
Abstract
Interleukin (IL)-21, a proinflammatory cytokine, has been developed as an immunotherapeutic approach due to its effects on various lymphocytes, including natural killer (NK) cells and T cells; however, the clinical success in cancer patients has been limited. Recently, mesenchymal stem cells (MSCs) have emerged as vehicles for cancer gene therapy due to their inherent migratory abilities toward tumors. In the present study, we hypothesized that MSCs, genetically modified to express high levels of IL-21 (IL-21/MSCs), can enhance antitumor responses through localized delivery of IL-21. For tumor induction, BALB/c mice were injected intravenously with syngeneic A20 B-cell lymphoma cells to develop a disseminated B-cell lymphoma model. Then, 6 days following tumor induction, the tumor-bearing mice were treated with IL-21/MSCs weekly, four times. Systemic infusion of A20 cells led to hind-leg paralysis as well as severe liver metastasis in the control group. The IL-21/MSC-treated group showed delayed tumor incidence as well as improved survival, whereas the MSC- and recombinant adenovirus-expressing IL-21 (rAD/IL-21)-treated groups did not show significant differences from the untreated mice. These therapeutic effects were associated with high levels of IL-21 delivered to the liver, which prevented the formation of tumor nodules. Furthermore, the infusion of IL-21/MSCs led to induction of effector T and NK cells, while potently inhibiting immune suppressor cells. Our findings demonstrate that IL-21-expressing MSCs have the therapeutic potential to induce potent antitumor effects against disseminated B-cell lymphoma through localized IL-21 delivery and induction of systemic antitumor immunity.
Introduction
I
Based on these results, IL-21 has been evaluated in phase I and II trials targeting metastatic melanoma [10 –14], renal cell carcinoma [12,13,15], and non-Hodgkin's lymphoma [16]. However, the overall efficacy of IL-21 in most clinical trials to date has been disappointing [17]. While IL-21 has been shown to be considerably less toxic than the related cytokine IL-2, its clinical efficacy has been insignificant. The clinical outcome of most metastatic melanoma and renal cell carcinoma cases treated with IL-21 as a single agent showed progressive or stable disease [10 –14]. Studies have also evaluated the potential of combination strategies, such as the use of sorafenib or rituximab, to improve response rates [15,16]. Although some clinical trials have indicated a modest additive effect and durable responses in a small subset of patients, there is still insufficient evidence as to whether the addition of IL-21 contributed to the observed outcomes. Thus, while IL-21 is a unique immunotherapeutic agent with an encouraging antitumor mechanism of action, it is also becoming evident that durable responses occur only in a minority of treated patients, indicating a strong rationale for improving IL-21-based therapeutic approaches.
Overall, the low response rates of IL-21 treatment may be attributed to the limiting features of most biological agents, including restricted delivery and low site specificity of the agent administered. Recently, mesenchymal stem cells (MSCs), defined as self-renewing multipotent progenitor cells with multilineage potential, have attracted interest in various clinical applications due to their unique properties, including their migratory capacity [18]. Such migratory potential has encouraged the development of cell-based gene therapy using MSCs as a delivery vehicle to deliver various therapeutic proteins to target sites. Although the exact mechanism remains to be determined, it has been suggested that MSC migration is dependent on various chemokine and receptor interactions between the molecules constitutively produced in a tumor microenvironment and the corresponding receptor pairs present on MSCs. Among the chemokine receptor pairs, stromal cell-derived factor 1 (SDF-1) and C-X-C chemokine receptor type 4 (CXCR4) are important mediators of stem cell recruitment to tumors [19]. Furthermore, in a recent study, in vivo bioluminescent imaging of systemically administered MSCs demonstrated direct evidence of MSC tropism to tumors [20]. Luciferase-labeled MSCs displayed colocalization and engraftment at tumor sites in both breast carcinoma and ovarian tumor models. Based on these observations, MSCs have been engineered to produce anticancer genes, including proinflammatory cytokines [21 –25], proapoptotic proteins [26,27], and antiangiogenic agents [28], for the development of MSC-based gene therapy that may overcome the limitations of previous biological drugs.
