
Abstract
T-Box transcription factors are expressed throughout the gestational period and coordinate a variety of embryonic events that enable proper development, from the first differentiation of embryonic and extraembryonic tissues until final organogenesis. Although the T-Box gene family comprises essential roles in early cellular differentiation, in adult tissues it has also been associated with cancer development. In spite of their common T-Box regulatory binding domain, T-Box family members utilize different cofactors and different spatiotemporal expression patterns to confer their specificity in diverse developmental processes. The earliest expression note of T-Box factors can be observed even before fertilization in primordial germ cells and just after zygotic gene activation (around the eight-cell blastomere stage). Thus, particularly the early stages of development are highly influenced by these key regulators in line with the notion that T-Box mutations lead to developmental disorders and even lethality. In this review, we summarize recently acquired findings on T-Box factors to provide a comprehensive overview on their role during early embryogenesis.
Introduction
E
A prominent family of transcription factors acting in both embryogenesis and organogenesis is the T-Box gene family [1]. In the 88 years since the identification of the first T-Box gene by random mutagenesis [2], embryologists have concentrated efforts in depicting the pathways in which T-Box transcription factors are involved.
Most of the initial work, as early as in 1927, was based on the careful morphologic analysis of Brachyury (T) mutant mouse embryos presenting a Brachyury large deletion. Severe developmental impairments and embryonic lethality exhibited by heterozygous and homozygous mice, respectively, indicated the relevance of T during development [3 –5]. Advances in molecular techniques permitted a deeper insight into T-Box protein functions. Besides gene expression patterning (Fig. 1) that can provide information about the spatiotemporal role of a particular gene, gain- and loss-of-function studies significantly increased our understanding about the role of T-Box factors, especially during the early stages of development. However, much of the T-Box regulators, target genes and downstream pathways, remain to be identified.
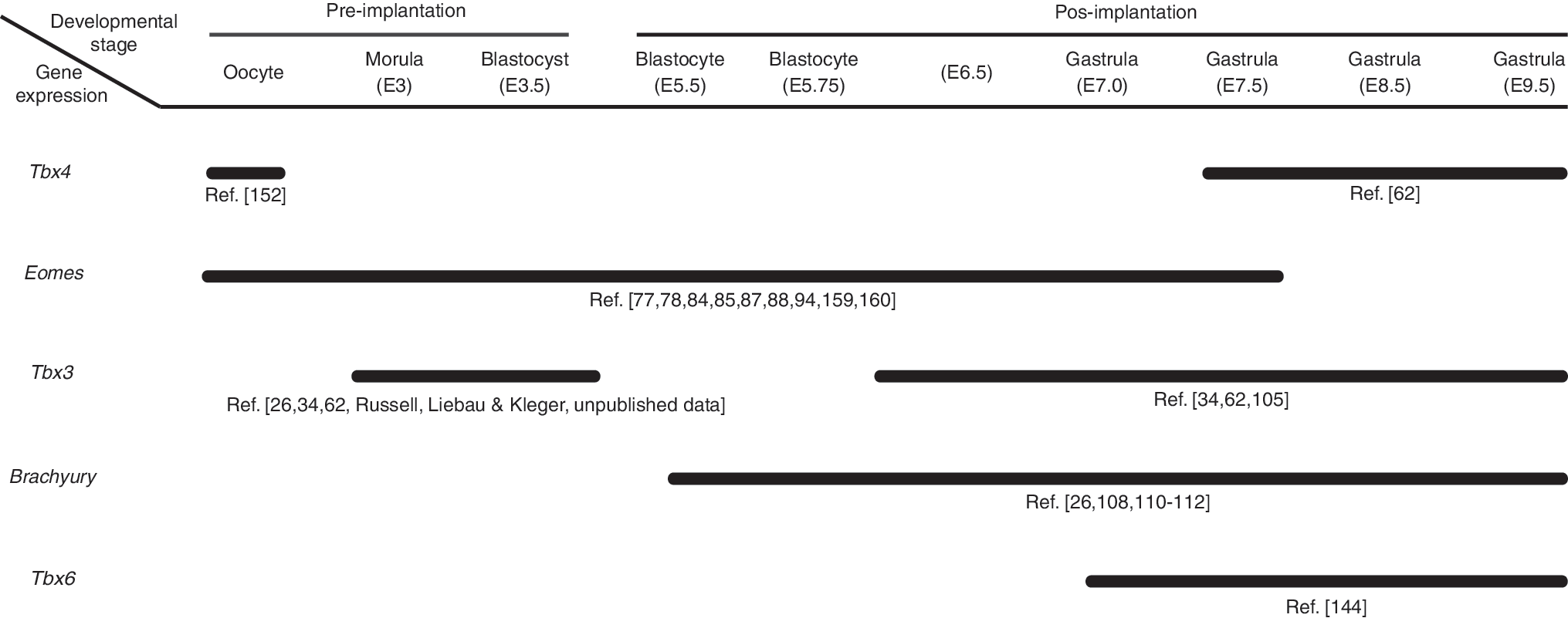
T-Box gene expression in the early phases of mouse development. Maternal mRNA expression is seen in Oocytes for Tbx4 and, in oocytes and preimplantation embryo for Eomes. Tbx4: [62,152], Eomes: [77,78,84,85,87,88,94,159,160], Tbx3: Russell, Liebau & Kleger (unpublished data) [26,34,62,105], Brachyury: [26,108,110 –112], Tbx6: [144]
Being a highly conserved family of proteins, findings on their vertebrate orthologues constitute an invaluable source of information about human embryogenesis.
Finally, the T-Box family has been extensively studied and therefore also discussed in many different reviews, which compile very diverse aspects of the transcription factor family. This review focuses on the role of T-Box factors during early embryo development by covering the first nine Carnegie stages, from fertilization to late-gastrula stage. Although “T-box genes in early embryogenesis” [6] targets the same development period, our aim is to serve as a complement, presenting recent findings on the field. Moreover, “The T-box gene family: emerging roles in development, stem cells and cancer” [7] is a very recent and complete review, also covering organogenesis and adult roles of T-Box genes.
Early Cell Fate Determination in the Embryo
Normal embryonic development undergoes numerous stages of cellular proliferation, programmed death, migration, and differentiation. Starting from oocyte fertilization, embryogenesis proceeds with zygotic cleavage, resulting in two blastomeres (two-cell stage) and subsequent mitotic events and forming a compact sphere, the morula (sixteen-cell stage). The first evidence of morphological changes occurs in the morula when the blastocyst evolves (thirty two-cell stage). These earliest patterning events are dependent on transcription factor-based cell fate specification events, particularly governed by caudal-type homeobox protein 2 (Cdx2) and octamere-binding transcription factor 3/4 (Oct3/4). At the eight-cell stage, both factors are co-expressed but repress each other when at the blastocyst stage two different lineages become evident, the Oct3/4 and Tbx3-positive inner cell mass (ICM) and the Cdx2-positive trophectoderm (TE) [8].
