker phospho-histone H3 (Fig. 3C, D) as well as cleaved caspase 3 (Fig. 3E, F), a marker identifying apoptotic cells. Counts of phospho-histone H3-positive cells per EB did not reveal significant differences in proliferation between wild-type and Sufu−/− EBs (Fig. 3G). Apoptosis at d2 + 12, however, appeared to be lower in EBs lacking Sufu. We conclude that the absence of Sufu does not prevent EB formation, but does affect the size of later stage EBs.
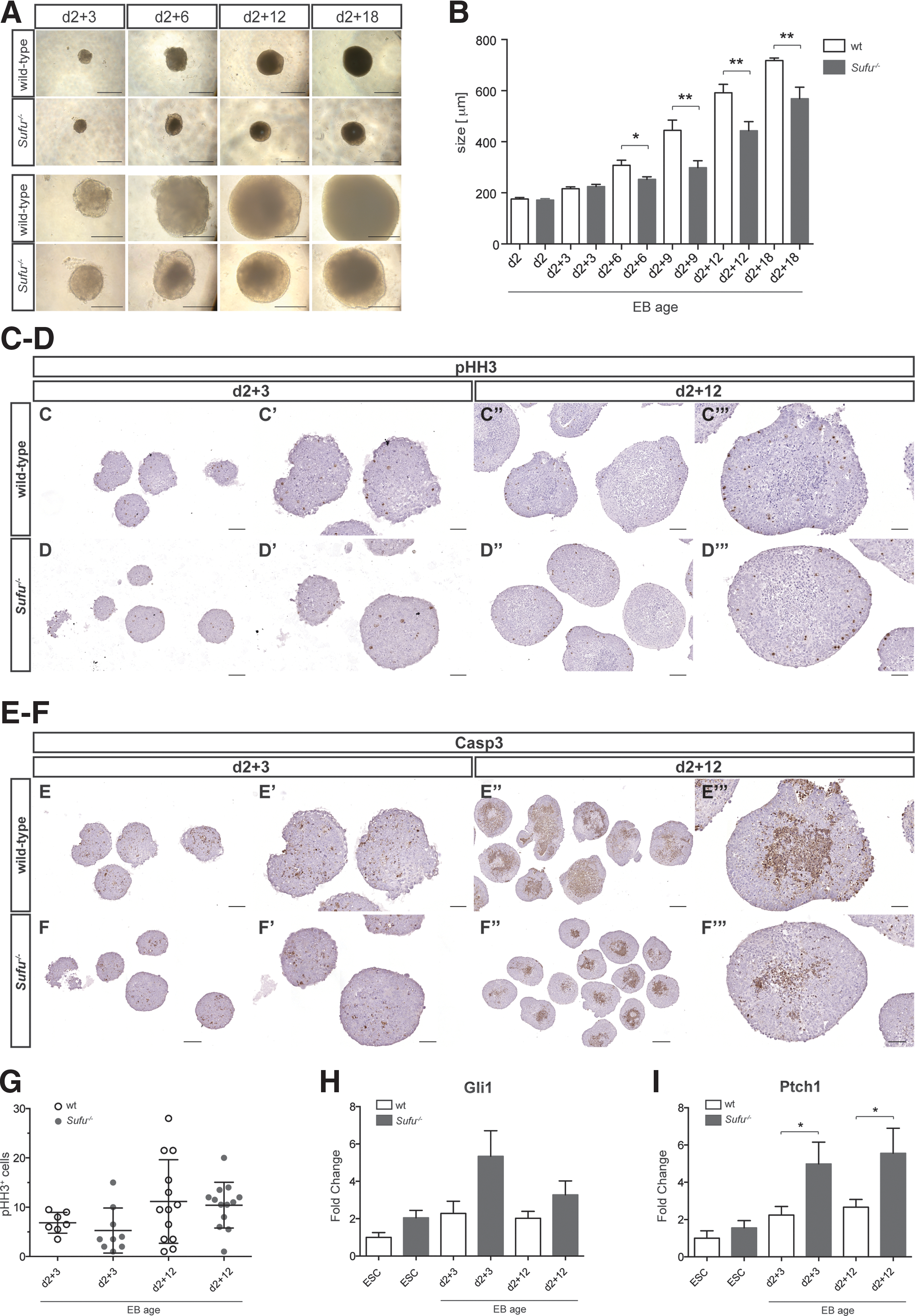
Loss of Sufu affects the growth of late-stage embryoid bodies (EBs).