In the present study, we produced MSCs genetically modified to express high levels of IL-21 (IL-21/MSCs). We hypothesized that IL-21/MSCs could maximize the antitumor effects of IL-21 against disseminated lymphoma through localized delivery. Our results demonstrate that IL-21/MSC treatment inhibits tumor cell proliferation at various sites of tumor dissemination, which was associated with efficient delivery of IL-21. Furthermore, the presence of high levels of IL-21 promoted redirection of the host immune system against tumor cells by recruiting effector cells while inhibiting immunosuppressive cells at the tumor site. Thus, we propose IL-21/MSC treatment as an improved IL-21-based immunotherapy and a novel therapeutic approach to target disseminated B-cell lymphoma.
Materials and Methods
Cell lines and treatment
A20 cells were obtained from the American Type Culture Collection and cultured in RPMI 1640 (Gibson) containing 2 mM
A20 cells (1 × 104/well) were treated with recombinant mouse IL-21 (rIL-21; 100, 200, 500 ng/mL) purchased from R&D Systems or cell culture supernatant in 96-well plates for 96 h. Cell growth was assessed using the Cell Counting Kit-8 (CCK-8) assay (Dojindo) according to the manufacturer's protocol.
Mice
Eight-week-old female BALB/c mice were purchased from OrientBio. The mice were maintained under specific pathogen-free conditions in an animal facility with controlled humidity (55% ± 5%), light (12-h light/12-h dark), and temperature (22°C ± 1°C). The air in the facility was passed through a HEPA filter system designed to exclude bacteria and viruses. Animals were fed mouse chow and tap water ad libitum. The protocols used in the present study were approved by the Animal Care and Use Committee of The Catholic University of Korea.
Isolation and culture of MSCs
Bone marrow cells were collected from BALB/c mice by flushing femurs and tibias with Dulbecco's modified Eagle's medium (Gibco) containing 2 mM
Characterization of MSCs
For phenotypical analysis, MSCs were stained for surface markers Sca-1 phycoerythrin (PE) (Ly-6A/E), CD29-FITC, c-kit-FITC (CD117), CD34-PE (all from BioLegend), CD11b-allophycocyanine (APC), CD90-FITC, CD73-PE-Cy7, CD45-PE, CD105-PE, and CD44-fluorescein isothiocyanate (FITC) (all from eBioscience). Before surface staining, MSCs were Fc-blocked with CD16/CD32 (BD Pharmingen), as described previously [30].
For differentiation assays, MSCs were seeded into 24-well plates at a density of 1 × 104 per well and then induced to differentiate into adipocytes, osteoblasts, or chondrocytes using the Mouse MSC Functional Identification Kit (R&D Systems) following the manufacturer's protocol. The medium was changed three times a week for 14–21 days. Adipocytes, osteoblasts, and chondrocytes were detected using goat anti-mouse FABP4 polyclonal antibody, sheep anti-mouse osteopontin polyclonal antibody, and sheep anti-mouse collagen II antigen affinity-purified polyclonal antibody, respectively.
Adenoviral expression vectors for IL-21 and transduction conditions
Adenoviruses expressing enhanced green fluorescent protein (eGFP) and IL-21 were generated using the AdEasy system (Seolin Bioscience Co. Ltd.). The open reading frame of IL-21 was subcloned into a pShuttle-CMV transfer vector expressing eGFP. The resulting plasmids were linearized with PmeI and cotransformed with the pAdEasy into Escherichia coli BJ5183 cells. Recombinants were selected for kanamycin resistance, and recombination was confirmed by restriction analysis and sequencing. High-titer stocks were generated by transfecting the linearized recombinant plasmid with PacI into 293 cells. Primary supernatants containing recombinant adenoviruses were plaque purified and amplified in 293 cells. MSCs were generated as described above and transduced with adenoviral IL-21 at multiplicities of infection of 100 in the presence of 8 μg/mL polybrene. The expression of eGFP was analyzed by flow cytometry, and IL-21 expression was confirmed by real-time quantitative polymerase chain reaction (qPCR) and enzyme-linked immunosorbent assay (ELISA).