Before the implantation into the uterus when the development of the late blastocyst occurs, the ICM splits into two distinct cell types, the epiblast and the primitive endoderm (PE). Similar to the reciprocal interaction of Oct3/4 and Cdx2, Nanog, Tbx3, and Gata6 specify these two lineages in a most likely mutually exclusive fashion, with Nanog the epiblast and Gata6 the PE [9,10]. While the cells from the epiblast will originate from the three embryonic germ layers and, subsequently, the embryo itself with just a few extraembryonic structures, the PE and TE give rise to extraembryonic tissues being essential for embryo implantation. However, at the molecular level, it has been reported that the induction of embryonic development is modulated by extraembryonic signals [11]. Shortly after implantation, early molecular asymmetries instruct the proximal–distal axis of the embryo to form the egg cylinder stage of the embryo. Subsequent movements and patterning events of the extraembryonic endoderm and the visceral endoderm instructed predominately by gradients of WNT-, BMP-, and NODAL signaling finally establish the anterior–posterior polarity of the embryo. Interestingly, the regional differences are primarily established by gene expression differences prepatterning the morphologically rather symmetrical embryo [12 –15].
The next obvious morphological changes occur when epiblast cells congeal at the posterior pole of the epiblast to form the primitive streak (PS), a characteristic groove structure marking the gastrulation phase of mammalian development. The PS is shaped with the migration of epiblast cells that are later committed to the mesendodermal lineage, while the nascent mesoderm arises via Epithelial–Mesenchymal Transition (EMT) and forms a new layer between the remaining epiblast and the covering visceral endoderm [16]. The first evidence of streak formation can be observed at the proximal posterior pole of the epiblast and progresses to the distal tip of the embryo. Here, spatiotemporal streak formation defines the different derivates. The posterior mesoderm gives rise to the extraembryonic mesodermal tissues. The intermediate and anterior streak originates from the lateral plate, paraxial and cardiac mesoderm while the anterior streak forms the mesendoderm. This common germ layer comprises the prechordal plate, the notochord and the node, and the definitive endoderm. The remaining epiblast cells form the ectoderm. The regionalization of the PS is due to gradients of NODAL signaling showing the highest levels at the posterior proximal pole of the embryo [11,17,18]. As gastrulation proceeds, the embryo continues to fold by cell migration and a cellular rod initiates to establish the forthcoming notochord transient structure. The central localization of the notochord (from anterior to posterior side) is correlated to its function. This cell network releases factors that have an influence on tissue specialization on its surroundings and provides a primitive axis for the embryo development.
At late gastrulation, a neural plate is formed above the notochord from the dorsal ectoderm (neuroectoderm). This is the neurulation onset, a process that later continues with folding of the neural plate and its detachment from the ectoderm to give rise to the neural tube, a precursor of the brain and spinal cord [19].
Concomitantly to neural tube formation, its adjacent mesoderm (paraxial mesoderm) starts subdividing into somites, segments that will generate muscles and skeleton. Somitogenesis initiates cranially during gastrulation and extends in the course of the next developmental stages [20 –22].
The early development phase is accomplished by the beginning of the fourth week, when neurulation is concluded, and it is followed by organogenesis with posterior maturation of organs. Figure 2 provides a summary of the early T-Box gene expression covered in this review, whereas the family gene expression during organogenesis is discussed elsewhere [23].
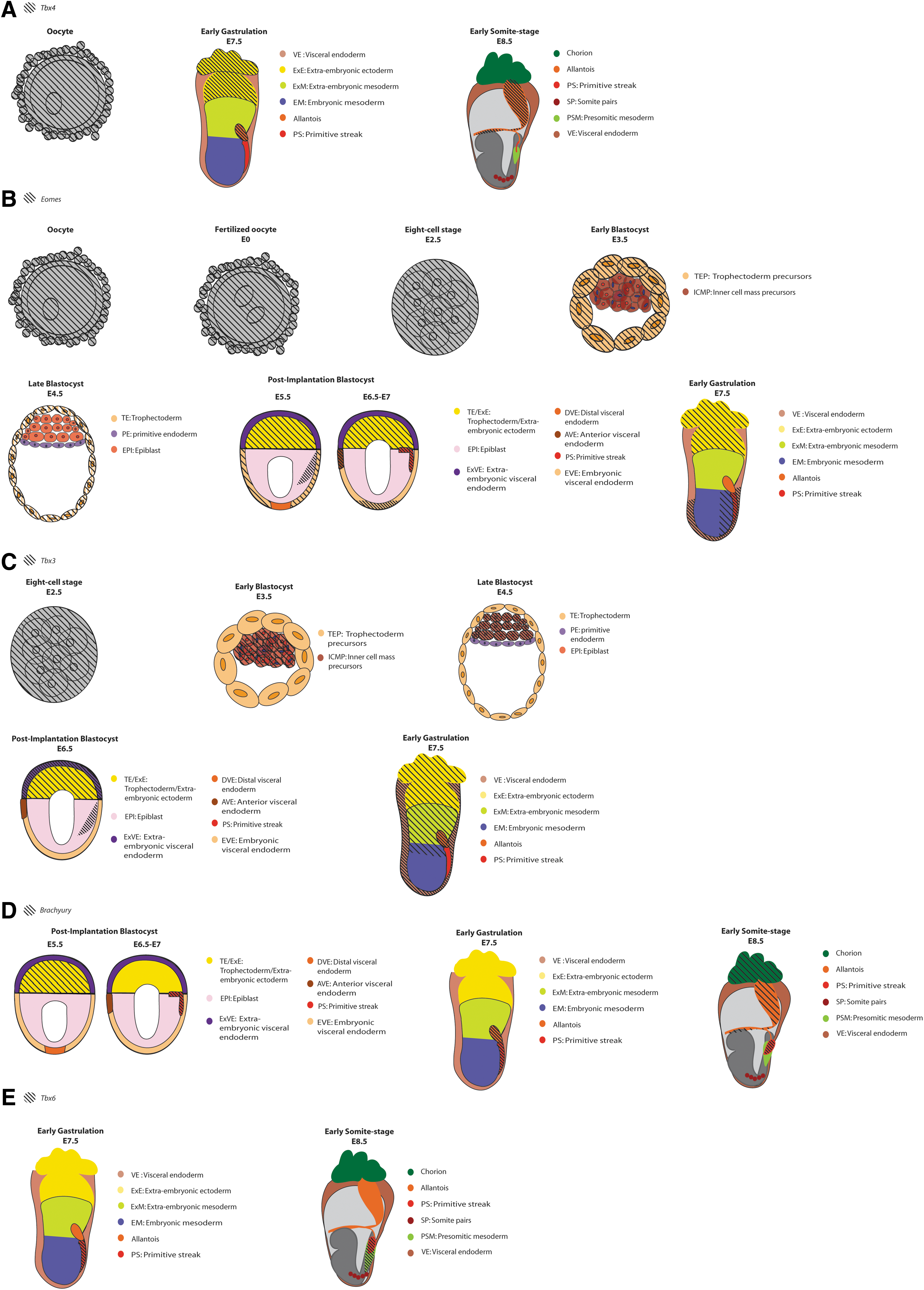
Schematic of T-Box gene expression in early mouse development stages. Embryo age in embryonic days (E) is indicated below each stage. References can be found in Figure 1.