Sufu ablation stimulates Hh pathway activity during differentiation in vitro and affects lineage specification
Previous studies have shown that Hh signaling is increased in differentiating EBs [23]. Since we could detect small but significant differences in pathway activation in undifferentiated ESCs, we were interested in whether Hh pathway activity would be augmented further upon spontaneous differentiation in vitro.
We analyzed the levels of Ptch1 and Gli1 mRNA expression in undifferentiated ESCs as well as in d2 + 3 and d2 + 12 EBs. Our data demonstrate that Gli1 increased during the early differentiation of wild-type and Sufu−/− ESCs into EBs, then decreased marginally at later EB stages (Fig. 3H). Gli1 expression peaked at d2 + 3 in Sufu−/− EBs and was increased in early and late stages compared with wild-type EBs, although this was not statistically significant. As for Gli1, we detected an increase in Ptch1 levels in EBs of both genotypes during the early differentiation stages (Fig. 3I). Strikingly, Ptch1 expression was significantly higher in both early and late stage Sufu−/− EBs compared with wild-type EBs. Hence, in the absence of Sufu, differentiating cells showed a greater increase in Hh signaling activity compared with wild-type EBs.
Since Hh signaling has been demonstrated to play a role in regulating cell fate [8,23,25 –27], we sought to investigate the consequences of the observed Hh pathway activation on lineage choice. Brachyury and Flk1, both genes critical for mesoderm development [28,29], and Mef2c, a marker involved in cardiomyogenesis [30], did not display significant differences in expression in EBs derived from wild-type and Sufu−/− ESCs (Supplementary Fig. S3A–C).
Sox17 and FoxA2, markers for definitive and visceral endoderm, respectively [31,32], were upregulated during differentiation in both wild-type and Sufu−/− EBs. Whereas Sox17 was slightly lower expressed in late-stage Sufu−/− EBs compared with wild-type EBs, FoxA2 was upregulated (Supplementary Fig. S3D, E). However, neither difference reached statistical significance. Expression of the primitive ectoderm markers, Fgf5 [33] and Sox1, for identification of neuroectodermal development [34], was induced in both wild-type and Sufu−/− EBs already at an early stage (d2 + 3). However, both markers were expressed at a lower level in EBs lacking Sufu compared with wild-type EBs (Supplementary Fig. S3F, G), suggestive of a reduced capacity to differentiate into neuroectoderm.
Loss of Sufu limits mesodermal development in vivo
Teratomas, tumors that contain tissue derivates from all three germ layers, are frequently used to assess the pluripotent capacity of ESCs [35]. To evaluate the developmental potential of our Sufu−/− ESCs in a teratoma assay, ESCs from a range of 5,000 to 1 million cells were injected subcutaneously into the flanks of nude mice. Both wild-type and Sufu−/− ESCs were able to form tumors at comparable efficiencies and latencies. The tumors were solid and appeared to move freely against the covering skin during the growth period of up to 12 weeks (Supplementary Fig. S4A, B).
The initial titration experiments, starting with 1 million cells, produced very rapidly growing tumors independent of genotype and were morphologically very immature (data not shown). Once the numbers of cells were reduced, a less immature picture appeared. No obvious differences in proliferation or apoptosis were detected between Sufu−/− and wild-type tumors as revealed by Ki67 (Fig. 4A–B′), phospho-histone H3 (Fig. 4C–D′), and cleaved caspase 3 (Fig. 4E–F′) staining. Taken together, these results suggest that loss of Sufu does not influence the ability of ESCs to form teratomas in vivo.