Induction and treatment of a disseminated lymphoma model
Eight-week-old female BALB/c mice were injected intravenously with 1 × 106 A20 lymphoma B cells resuspended in 200 μL phosphate-buffered saline (PBS) to induce a disseminated lymphoma model. Six days after tumor inoculation, mice were divided randomly into four groups and injected intravenously with PBS (n = 10), 2 × 107 plaque-forming units (pfu) of recombinant adenoviral IL-21 (rAd/IL-21, n = 10), 1 × 105 MSCs (n = 10), or 1 × 105 IL-21/MSCs (n = 10) on days 6, 13, 20, and 27. Mice were monitored daily for the presence of visible tumors or hind-leg paralysis. Mice were sacrificed if signs of hind-leg paralysis were observed.
Tumor histopathology
Mice were sacrificed 41 days after the A20 cell challenge. Livers were removed, weighed, and assessed for the number of liver metastases. Liver sections were then fixed in 10% buffered formalin (Sigma-Aldrich) and stained with hematoxylin (Sigma-Aldrich) and eosin Y 1% solution (Muto PureChemical Co., Ltd.) for histological examination.
For immunohistochemical analysis, tissues were incubated first with the primary rabbit anti-mouse Ki67 and programmed-death ligand-1 (PDL-1; Abcam) antibodies overnight at 4°C and a horseradish peroxidase-conjugated secondary antibody for 1 h. The final color product was developed using diaminobenzidine (Sigma-Aldrich). Sections were counterstained with hematoxylin, and samples were photographed using an up-light photomicroscope (Leica).
For MSC trafficking, to replicate the same tumor model used throughout the study, we first inoculated the mice intravenously with 1 × 106 A20 tumor cells on day 0. IL-21/MSCs were then labeled with PKH-26 (Sigma-Aldrich) as per the manufacturer's protocol. Then, 6 days after tumor inoculation, PKH-26-labeled IL-21/MSCs were injected into tumor-bearing cells and mice were sacrificed at different time points following administration. Liver and spleen tissues were excised, snap-frozen in liquid nitrogen, counterstained with 4′,6-diamidino-2-phenylindole (DAPI), and photographed using an inverted fluorescent microscope (Carl Zeiss).
Flow cytometric analysis
Mononuclear cells from spleens were immunostained with various combinations of the following fluorescence-conjugated antibodies: CD360-biotin (IL-21R), streptavidin-PerCP, CD4-PerCP, CD8-FITC, IFN-γ APC, CD3-PerCP, DX5-APC (CD49b), CD25-APC, Foxp3-PE, IL-10-APC (all from eBioScience), and Gr-1-PerCP (Ly-6G/C; BioLegend). Before intracellular cytokine staining, cells were stimulated in the culture medium containing phorbol myristate acetate (25 ng/mL; Sigma-Aldrich), ionomycin (250 ng/mL; Sigma-Aldrich), and monensin (GolgiStop, 1 μL/mL; BD Pharmingen) in an incubator with 5% CO2 at 37°C for 4 h. Intracellular staining was performed using an intracellular staining kit (eBioscience) according to the manufacturer's protocol. Flow cytometry was performed on a FACSCalibur flow cytometer (BD Pharmingen) using the FlowJo software (TreeStar).
Annexin V/PI assays for apoptosis
A20 cells (2 × 105/well) were treated with rIL-21 (100, 200, 500 ng/mL) or cell culture supernatant in 12-well plates for 96 h. Cells were stained with annexin V-FITC and propidium iodide (PI) and evaluated for apoptosis by flow cytometry according to the manufacturer's protocol (BioVision). Briefly, 1 × 106 cells were washed twice with PBS and stained with 5 μL annexin V-FITC and 5 μL PI in 1× binding buffer for 5 min at room temperature in the dark. The apoptotic cells were determined using a FACSCalibur flow cytometer. Early apoptotic (annexin V positive, PI negative) and late apoptotic (annexin V positive and PI positive) cells were included in the analysis.