The T-Box Gene Family
The human T-Box gene family consists of seventeen genes, divided into five subfamilies according to their phylogenetic proximity (reviewed by Naiche et al. [23]). A plethora of knowledge has been gathered about the T-Box gene family through the investigation of its emblematic member, Brachyury (T), the first to be discovered and, by far, the most studied. The relevance of this gene to embryogenesis was unveiled by Dobrovolskaïa-Zavadskaïa [2] with the description of a heterozygous deletion that produces tailless or short-tailed mice. Further evidence of Brachyury activity on developmental processes has been reported by Chesley [24] demonstrating in utero lethality in homozygous mutated mice at embryonic day 11 (E11).
Interestingly, the initial discovery of the T-Box gene family has been reproduced in various species and demonstrated high conservation through evolution. In fact, phylogenetic analyses indicated ancestor gene duplications during early evolution of vertebrates and a posterior divergence resulted in the current known gene family [25]. A conserved peptide motif among Brachyury homologous proteins led Bollag et al. [26] to classify the corresponding genes into a family, the T-Box gene family. Each member of this family exhibits a characteristic spatiotemporal expression pattern and therefore instructs specificity regarding the respective target genes. However, the variance found among DNA-binding sequences in the T-Box family members [27] does not solely explain gene target specificity, since several T-Box proteins are able to bind a Brachyury core sequence. Tada & Smith [28] discussed some aspects involved in the spatiotemporal specificity of T-Box genes, for instance, an effect mainly determined by respective interaction partners to regulate target gene activation or repression. This process is exemplified by Tbx3, whose promoter is bound by different molecules depending on its action [29,30]. As in the case of the Brachyury gene, the presence of different regulatory elements also illustrates the spatiotemporal specificity of the T-Box family. Brachyury presents both an upstream region (“T-streak” region), active in the PS and derivatives, and a trans- or long-range cis- regions, acting within the notochord [31,32].
Although accumulating evidence confirms the action specificity of the T-Box, the common binding site raises the possibility of competition between various T-Box members to regulate the same target gene, as suggested by Wehn & Chapman [33]. Such an example is given by the ectopic expression of Tbx6 in the paraxial mesoderm, which may compete with Tbx15 and Tbx18 for their target regulatory sequences [33]. In contrast, this common binding feature could present beneficial aspects in case of gene deregulation, such as a compensatory effect via activity of a closely related family member, which has an overlapping expression pattern, as reported for Tbx2/3 and Tbx4/5 [25]. Analysis of Xenopus levis gastrulation also revealed a possible compensatory mechanism of TBX2 and TBX3, as recently shown by Weidgang et al. [34].
Similar compensatory events are suggested later in development, for example, during mice lung formation. Tbx2 null mutants present deficient lung development just in the later phase of organ formation. In fact, this event coincides with downregulation of Tbx3, which shows an overlapping expression pattern with Tbx2 in the earlier phase. Furthermore, both genes contain binding sites to repress Cdkn1a and Cdkn1b, an event allowing sufficient lung formation [35].
Another example of compensatory gene function in this family is the C. elegans tbx8/tbx9. Burga et al. [36] detected a compensatory response of tbx8 for tbx9 loss, with the opposite also being valid. There, the authors, in addition, suggest that incomplete penetrance in human diseases, including those caused by T-Box gene mutations, might be a consequence of compensatory mechanisms [36].
Apart from compensatory effects, various T-Box members have been revealed to act in a dose dependent mode, such as Brachyury, which presented assorted shades of mutant phenotypes [37]. Likewise, other family members show the same dose dependence, as demonstrated by developmental disorders found in case of TBX1, TBX3, TBX4, TBX5, and TBX22 haploinsufficiency [38 –42].
This dosage sensitivity seems to be of particular relevance as spatiotemporal mis-/overexpression of T-Box factors, either entirely disrupting organ development or ascribing dominant phenotypes based on the over-dosage of the respective T-Box factor. Here, overexpression of TBX4 and TBX5 genes interferes with the normal heart and limb formation [43,44], while high levels of TBX3 [45,46] and TBX2 [46 –51] are associated with tumorigenesis. Moreover, TBX3 also ascribes pacemaking features to the working myocardium on misexpression [52]. Lastly, differences in T-Box gene expression are relevant even in adult tissues as seen for TBX15 misexpression, which is implicated with fat deposit distribution and risk of metabolic diseases [53,54].
An additional relationship between the different T-Box proteins also becomes evident early in development, when one is activating or repressing the expression of the other, for example, TBX3 is activating T and EOMES to induce mesendoderm commitment [34]. Thus, proper coordination of T-Box gene expression may affect cell differentiation and movement during embryogenesis, while receiving impact from various signaling pathways. The regulatory mechanisms underlying these processes, by which transcription factors localize to specific regions in an embryo and drive differentiation, require further investigation. Although most of the T-Box upstream regulators and targets are still unknown, technological progress in this field has been accomplished [55].
T-Box Proteins and Their Importance in Early Development
Progressive differentiation is a hallmark of embryogenesis. Although many questions regarding the molecular basis of this process remain to be answered, the morphological events occurring during early lineage commitment become more and more defined. The importance of gene expression regulatory mechanisms in embryo development is, however, a consensus (reviewed by Zernicka-Goetz et al. [56]).
According to Hamatani et al. [57], translation of maternal inherited RNA is required for a primary gene expression in the embryo, which occurs between the four- and eight-cell stages [58]. Afterward, events in morula and blastocyst formation are definitely correlated to this process as emphasized by a transition in the transcriptome pattern. This new lineage-specific signature is gradually set along with developmental pathway activation, where proper interaction between transcription factors plays a major role [59].
The dominating molecular players orchestrating the first events of life are the transcription factors. In line with highlighting this importance, the leading cause of early developmental disorders are mutations in various transcription factors [60].
Transcription factors are proteins that exhibit an interaction domain with regulatory DNA domains and/or promoters to activate or repress gene expression. Frequently, those molecules interact with other co-factors, increasing the specificity of expression control. Besides the presence of co-regulators, the degree of chromatin condensation and the existence of other mechanisms such as different regulatory sequences (cis- and trans- acting) and DNA methylation illustrate the complexity of the eukaryotic gene regulatory mechanism/network [61].
Human developmental diseases resulting from T-Box genes mutations reveal the relevance of the gene family in both embryogenesis and organogenesis. Currently, diverse developmental disorders are known to be caused by T-Box genes, due to problems such as mutations or dysregulation of gene expression (Fig. 3).