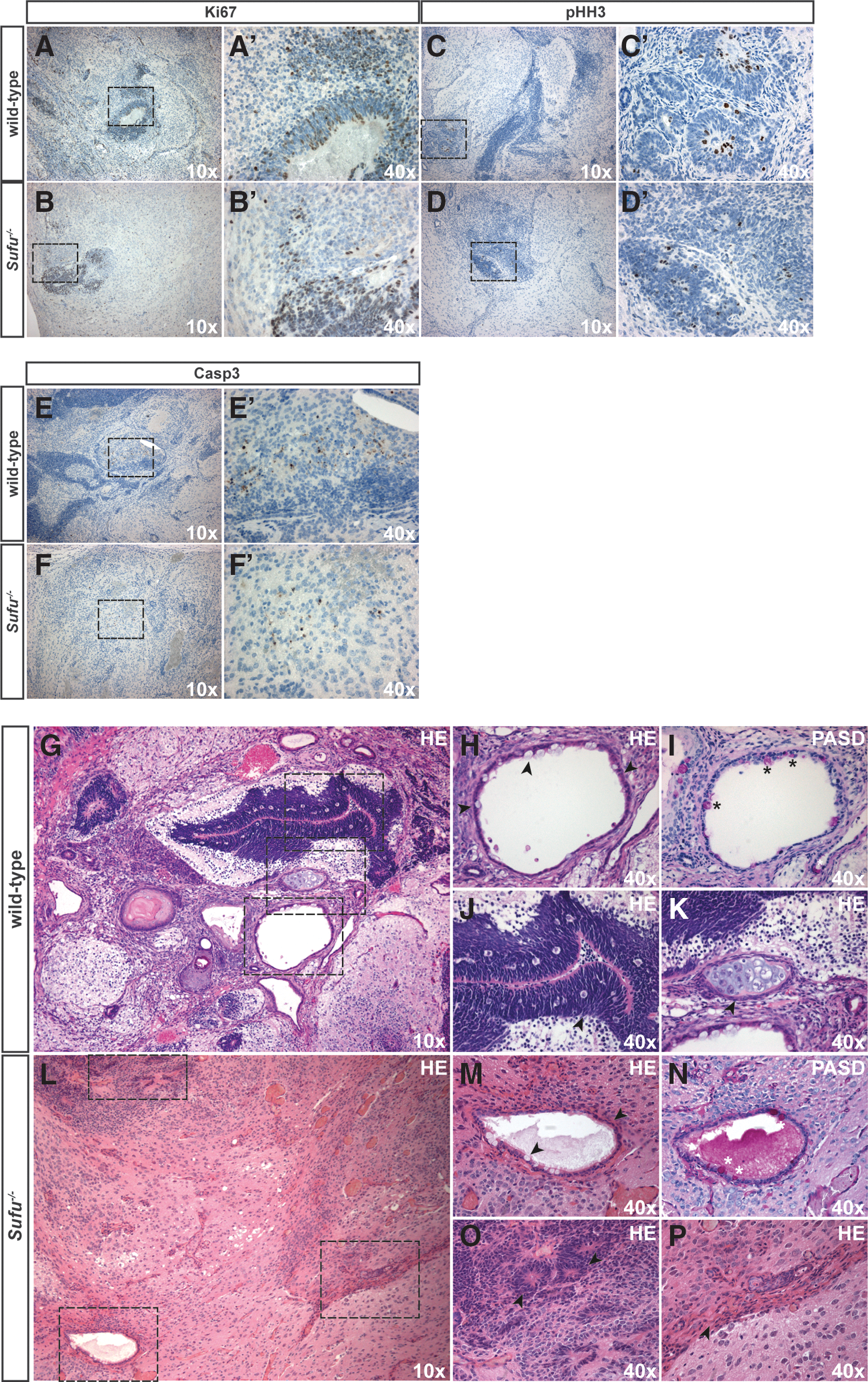
Loss of Sufu does not affect proliferation or apoptosis, but restricts mesodermal differentiation in vivo. Immunostaining of teratomas with antibodies against the proliferation markers, Ki67
To evaluate the development and composition of the different germ layers in the teratomas in more detail, we undertook a systematic histological assessment of H&E-stained tissue sections in addition to utilizing special stains and immunohistochemistry (Supplementary Table S2).
We found that wild-type tumors were characterized by a differentiation pattern agreeing with derivation from all three germ layers (Fig. 4G–K). With a similar frequency as in Sufu−/− teratomas, we observed small foci of cells consistent with the morphology of embryonal carcinoma (data not shown). The ectoderm prevailed in wild-type teratomas and was represented mainly by neural rosettes and neuropil (Fig. 4G, J). This was confirmed using immunostaining for Sox2 (Fig. 5A, A′), an early neuronal marker important for the maintenance of neural stem and progenitor cells, and synaptophysin (Fig. 5C, C′) present in presynaptic vesicles.