Real-time quantitative PCR
Total RNA was extracted from splenocytes using the TRIzol reagent (Invitrogen). Total RNA (2 μg) was reverse transcribed at 50°C for 2 min, followed by 60°C for 30 min. Quantitative PCR was performed using the IQ SYBR Green Super Mix and CFX-96 real-time system (Bio-Rad) under the following conditions: denaturation at 95°C for 3 min and annealing for 30 s at 58°C followed by 30 s of extension at 72°C. For quantification, relative mRNA expression of specific genes was obtained using the 2−ΔΔCt method and the β-actin housekeeping gene for normalization. The following gene-specific primers were used: β-actin (forward: 5′-GAA ATC GTG CGT GAC ATCAAA G-3′; reverse: 5′-TGT AGT TTC ATG GAT GCC ACA G-3′), IL-21 (forward: 5′-CGC AAG ATT CCT GAG GAT CCG AGA AG-3′; reverse: 5′-GCA TTC GTG AGC GTC TAT AGT GTC-3′), tumor necrosis factor (TNF)-α (forward: 5′-GCC TCT TCT CAT TCC TGC TTG-3′; reverse: 5′-CTG ATG AGA GGG AGG CCA TT-3′), interferon (IFN)-α (forward: 5′-GGA CTT TGG ATT CCC GCA GGA GAA G-3′; reverse: 5′-GCT GCA TCA GAC AGC CTT GCA GGT C-3′), IFN-β (forward: 5′-AAC CTC ACC TAC AGG GCG GAC TTC A-3′; reverse: 5′-CCC ACG TCA ATC TTT CCT CTT GCT TT-3′), IFN-γ (forward: 5′-CGG CAC AGT CAT TGA AAG CCT A-3′; reverse: 5′-GTT GCT GAT GGC CTG ATT GTC-3′), granzyme B (forward: 5′-TGTCTCTGGCCTCCAGGACAA-3′; reverse: 5′-CTCAGGCTGCTGATCCTTGATCGA-3′), perforin (forward: 5′-GTACAACTTTAATAGCGACACAGTA-3′; reverse: 5′-AGTCAAGGTGGAGTGGAGGT-3′), forkhead box P3 (Foxp3) (forward: 5′-GGC CCT TCT CCA GGA CAG-3′; reverse: 5′-GCT GAT CAT GGC TGG GTT GT-3′), transforming growth factor (TGF)-β (forward: 5′-CCC TAT ATT TGG AGC CTG GA-3′; reverse: 5′-GTT GGT TGT AGA GGG CAA GG-3′), IL-10 (forward: 5′-GTG AAG ACT TTC TTT CAA ACA AAG-3′; reverse: 5′-CTG CTC CAC TGC CTT GCT CTT ATT-3′), and indoleamine 2,3-dioxygenase (IDO) (forward: 5′-CGG ACT GAG AGG ACA CAG GTT AC-3′; reverse: 5′-ACA CAT ACG CCA TGG TGA TGT AC-3′).
ELISA for IL-21
IL-21 concentrations were measured by sandwich ELISA from culture supernatants and liver homogenates. Liver was homogenized in a homogenizer buffer (50 mM Tris-HCl, pH 7.4, 250 mM NaCl, 5 mM EDTA) plus protease inhibitors (Roche). The homogenate was centrifuged (11,000 g, 15 min). The resulting supernatants were collected and measured by sandwich ELISA as follows. Briefly, anti-IL-21 (R&D Systems) was added to a 96-well plate and incubated overnight at 4°C. The wells were blocked with blocking solution (PBS containing 1% bovine serum albumin) for 2 h at room temperature. The test samples and standard recombinant IL-21 were added to separate wells of the 96-well plate, and the plate was then incubated at room temperature for 2 h. The plate was washed, biotinylated IL-21 polyclonal antibody (R&D Systems) was added, and the reaction was allowed to proceed for 2 h at room temperature. The plate was washed, 2,000-fold diluted ExtrAvidin–alkaline phosphatase (Sigma-Aldrich) was added, and the reaction was allowed to proceed for a further 2 h. The plate was then washed, and 50 μL of p-nitrophenyl phosphate disodium salt (Pierce Chemical Company) diluted to 1 mg/mL in diethanolamine buffer (Sigma-Aldrich) was applied. Absorbance was measured at 405 nm on an ELISA microplate reader (Molecular Devices).