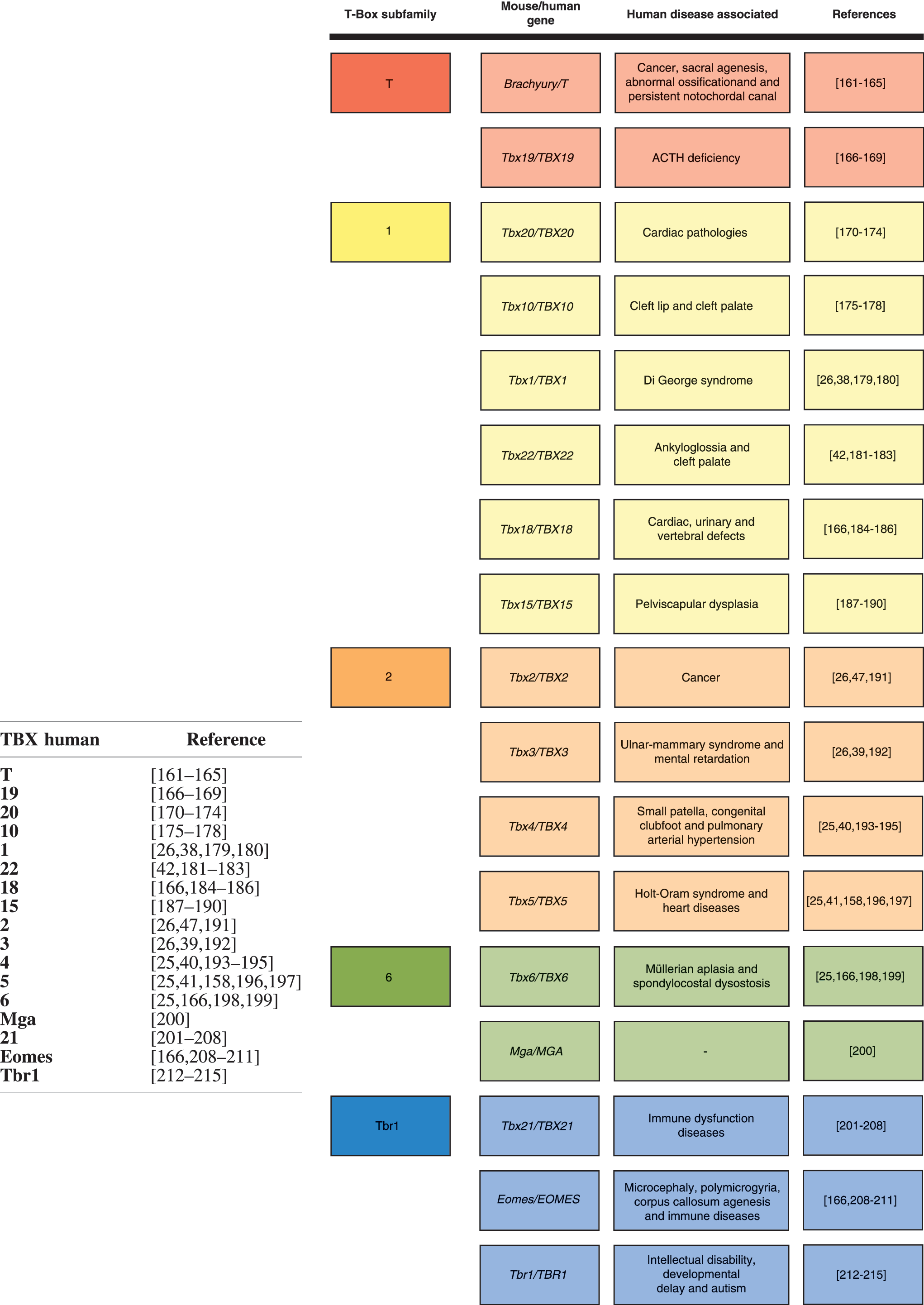
Mouse and human T-Box genes, their respective subfamilies, and human associated diseases. T-Box subfamilies are color coded, respecting phylogenetics.
Next, we are going to explore the most significant aspects of T-Box gene family in early development.
Tbx3
Tbx3 represents a unique member of the T-Box family of transcription factors due to its uniquely early expression pattern in the mouse embryo. Tbx3 expression can be observed as early as in the morula stage mimicking a “salt and pepper” pattern as previously described for Nanog (Russell, Liebau & Kleger, unpublished data). Later, Tbx3 expression localizes to the ICM [62] and later during onset of gastrulation gets restricted to the proximal pole of the epiblast in the mouse embryo [34]. Consistent with the very early expression in the ICM [62], different studies on stem cells reinforced the idea of Tbx3 being an important pluripotency factor [34,63 –66]. Interestingly, Tbx3 null mice die around developmental stage E12.5 due to both cardiac and yolk sac defects [67] and lack any defects associated with the integrity of the ICM (Liebau & Kleger, under revision). Afterward, Tbx3 is abundantly expressed in the extraembryonic visceral endoderm and visceral endoderm overlying the epiblast in pre- and mid-streak embryos [34]. This observation supports previous data reporting roles of Tbx3 toward this lineage by directly activating Gata6 expression [63]. Others [14,68,69] and we [34] have hypothesized that this expression might foster patterning events in the adjacent epiblast. We have shown in a recent study that NODAL plays a central role as an immediate target of Tbx3 actions and demonstrated that blockade of the NODAL/SMAD2 pathway significantly hampers Tbx3 inductive effects [34]. Intriguingly, the Tbx3 target Gata6 is also targeted by NODAL/SMAD2 signaling [70]. These data are well in line with the developing mouse embryo where NODAL cues get restricted during onset of gastrulation to the proximal pole of the epiblast to promote a posterior fate. Notably, Tbx3 also drives the expression of the NODAL target genes Lefty1 [71,72] and cerberus [73]. This matches the in vivo expression pattern of these genes in a discrete cell population within the visceral endoderm at the epiblast border. Nodal itself is expressed throughout this tissue layer [14]. Therefore, Tbx3 most likely helps reinforce NODAL signaling as a major effector of cell sorting in the posterior-proximal epiblast, enabling gastrulation onset. Those include a NODAL auto-regulatory feed-forward loop [74,75], Nodal-protein activation by convertases in the extraembryonic ectoderm [75], and a regulatory feed-forward loop involving Bmp4 and Wnt3/beta-Catenin signaling that transcriptionally regulates maintenance of Nodal expression [75]. Of note, extensive dispersion between the visceral and definitive endoderm is present, leading to epiblast-derived organs containing extraembryonic elements [11]. It would be interesting to know to what extent the Tbx3-positive cells within the extraembryonic visceral endoderm/visceral endoderm contribute to the endoderm-derived organs [11]. Interestingly, gastrulation does not occur solely Tbx3 dependent in mammalians; however, compensation driven by other T-Box factors is likely to mask any gross defects [67]. In fact, gastrulation defects became evident just when using lower vertebrate model systems such as the Xenopus levis. Here, we were able to show that the Tbx3-paralogue Tbx2 is able to compensate via subsequent upregulation on Tbx3 loss. Of note, despite a strong structural difference between Tbx3 and Tbx6, the latter was also downregulated when Tbx3 was overexpressed or vice versa [34]. In sum, these data may suggest the evolution of a mechanism for safeguarding normal gastrulation and subsequent development by directly and/or indirectly regulating expression of key components of streak formation involving Tbx3.