Teratomas from Sufu−/−
ESCs do not show chondrogenic differentiation. Immunohistochemical staining for the neuroectodermal markers, Sox2
Ciliated epithelium with or without mucous-producing goblet cells (Fig. 4H), as revealed by PAS-D staining (Fig. 4I), indicated the presence of endodermal derivatives. Endodermal epithelium was further confirmed by the presence of Keratin 5 (Fig. 5E, E′), known to be expressed in basally located cells in the pseudostratified respiratory epithelium.
Additionally, immunostaining for FoxA2, a marker for definitive endoderm, was used to verify the presence of endodermal components (Fig. 5G, G′). The mesoderm was clearly identified by the presence of cartilage and bone (Fig. 4K), adipocytes, and muscles. Furthermore, the presence of muscle was confirmed by smooth muscle actin staining (Fig. 5I, I′). In addition, Sox9 staining (Fig. 5K, K′) was used to identify proliferating chondrocytes and Alcian blue staining (Fig. 5M, M′) revealed the presence of glycosaminoglycans in the chondroid matrix.
While wild-type cells differentiated as expected, teratomas developing from Sufu−/− ESCs showed a more restricted pattern of development (Fig. 4L–P). As in wild-type teratomas, despite the reduced ectodermal differentiation detected in vitro, neuroectodermal lineages dominated and stained positive for Sox2 (Fig. 5B, B′), synaptophysin (Fig. 5D, D′), and Sox9 (Fig. 5L, L′), a marker not only involved in chondrogenesis but also expressed in neural stem/progenitor cells.
The endoderm was represented by ciliated epithelium with and without mucous-producing cells, as revealed by PAS-D (Fig. 4N) and Alcian blue staining (Fig. 5N, N′). Endodermal epithelium and visceral endoderm were present as revealed by Keratin 5 (Fig. 5F, F′) and FoxA2 (Fig. 5H, H′) immunoreactivity, respectively. The mesoderm was represented in the teratomas from Sufu−/− ESCs by muscle and adipose (Figs. 4L, P, and 5J, J′) tissue.
Strikingly, whereas around half of the teratomas from wild-type ESCs contained cartilage and bone structures (Supplementary Table S3), these were not observed in any of the Sufu−/− ESC-derived tumors when screening for Alcian blue (Fig. 5M–N′) and Sox 9 (Fig. 5K–L′). Taken together, our data indicate that Sufu is an important determinant in controlling the mesodermal lineage differentiation process in vivo.