Statistical analysis
All statistical tests were performed using the SPSS software (ver. 10.0 for Windows; SPSS, Inc.). Tests were two sided, and values of P < 0.05 were considered to indicate statistical significance. Comparisons between groups were analyzed using the Kruskal–Wallis test. Pair-wise group comparisons were performed using the Mann–Whitney U test, and P values were adjusted for multiple comparisons using Bonferroni's method to determine the statistical significance of the comparisons. The experimental values are expressed as the mean ± standard error of the mean.
Results
Characterization and IL-21 gene transduction of MSCs
To develop a cell-based delivery system for IL-21, we first culture expanded MSCs from BALB/c mouse bone marrow through repeated passaging based on plastic adherence. MSCs demonstrated a typical spindle-shaped cellular morphology and differential potential toward adipocytes, osteoblasts, and chondrocytes under in vitro culture conditions (Fig. 1A). For phenotypic characterization, surface protein expression at passage 10 was examined by flow cytometry. The cells were positive for Sca-1, CD44, and CD29 but negative for c-kit, CD11b, and CD34 (Fig. 1B). MSCs were then transduced with recombinant adenovirus expressing IL-21 (rAD/IL-21) and eGFP. To confirm that genetically modified IL-21/MSCs successfully expressed IL-21, we first used flow cytometry to determine eGFP expression. On day 3 following transduction, eGFP expression could be visualized under the fluorescence microscopy, and the transduction efficiency, as determined by the percentage of eGFP-positive cells using flow cytometry, was 87.0% relative to nontransduced MSCs (Fig. 1C). We also examined IL-21 expression using real-time PCR (Fig. 1D) and ELISA (Fig. 1E). IL-21/MSCs showed elevated production of IL-21, by at least a 20,000-fold increase in mRNA level and a 200-fold increase in secreted IL-21 protein level, compared with nontransduced MSCs. Furthermore, we investigated any phenotypic changes in the genetically modified IL-21/MSCs; however, there was no difference in the phenotype or chemokine receptor expression compared with nontransduced MSCs (data not shown).