Eomes
The identification of eomesodermin (Eomes) [76], another member of the T-Box gene family, provided a significant contribution on defining the importance of T-Box genes during early development. At least to our knowledge, Eomes represents the T-Box family member having the earliest effect in embryonic development [23,77]. Corroborating to its developmental role, maternal Eomes protein was detected in mice from oocytes up to the E3.5 preimplantation stage [78]. Furthermore, Eomes has been described as a regulator of several genes related to development [79,80].
Eomes presents intricate ways to drive its expression, such as the interaction among different activators leading to the formation of a loop between promoter and enhancer, proposed by Kartikasari et al. [80]. Moreover, two different transcriptional cis- regulatory regions have been identified for Eomes in mouse stages E10.5 and E14 [81,82], and an additional region has been described as driving Eomes expression during gastrulation [83].
In the preimplantation embryo, the earliest expression domain is noted in the TE of the blastocyst and expression is ongoing in the extraembryonic ectoderm of the postimplantation embryo. Around E5.75 of mouse embryogenesis, a second expression domain can be observed located to the posterior part of the epiblast, the region of gastrulation onset. Later on, Eomes expression extends to the entire PS and nascent mesoderm [84,85].
The morula outer cell layer presents a TE progenitor nature, and it therefore exhibits a different gene expression signature when compared with its inner portion. As stated earlier, there is a reciprocal interaction between Oct3/4 and Cdx2; to specify these two lineages in the most likely mutually exclusive fashion, Oct3/4 is the epiblast and Cdx2 is the trophoblast [9,10]. Indeed, Cdx2 is a transcription factor acting upstream of Eomes. Although Eomes expression is restricted to outer cells of the morula, Eomes loss does not affect the decrease in Oct4 during TE specification. In contrast, the pluripotency state of outer morula cells is maintained when Cdx2 is absent [8]. Furthermore, Eomes mutants showed that ICM pluripotency restriction was not under direct influence by Eomes deficiency itself, but the trophoblast differentiation was impaired. Therefore, in the trophoblast, representing the placenta precursor, a lack of Eomes prevents the embryo from implantation, an additional and crucial feature of mammalian development [86 –88].
After a successful implantation of the early embryo into the maternal placenta, the postimplantation stage is preceding the gastrulation stage, which is characterized by a cellular transition from an epithelial to a mesenchymal state (epithelial–mesenchymal transition, EMT) as a result from downregulation of E-cadherin, a key protein in cell–cell adhesion. Downregulation of E-cadherin enables epiblast cells to lose their epithelial fate and migrate toward originating from the mesendoderm. In turn, formation of the ectoderm arises from the nonmigrating remaining epiblast cells. Arnold et al. [89] could demonstrate a role of Eomes in downregulating E-cadherin, while knockdown of EOMES in human embryonic stem cells (hESCs) maintained a decrease in E-cadherin levels [79]. This contradiction could be explained by additional factors, absent in vitro, that might contribute to this repression.
In Xenopus, Ryan et al. [76] described a dose-dependent Eomes induction of various mesodermal genes, revealing the essential role of these genes in gastrulation and further embryonic development. Although Xenopus studies suggested that the development of residual somatic mesoderm in mouse T mutants is just possible due to the presence of Eomes [90], increased expression of mesodermal markers on the knockdown of EOMES was observed in mammals [79].
As Eomes is disrupted, the migration of epiblast cells is impaired [89]. Thus, in mammals, Eomes expression is likewise important during gastrulation, but differently from what occurs in Xenopus, Eomes plays a central role in repressing mesodermal genes while favoring definitive endoderm differentiation [79].
Mammalian mesodermal lineage specification is accomplished in Eomes mutant cells, whereas epiblast cell migration toward PS is defective. Thus, privation of proper PS formation rather than Eomes deficiency is responsible for the block of further gastrula development. Although mesodermal cell differentiation is maintained, Eomes deletion leads to complete absence of definitive endoderm [79,89]. Studies in zebrafish corroborate the fundamental role of Eomes in endoderm specification, while genes involved in mesoderm formation have a strong alternative path for their downstream activation [91]. It is still unknown whether epiblast cells migration per se provides additional signals to induce endoderm commitment but, as demonstrated by Tada et al. [92], mesoderm cell presence is essential during endoderm specification in vitro. In addition, differently from what happens during TE differentiation, Teo et al. [79] describe the participation of EOMES in the inhibition of pluripotency gene expression (NANOG, OCT4, and SOX2), directing endodermal fate, but highlight the downregulation failure when differentiating hESCs. Taken together, the positioning of mesoderm progenitors via migration is likely to provide the missing cue to endoderm differentiation observed by Tada et al. [92] and Teo et al. [79] in vitro.
Recently, a model accumulating the several molecules playing a role in EOMES expression regulation was proposed, with subsequent definitive endoderm specification and mesodermal inhibition [79].
Before gastrulation, a defining change in the embryo axis has to take place. The antero-posterior axis is shaped postimplantation, as the anterior visceral endoderm (AVE) migrates toward the embryonic-extraembryonic boundary of the blastocyst. The resulting AVE cell arrangement and its subsequent influences on signaling are a prerequisite to set the PS emergence location [93].
The AVE cells express NODAL, BMP, and WNT inhibitors, establishing an embryo anterior microenvironment that is regarded as ideal to prevent signaling for mesoderm allocation in this region, thus restricting PS and mesodermal fate to the proximal posterior portion of the gastrula [68,94 –99]. Eomes role in this process was assessed by its selective deletion in the visceral endoderm [94]. The authors concluded that NODAL signaling from epiblast cells is regulating Eomes expression, and that Eomes promotes AVE specification and migration. Previously, extraembryonic ectoderm was found to repress the ectopic expression of AVE genes, and to play a role in migration of those cells [100]. Eomes mutant embryos with epiblast-restricted NODAL signaling permitted distal visceral endoderm cell migration, suggesting that Eomes is essential for the AVE differentiation. But it seems to play a minor role in migration. Therefore, loss of migration could be mediated by an imbalance of NODAL signaling, apparent in the absence of extraembryonic ectoderm or through a possible loss of a NODAL feedback inhibition exhibited by Lefty, an Eomes downstream molecule [94].
The importance of NODAL in development is highlighted by Ben-Haim et al. [75], where they proposed that the decision fate into either mesoderm or endoderm might ensue according to the magnitude of exposure to NODAL signaling. Accordingly, Tam & Loebel [101] found that different levels of NODAL activity throughout PS are required for mesoderm and endoderm generation.
Mesodermal/Endodermal decision can also be related to TBX3, as outlined. TBX3 protein is expressed in extraembryonic ectoderm and visceral endoderm and activates Eomes and Nodal promoters [34]. Further regulation of Eomes expression is achieved by TBX3 on an epigenetic level. The association of TBX3 to a histone demethylase (Jmjd3) provides chromatin restructuring at the enhancer element of the Eomes locus that enables enhancer–promoter interactions and, ultimately, drives endodermal fate [80].