Sufu−/− ESCs can be differentiated into chondrocytes and osteocytes in vitro
Since loss of Sufu prevented the formation of cartilage and bone tissue in teratomas in vivo, we were interested in knowing whether Sufu−/− ESCs were able to differentiate into chondrocytes and osteocytes in vitro. To this end, we applied a differentiation protocol that is based on ATRA treatment of ESC-derived EBs in suspension culture to induce the formation of MSCs, followed by plating on gelatin and induction of chondrocyte or osteocyte cell fate using a lineage-specific differentiation medium (Fig. 6O) [20].
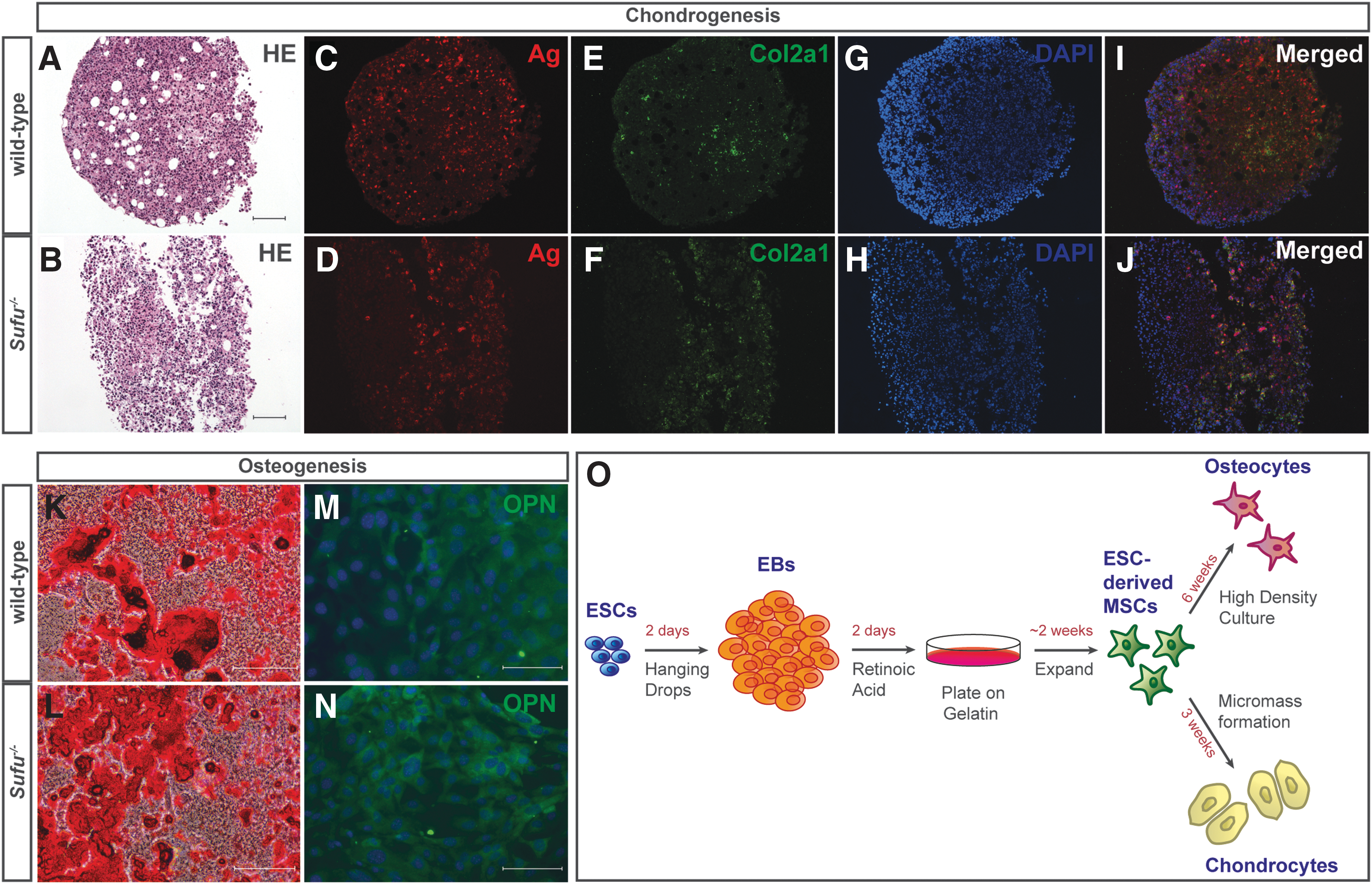
Sufu−/−
ESCs can differentiate into chondro- and osteocytes in vitro.
For chondrocyte differentiation, we formed cell micromasses in conical tubes and cultured them in differentiation medium. After 3 weeks, they were collected, fixed in formalin, and sectioned for subsequent analysis. Both wild-type and Sufu−/− EBs differentiated into MSCs and those derived from wild-type cells were able to form micromasses and adhere to each other during chondrocyte formation. In contrast, we observed that micromasses lacking Sufu were fragile and had a tendency to break up into several sister chondrocyte pellets during the differentiation process.
To promote differentiation to the osteocyte lineage, EB-derived MSCs were cultured at high density in osteogenic differentiation medium for a period of 6 weeks, followed by fixation and subsequent staining. As opposed to the chondrocyte differentiation, we could not detect any obvious differences between wild-type and Sufu−/− cells in phenotype and growth behavior.
Given the lack of cartilage and bone in the teratomas, we were surprised to find that both wild-type and Sufu−/− cells were able to differentiate into chondrocyte and osteocyte lineages. Chondroctyes were identified by the presence of aggrecan (Fig. 6C, D) and collagen 2a1 (Fig. 6E, F), extracellular matrix proteins found in cartilage, and Alizarin red indicated calcium deposition typical for ostoeogenic differentiation (Fig. 6K, L). Additionally, osteocytes were confirmed by the presence of osteopontin, a component of the mineralized extracellular matrix of bones (Fig. 6M, N). Thus, in the absence of Sufu, chondrocytes and osteocytes can be established from ESCs in an in vitro setting.