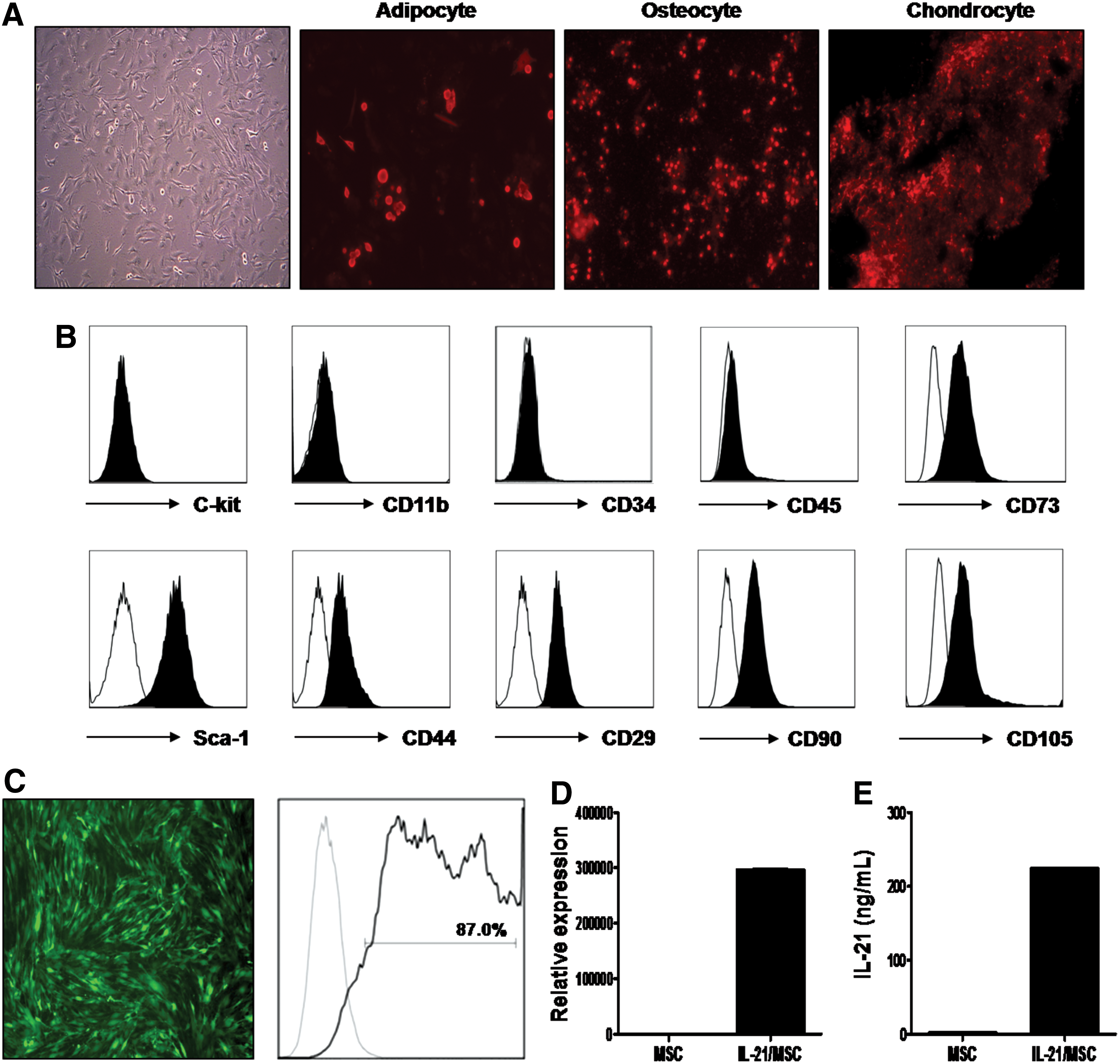
BALB/c-derived mesenchymal stem cells (MSCs) were genetically modified to express high levels of murine interleukin (IL)-21.
IL-21 contributes to lymphoma cell death induced by the MSC culture-conditioned medium in vitro. To investigate the biological effects of IL-21 in A20 cells, we first confirmed the expression of the IL-21R in murine A20 B-cell lymphoma cells (Fig. 2A). Next, we treated the cells with different concentrations of murine recombinant IL-21 protein (rIL-21). In addition, to further determine whether IL-21 secreted from IL-21/MSCs could demonstrate comparable effects to those of rIL-21, we collected the supernatant from nontransduced MSCs and IL-21/MSCs 3 days after transduction and treated A20 lymphoma cells with the collected supernatant. As a control, the complete culture medium was added to the A20 cells. We observed that the cells treated with supernatants from both MSCs and IL-21/MSCs showed significantly reduced A20 cell growth (P < 0.05, Fig. 2B), whereas rIL-21 alone induced moderate cell death. Furthermore, the culture supernatant of IL-21/MSCs induced more cell death than did rIL-21 (P < 0.05), suggesting that additional cytokines produced by MSCs may also be involved. Next, we evaluated apoptosis of A20 cells by flow cytometry using annexin V and PI staining. We observed that the treatment of rIL-21 induced both early and late apoptosis in A20 cells in a dose-dependent manner (P < 0.05). However, the culture supernatant from both nontransduced MSCs and IL-21/MSCs induced significant late apoptotic cell death (88.4% ± 0.4% and 78.4% ± 0.59%, respectively; P < 0.05; Fig. 2C), indicating that while IL-21 itself does not play a critical role in inducing lymphoma cell death, IL-21 plays a contributing role.