Thus, in line with what was presented here and as suggested by Weidgang et al. [34], TBX3 could be considered the extraembryonic ectoderm cue linking migration to Nodal and Eomes expression. Such a scenario is supported by recent studies on gastrointestinal cancer. Here, NODAL signaling drives EMT and migratory properties in pancreatic cancer, while Tbx3 does similarly in colorectal cancer [102 –104]. Furthermore, low levels of BMP have been proposed to be responsible for AVE specification [94], and alterations in Bmp4 expression were linked to mutation and overexpression of TBX3 [34,105].
Although NODAL/BMP/WNT-signaling pathways are already recognized as essential during germ-layer formation, their interactions with T-Box proteins and the molecular mechanisms underlying cellular migration and differentiation need further investigation.
Of note, mesodermal lineage specification remains accomplished in Eomes mutant cells as judged by the expression of key markers Brachyury and Bmp4 labeling posterior and extraembryonic mesoderm tissue [89]. In line with the observation that Brachyury may label a common mesendodermal progenitor, T-positive cells from the posterior streak (properly induced in Eomes mutants) fail to anteriorize, leading to the mentioned lack of definitive endoderm formation. Lineage tracing using an Eomes-cre allele in combination with a Rosa LacZ reporter nicely illustrates the derivatives of various waves of Eomes-positive cells arising early and late during PS formation [106]. Here, the authors of this brilliant study defined context- and time-dependent, cell-autonomous roles of Eomes as giving rise to the prospective cranial and cardiac mesoderm and to the definitive endoderm, node, and notochord. Here, low levels of NODAL signaling at the posterior streak allow Eomes to activate Mesp1/2, labeling the earliest cardiovascular progenitors, and to promote EMT and subsequent cell migration. In contrast, the second wave of Eomes-positive cells arising at the anterior streak requires high levels of NODAL to form definitive endoderm, node, and notochord [106]. These data are reflected in a simultaneously published in vitro study using ESCs [107].
Another role of Eomes during early development becomes apparent slightly before gastrulation, when Eomes functions during pluripotency inhibition and mesendoderm promotion [79], as already mentioned in this section. Here, EOMES governs much of the transcriptional network characterizing endodermal cells in the presence of high NODAL/ACTIVIN signaling [79], whereas low levels instruct the earliest cardiovascular progenitor cells [107]. Thus, a series of both in vitro and in vivo studies in various species complement each other to define not only a time- and cell-autonomous but also noncell autonomous scenario, where Eomes governs gastrulation and, subsequently, a variety of endodermal and mesodermal derivatives [79,92,106,107].
Most likely, the known gradients of NODAL signaling, generated by various sources of the early embryo, enable the subsequently arising waves of Eomes-positive progenitor cells in different streak regions and give rise to the progeny mentioned earlier. Such a scenario may be also possible for Tbx3 but despite several, independent studies reporting Tbx3 in an upstream position of Eomes [34,80] the lack of a relevant gastrulation phenotype in Tbx3 mutant mouse embryos points to a better secured mechanism (compared with Eomes). This is most likely because of compensating events of other T-Box factors as outlined by Weidgang et al. [34]. However, also extrinsic signaling clues such as WNT- and BMP-signaling, both of which are altered on Tbx3 expression, may mask a Tbx3 phenotype during gastrulation. Thus, intelligent mouse models allowing spatiotemporal deletion of T-Box genes as performed in the study by Costello et al. [106] are warranted. These will help in further delineating the various functions of Tbx3 and Eomes during early development and germ layer formation. Particularly, NODAL/BMP/WNT signaling pathways must be taken into account when generating respective models as their interactions with T-Box proteins and the molecular mechanisms underlying cellular migration and differentiation still remain incompletely understood.
Brachyury (T gene)
As previously mentioned, initial studies on T-Box gene were concentrated on the first identified family member, the Brachyury (Greek for “short tail”) or T gene.
Brachyury posterior expression in nascent and migrating mesendoderm as well as decreasing Brachyury levels in cells migrating away from the PS leads to the conclusion that this gene plays a crucial role in formation of the mesoderm in the posterior embryo [108,109].
Classically defined as an early mesoderm marker, the first Brachyury gene expression was for several years reported to be found in the proximal/posterior epiblast cells, the future PS site [108]. However, an even earlier Brachyury expression domain (E5.5) was believed to be detected in the extraembryonic ectoderm [110 –112], raising the possibility of Brachyury role in the extraembryonic ectoderm. In addition to the easily perceived tail absence phenotype, complete loss of posterior mesoderm, and abnormalities in allantois, posterior somites and notochord characterize the Brachyury homozygous deletion mice [3,113,114]. In view of this, the next sections will focus on the different embryonic and extraembryonic tissues that are impacted by Brachyury.
Allantois
Allantois is the posterior PS-extension, representing an extraembryonic mesoderm structure that, after fusing with the chorion, gives rise to the chorioallantoic placenta, which will, in turn, account for umbilical vessel formation, thereby contributing to embryonic vasculogenesis. Allantois defects were initially regarded as an outcome of the failure in mesodermal differentiation, since Brachury expression was at first not detected [115]. Along with detailed investigations on various gastrula stages, Brachury expression showed though that at least the allantois core presents Brachury expression [112]. Further studies accessed the gene role in murine allantois development, revealing a decrease in allantois cell proliferation and death of core cells in C-terminal truncated mutants. With arrest of allantois elongation, its fusion to the chorion is prevented apart from the fetoplacental vascularization [116]. Despite causing allantois elongation failure, the Brachury mutation does not interfere in (allantois derived) chorio-adhesive mesothelial cell differentiation, and from this perspective it has no influence in chorion-allantois fusion. However, the chorion Brachury expression during fusion draws attention to a possible role of the gene in this process [112,116].
Somitogenesis
Apart from its extraembryonic role in allantois formation, Brachyury expression significantly affects mesoderm formation. As mesoderm differentiation progresses, two anterior PS populations are formed, namely, axial and paraxial. The paraxial mesoderm is segmented into cell blocks called somites that will, ultimately, generate smooth muscle and cartilage, among other tissues. Somitogenesis is characterized by a progressive switch in gene expression patterns favoring the differentiation process of presomitic mesoderm (PSM). These mesodermal cells are localized in the embryonic posterior end, and their differentiation occurs during migration toward the embryo head. The maintenance of the PSM cellular pool and their identity transition are governed, among others, by FGF and WNT, two imperative signaling pathways in embryogenesis [117,118]. Several models provide evidence about this relationship, even when not direct, between the T-Box gene Brachyury and Tbx6 and FGF as well as WNT-signaling pathways. Canonical WNT signaling has been reported as essential for somitogenesis [119]. Along with the demonstration of shared Wnt-3a and residual somites phenotype of Brachyury loss-of-function mutants [120], additional experiments using transgenic mouse embryos and murine ESCs confirmed WNT as being a regulator of Brachyury in paraxial mesoderm cells [121,122]. These factors work synergistically with both Brachyury and Tbx6 in coordinating presomitic gene expression pattern and therefore somitogenesis [123]. Further experiments with zebrafish allowed access to the role of Brachyury orthologues (ntl/bra) in maintaining progenitor cells during somitogenesis via a positive regulatory loop with WNT signaling [119].