Discussion
Sufu is an essential repressor in the mammalian Hh signaling pathway as loss-of-function mutations in Sufu result in the upregulation of Hh signaling activity and mid-term embryonic lethality in mice [10,11]. To our knowledge, the current study provides the first investigation of a role for Sufu in murine ESCs, addressing pluripotency, lineage choice, and differentiation. For this study, we established ESC lines from preimplantation Sufu−/− and wild-type embryos and showed that when in an undifferentiated state, both wild-type and Sufu−/− ESCs were similar with respect to their morphology and expression of pluripotency markers.
We demonstrate that the Hh signaling pathway is activated in Sufu−/− ESCs, as indicated by a significant increase in Gli1 expression. Ptch1 showed an upward trend, but did not reach statistical significance. However, despite activated Hh signaling, there was no increase in spontaneous differentiation, demonstrating that in the absence of Sufu, ESCs are able to maintain their pluripotent character. This result agrees with other studies, which showed that neither the overexpression nor addition of exogenous Shh to cultures of huESCs or mESCs affected their undifferentiated state [8,9].
Although Gli1 expression was augmented significantly in undifferentiated Sufu−/− ESCs, this increase was surprisingly modest compared with the situation in MEFs where there was a much stronger upregulation of both Gli1 and Ptch1 mRNA when Sufu was ablated (Supplementary Fig. S1). Interestingly, the expression of Gli1 and Ptch1 in both wild-type and Sufu−/− ESC lines was higher than in wild-type MEFs.
These results indicate that the basal level of Hh signaling is much lower, or even absent, in already specified cell types, supporting the idea that Hh signaling in adult tissue is confined to stem and progenitor populations within various organs [36]. Hh signaling is essential for maintaining adult stem cell function [36,37] and the addition of exogenous Shh to mESCs was shown to regulate self-renewal [9]. Thus, a certain basal level of pathway activation might play a role in the self-renewing capacity of ESCs and other stem/progenitor cells.
However, since the Hh signaling activity in ESCs was lower compared with that seen in MEFs in the absence of Sufu, we surmise that undifferentiated mESCs are not competent to mount a full Hh response. Furthermore, we speculate that the presence of ESC-specific factors could dampen Hh signaling and/or that crucial factors are absent and are only introduced later once the ESCs start to differentiate. The identity and characteristics of the restrictions that control Hh signaling activity in ESCs remain to be elucidated.
We detected an increase in Gli1 expression in early differentiated wild-type EBs compared with undifferentiated ESCs, which later slightly decreased. A similar pattern of Gli1 expression was observed for the early differentiation of Sufu−/− ESCs into EBs, but it is worth noting that the level of Gli1 mRNA was higher (although not statistically significant) in early and late Sufu−/− EBs compared with wild-type EBs. Ptch1 was upregulated in both wild-type and Sufu−/− ESCs upon differentiation and remained high after 12 days in suspension culture. Furthermore, the Ptch1 expression level was significantly higher in Sufu−/− EBs in both early and late stages compared with wild-type EBs.
Our findings indicate in concordance with others [7,8] that as soon as ESCs start to differentiate, the restrictions that dampen Hh pathway activation in the pluripotent state are lost and the Hh pathway can be further activated. Thus, the effect of Sufu ablation in differentiating EBs can be observed as an increase in both Ptch1 and Gli1 mRNA expression.
As activation of Hh signaling is required for specification into neuroectoderm [23], we were surprised to see a reduced expression of Sox1 and Fgf5 in Sufu−/− EBs. Mesoderm and endoderm development, at least as assessed by the markers analyzed, seemed, however, in this setting to be unaffected by Sufu loss.
Previously, it was found that EBs deficient in Ihh or Smo were smaller than wild-type EBs [23]. Unexpectedly, we also found that Sufu−/− EBs were significantly smaller than their wild-type counterparts, and it is interesting that both aberrantly reduced and aberrantly increased pathway activities result in a similar phenotype. Initial EB formation and growth, however, were unaffected as the size difference only became obvious after 6 days in suspension culture. We noticed that later stage wild-type EBs appeared to have an increased apoptotic core, probably resulting from lack of nutrition influx due to the increased size. As we could not detect any apparent proliferative or apoptotic disadvantage of Sufu−/− EBs, we speculate that a skewed differentiation program, as indicated by the reduced neuroectodermal lineage, could account for the observed phenotype.