Likewise involved in the complex network ensuring proper somitogenesis, the FGF signaling activates Brachyury homologs in both invertebrates [124] and vertebrates [125]. FGF signaling, which is stronger at the caudal end of the embryo, activates Brachyury and Tbx6 (it will be addressed later) to maintain the progenitor identity of these cells and progressively decreases as differentiation occurs in the anterior end [126]. In line with this idea, Takahashi et al. propose a model in which such a gradient of FGF distribution along the embryo axis sets the pace of the somitogenesis front gene expression [127].
Although Fgf1r null allele mutant mice (PSM specific receptor) continue to express Brachyury [128], inactivation of two FGF members, Fgf4 and Fgf8, showed loss of Brachyury expression in the PSM progenitor population [126], confirming the previously described relevance of Fgf8 for establishing the somite boundary [117]. Furthermore, mesodermal cells from Fgf1r loss-of-function embryos, such as in Brachyury deletion mutants, are not able to migrate from the PS and current impaired somite formation [128]. These similarities suggest their relationship and cooperation during this developmental process.
In addition, FGF/Brachyury are also important players in PSM fate choice. Identified as central molecules in the maintenance of PSM mesodermal identity, FGF prevents neuronal differentiation and controls body length by switching to Brachyury expression. Complete body segmentation and end-of-body extension coincides with a less active FGF state and, as a result, lower Brachyury expression, favoring neuronal differentiation rather than conserving mesodermal progenitor potency [118].
WNT- and FGF-signaling pathways are believed to operate both independently and jointly. Analysis of FGF gene deletions in mouse PSM cells contributed to understanding the molecular hierarchy during somite formation. Fgf4 and Fgf8 deletion models showed no expression of WNT pathway genes. Restored WNT expression partially recovered some progenitor marker expression, including Brachyury and Tbx6, whose expression declined due to FGF deletion. This incomplete reestablishment of presomitic expression identity demonstrates FGF activation regardless of WNT signaling. Finally, the study concludes that a deficit in FGF signaling eliminates the progressive characteristic of somitogenesis, by disturbing the activation of Brachyury and Tbx6 at the embryo caudal end, where FGF signaling is usually stronger, resulting in a premature somite differentiation [126].
Despite this demonstration of FGF prevalence over WNT signaling, Stulberg et al. [129] describe a positive regulation between WNT and FGF, calling attention for the importance of the respective pathway balance, since they present opposing effects in many co-regulated genes.
Restored WNT signaling in FGF knockout mice could, as mentioned earlier, to some extent compensate for Brachyury expression. In spite of much evidence of Brachyury's role in maintaining a caudal PSM progenitor nature [118,119,126], the achieved protein level was not able to avoid premature somite differentiation. Therefore, it is possible to conclude that Brachyury might either possess another main role other than maintaining the progenitor population or, as seen in other T-Box protein circumstances, is dose dependent.
WNT-mediated activation of Brachyury transcription was described as having a specific promoter that drives expression in the PS, leading to paraxial mesoderm [121]. Furthermore, axial (notochord) and paraxial mesoderm display two distinct Brachyury promoters [31], meaning that axial mesoderm fate could depend on a different molecule or that besides WNT, another co-activator could be required. However, axial/paraxial differences in Brachyury transcriptional activation do not exclude influences that axial structures could have on somitogenesis.
The residual occurrence of somites in mice Brachyury homozygous knockout phenotype, long ago described by Chesley [113] as being a consequence of a defective notochord, was rejected due to a demonstration of somite formation in absence of the notochord [130]. Nevertheless, an interesting study using chick embryos challenged the accepted idea that axial mesoderm-derived structures do not influence somitogenesis. In fact, the authors observed a delay in the somitogenesis process when notochord and neural tube are absent. Sonic hedgehog (SHH) signals coming from the notochord were shown to influence timed onset of somite formation. fgf8 upregulation was detected in the presence of SHH signals, but this FGF family member could not, at least not alone, rescue normal somite development. This investigation further emphasizes that the somite segmentation delay is only observed in PSM progenitors cells, whereas the anterior third of the PSM is not influenced by notochord signals [131].
In light of that, the hypothesis conceived by Chesley [113], that a defective notochord could result in a phenotype displaying residual somites, could still be proved. Besides the lack of Brachyury, absence of SHH signaling could indirectly change other key molecule expression via FGF downregulation. Regarding PSM's anterior third, notochord impairment might not interfere with residual somite formation, since FGF and Brachyury are usually downregulated in this PSM cellular pool. Moreover, SHH signals seem to have no effect on the rostral PSM. Supporting this concept, a review on PSM rostro-caudal patterning during somitogenesis in different species was addressed by Kimelman and Griffin [132]. They described common caudal dependence on Brachyury ortholog expression and attributed the rostral portion independence to different and specific signaling pathways, even though Brachyury orthologs are expressed in both portions.
Notochord
As previously discussed, along with structures originating from paraxial mesoderm, the central axis mesoderm population has its fate equally disturbed by anomalous Brachyury expression.
As a central player in further embryo development, the notochord is a widely studied axial mesoderm derived-structure known to present important induction activity in its vicinity as in the case of neuroectoderm, the layer located beneath the notochord, which will eventually give rise to the central nervous system [133 –136].
Notochord abnormalities are present in different Brachyury mutants [3,113,114]. Along with Brachyury's important role in signaling, these phenotypes have motivated diverse studies that tried to access Brachyury function in notochord development. Besides mesoderm cells and their epithelial progenitor cells, the notochord is one of the early embryonic structures found to express Brachyury and malformations were associated with Brachyury expression in a dose-dependent manner [108,137]. Chimaera studies revealed an autonomous function of the Brachyury protein in the notochord and in PS cells, but they did not exclude nonautonomous contribution of the gene in the differentiation of other mesoderm-derived tissues [138]. So, regardless of different Brachyury transcription regulatory regions in somites and the notochord (paraxial and axial, respectively), early notochord Brachyury expression is consistent with the previously exposed idea that the notochord may offer somite precursor differentiation signals that are dependent on Brachyury protein expression [31,131]. Still, with respect to the role of the notochord in providing signaling for further cell type specification, Yamada et al. described a significant correlation of neural cell fate with both chick embryo notochord presence and position [136].
In vertebrates, Brachyury expression is generally observed earlier than the appearance of the notochord; however, it is conserved in this structure during its formation [108,139]. Considering the lack of a notochord in mouse knockout phenotypes and expression analysis, early studies in vertebrates regarded Brachyury as indispensable for notochord development [113,140,141] and accounted for the lack in notochord formation as a consequence of embryonic failure to proceed with notochord precursor differentiation.