In addition, we used teratoma formation to study the ability of Sufu−/− ESCs to differentiate in vivo and demonstrated that tumors developed with similar efficiency, latency, and growth rate irrespective of ESC genotype. This was somewhat surprising as Hh signaling controls proliferation [38], and aberrant activation leads to increased tumor growth [39]. However, teratomas comprise heterogeneous cell populations, and full activation of the pathway might not occur in all cell types represented and/or might lead to opposing proliferative outcomes as revealed for the differentiating Sufu−/− EBs. Since the teratoma size was limited to 1 cm in diameter, we were unable to investigate any long-term effects of Sufu loss on tumor growth.
Teratomas originating from wild-type ESCs contained representatives of all three germ layers, although neuroectodermal tissue prevailed. The endoderm and mesoderm were represented by ciliated epithelium, with or without mucous-producing goblet cells, muscles, adipocytes, cartilage, and bone, respectively. Sufu−/− teratomas also included derivatives of the endodermal and, to a certain extent, mesodermal cell lineages. The neuroectodermal contribution in Sufu−/− teratomas was comparable with that seen in wild-type teratomas, indicating that the lower level of ectoderm induction observed during EB differentiation is sufficient to induce ectoderm formation in vivo.
Recently, it has been shown that Sufu is crucial for endochondral bone formation, regulating chondrocyte differentiation and proliferation [12]. Thus, although we did not observe any changes during mesoderm development in vitro, we expected to possibly observe some changes in these relatively late differentiation processes in Sufu−/− teratomas compared with wild-type teratomas. However, the changes were perhaps more prominent than expected since none of the tumors that originated from Sufu−/− ESCs developed cartilage or bone, indicating that Sufu plays a pivotal role in already early mesodermal differentiation processes in vivo.
One difference between our study and that of Hsu et al. [12] is that in our ESCs, Sufu was absent in undifferentiated ESCs and its absence was maintained throughout the differentiation program, while Hsu's group deleted Sufu using cre recombination driven by the Col2a1 promoter expressed in differentiating chondrocytes.
When we exposed our cells to a directed in vitro differentiation protocol, chondrocytes and osteoctyes could be obtained from both wild-type and Sufu−/− ESCs. However, this is not particularly surprising as the regulatory cross talk between cells and their microenvironment is ablated in this situation, while exposure to possible differentiation factors present in the culture media might compensate for the loss of Sufu function.
In conclusion, we have shown that Sufu−/− ESCs can be kept undifferentiated in culture, express classical pluripotency markers similar to wild-type ESCs, and form EBs in vitro. In contrast, a pronounced increase in Hh pathway activity is detected in Sufu−/− EBs during early differentiation and their size is reduced at later stages. Sufu−/− ESCs can also form teratomas in vivo, but here fail to develop cartilage and bone, suggesting a crucial role for Sufu in mesodermal differentiation processes. Furthermore, our findings emphasize the importance of in vivo studies as chondrogenesis and osteocyte development were perturbed in vivo, but not in vitro. Our results have opened up further avenues for future study, including investigations into the cause of the size reduction of the Sufu−/− EBs and the role of Sufu in cartilage and bone differentiation programs.
Footnotes
Acknowledgments
The authors thank Z. Alekseenko for invaluable advice concerning ESC derivation, Å. Bergström for technical assistance during injections, C. Lundmark, T. Schröder (Laboratory for morphologic phenotype analysis; FENO), and M. Sagici (ZeMac histoanalys AB) for teratoma and EB embedding and sectioning, and S. Lang for language editing. The authors also acknowledge the Karolinska Center for Transgene Technologies for help with MEF cells and general advice on ESC culturing.
Part of this study was performed at the Live Cell Imaging unit/Nikon Center of Excellence, Department of Biosciences and Nutrition, Karolinska Institutet, Huddinge, Sweden, supported by grants from the Knut and Alice Wallenberg Foundation, the Swedish Research Council, the Centre for Innovative Medicine, and the Jonasson donation to the School of Technology and Health, Kungliga Tekniska Högskolan, Huddinge, Sweden.
In addition, the authors thank L. Hoaglin from the Histology Core of the Charles C. Gates Center for Regenerative Medicine and Stem Cell Biology, University of Colorado, for processing and sectioning chondrogenic micromasses.
This work was also supported by the Swedish Research Council (S.T., R.T.), Magnus Bergvall's Foundation (S.T.), and Karolinska Institutet's Research Foundations (S.T.), the Swedish Cancer Society (R.T.), the Wallenberg Consortium North (B.R.), and the Robert Lundberg Memorial Foundation (K.H.-H.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.