A variety of studies have demonstrated a positive regulation between FGF signaling and different Brachyury orthologs during notochord formation [119,124,125,142]. In agreement with this observation, chick embryo analysis by Olivera-Martinez [118] reported that gain of FGF activity promotes both Brachyury expression and an axial stem cell state. Conversely, retinoid signaling was found to decrease expression of Brachyury on the notochord, shifting the population status from a mesodermal to a neuronal fate, and providing means for body axis elongation cessation. The authors associate this event to the fact that body axis elongation is disrupted in the Brachyury knockout mouse as originally described by Dobrovolskaïa-Zavadskaïa [2]. Thus, the chick study brings retinoic acid to the intricate network that coordinates correct body axis length, and it finds a correlation between retinoid signaling and decline of FGF/Brachyury activation.
Final remarks on Brachyury
By drawing a parallel between cancer cell environment and somitogenesis, Martin and Kimelman [143] conclude that Wnt signaling, which is maintained in somite formation by Brachyury expression, could act in the same way during both processes supporting cancer growth and paraxial progenitor maintenance during the whole event of somitogenesis.
A recent publication aimed at ascertaining Brachyury targets and revealed that Brachyury is involved in processes such as morphogenesis, cell adhesion, and cell polarity [55]. A ChIP approach assembled expression of several mouse Brachyury target genes, and it showed that 63% of the targets have their expression activated after Brachyury peak expression (E7.5) and that the majority are mesoderm-inducing genes, followed by the ectoderm and endoderm. While illustrating complexity of Brachyury pathways, 13.4% of the promising target genes are transcription factors. The study also suggested a conservation of targets within mammals, excluding other vertebrates. Accordingly, despite T-Box family conservation through evolution, caution must be taken when comparing and inferring conclusions from distinct animal models [55].
Tbx6
Besides Brachyury, Tbx6 was also shown to be decisive in somitogenesis. Tbx6 initial expression occurs in the PS and PSM [144]. Tbx6 knockout in mice gives rise to few and abnormal somites and replaces somitogenesis by development of neural-tube-like structures [145]. In addition, Tbx6 regulates paraxial mesoderm formation and enables the appropriate segmentation process [146].
While providing evidence of a progressive expression pattern change in the course of somitogenesis, Xenopus analysis exhibited a negative regulatory feedback by Tbx6, activating the expression of its repressor in cooperation with co-factors, promoting Tbx6 down-regulation in the anterior somitogenesis front, the point at which segmentation comes to an end [147]. Additional mechanisms of regulating the tbx6 gradient were recently uncovered. Besides gene expression control, the zebrafish tbx6 can be also regulated in a post-transcriptional manner, in which the protein level is decreased by the Ripply protein, a general transcriptional repressor protein, in the anterior PSM [148]. This post-transcriptional regulation of tbx6 is believed to be conserved among species, since Tbx6 protein rather than mRNA was increased in Ripply null mice embryos [127]. In addition, Brachyury protein also had its levels decreased by Ripply, showing that Ripply is a key protein in regulating a somitogenesis boundary, by controlling T-Box protein levels [148].
As pointed out earlier, Tbx6 exhibits an important association with WNT signaling, and a deeper investigation of Tbx6 neural fate inhibition [149] led to a better understanding of WNT-Tbx6 axis during paraxial specification. The study showed high expression levels of Wnt3 in the paraxial mesoderm of Tbx6 null mutant embryos and correlated this to the Wnt3 ability to activate the Sox2 enhancer, in this way promoting neural differentiation. Furthermore, they could demonstrate that unlike Wnt3, Tbx6 showed inhibition of Sox2 enhancer.
Griffin [142] noticed a change in the regulatory order by the eight-cell stage when zebrafish Brachyury homologue (ntl) becomes more influential on other T-Box genes expression, namely Tbx6 and spt (tbx16). The work still points out the common defective paraxial mesoderm formation in both spt and murine Fgf receptor 1 (Fgfr1) knockout mutant phenotypes [150], hypothesizing the existence of an spt murine orthologue located downstream of Fgfr1. Although no mammalian orthologue of spt was found, murine Tbx6 presents high sequence similarity and a comparable expression pattern. Thus, it is certainly possible to associate FGF signaling with T-Box gene regulation during mesodermal differentiation in zebrafish and mice.
Finally, Brachyury and Tbx6 interactions in somitogenesis are not totally elucidated. More information about how they operate, and in which pathways, is still required. Although Brachyury is not essential for Tbx6 expression activation, it might act in its expression maintenance [144]. Moreover, if a more direct interaction is not applied, similar regulatory pathways could be shared, once both present analogous functions and overlapping expression patterns.
Other T-Box Members
Besides the T-Box proteins known to play a role in early embryogenesis, the T-Box family also has other members; a list of them with their mouse counterparts and associated human diseases is found in Figure 3. This section gives succinct information about other T-Box proteins, which are also known to be synthesized in early phases of development.
Two pairs of evolutionarily closely related T-Box proteins, Tbx2/3 and Tbx4/5, are mentioned in the T-Box gene family section with respect to their overlapping expression pattern and their importance in equivalent embryonic development processes.
Although Tbx2 was found to be expressed in an early development process, somitogenesis, Tbx2 null mutant presented no obstacle to segmentation. It is worth noting that Tbx2 expression was detected only in mouse 16–18 somite pair stage, posterior to the Brachyury gene expression that, on the contrary, has considerable effects on somite formation [151].
Tbx3 was found to be the first Tbx2 subfamily member expressed during embryogenesis after the forming of the zygote [62]; however, Tbx4 has the earliest detected expression, when considering even earlier events (Fig. 1). The referred gene is expressed in primordial germ cells and during the whole folliculogenesis up to oocyte maturation, and its role in fertility has been postulated [152,153]. Another function of Tbx4 in development is currently considered, as besides Brachyury, Tbx4 loss of function also impacts allantois development [154]. Tbx4 lineage tracing experiments revealed a noncell autonomous effect of the gene on vasculogenesis during chorioallantoic fusion [155]. In turn, Tbx5 expression analysis showed discrepant results regarding allantois expression in different studies [62,154,156]. Furthermore, Tbx5 is marking the lateral plate mesoderm [157] and is associated with heart development and mutations leading to cardiac and limb malformations [41,44,158]. Differently from the mouse, Tbx5 expression in the chick notochord was described [156]; however, no abnormality of the structure was observed so far.
Footnotes
Acknowledgments
This study was funded by the Deutsche Forschungsgemeinschaft (DFG, K.L. 2544/1-1, S.L. BO1718/4-1), the German Foundation for Heart Research (F/34/11; to A.K. and S.L.), the Forschungskern SyStaR to A.K., BIU (Böhringer Ingelheim Ulm to A.K.), Fortüne program (Faculty of Medicine Tübingen to L.L.), and the Else-Kröner-Fresenius-Stiftung (2011_A200; to A.K. and S.L.). A.K. is indebted to the Baden-Württemberg Stiftung for the financial support of this research project rendered by the Eliteprogramme for Postdocs. A.K. is also an Else-Kröner-Fresenius Memorial Fellow.
Author Disclosure Statement
No competing financial interests exist.