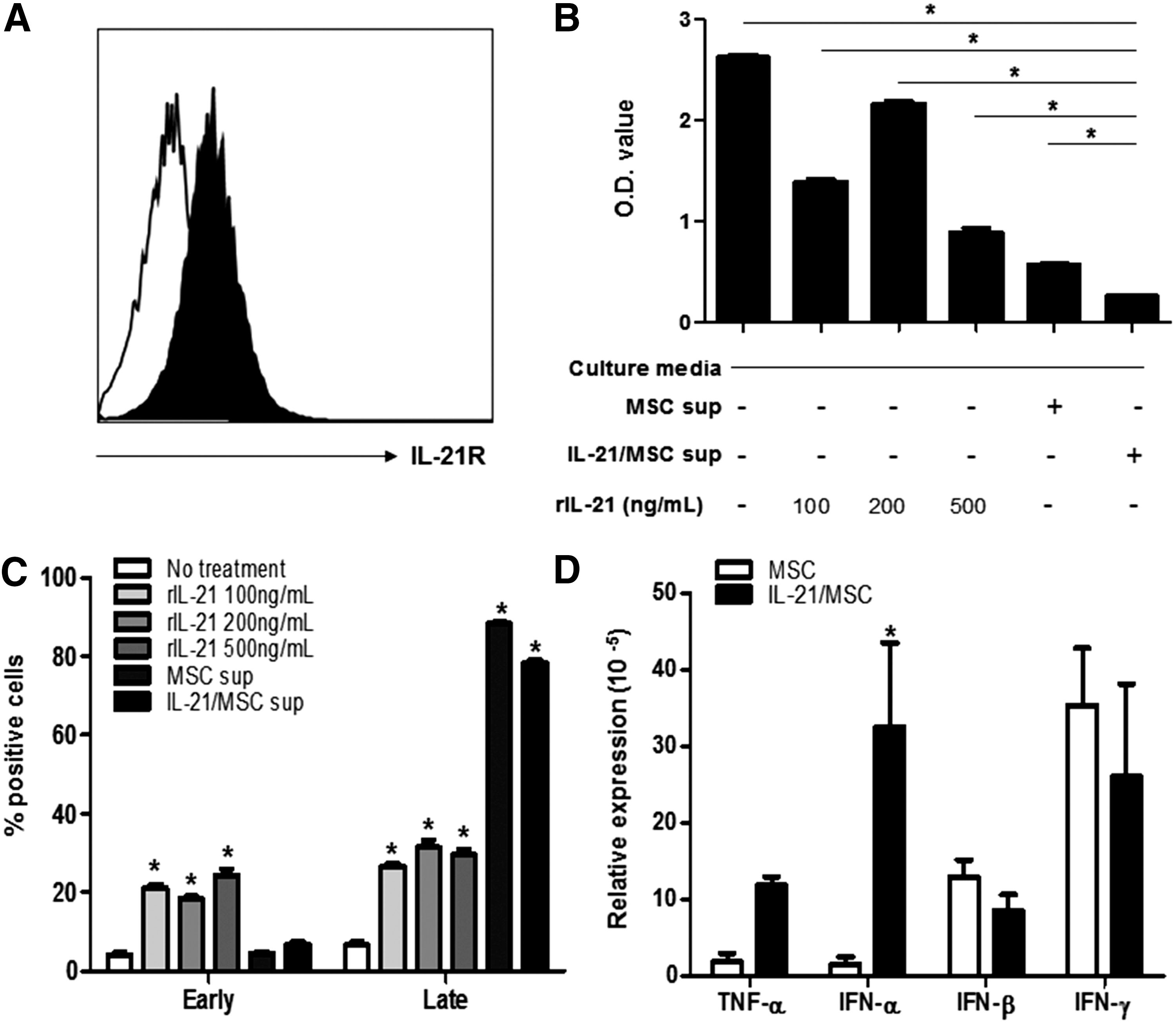
IL-21 induces cell death in lymphoma cells in vitro.
Because significant cell death was induced by the MSC culture supernatants, we examined the expression levels of other cytokines produced by MSCs that may potentially be involved in apoptosis, including TNF-α, IFN-α, IFN-β, and IFN-γ, by quantitative real-time PCR. MSCs showed expression of IFN-β and IFN-γ. While IL-21/MSCs showed similar levels of IFN-β and IFN-γ, there was a substantial increase in IFN-α, by 20-fold, compared with nontransduced MSCs (P < 0.05). Thus, we hypothesized that IL-21/MSCs could not only effectively deliver IL-21 but also directly induced cell death at a tumor site.
Therapeutic effects of IL-21/MSCs in prevention of disseminated B-cell lymphoma growth and liver metastasis
We developed a disseminated B-cell lymphoma model in mice to mimic the systemic disease commonly present in human lymphoma patients. First, we inoculated the mice intravenously with 1 × 106 A20 murine B-cell lymphoma cells through the tail vein. Six days after tumor inoculation, mice were treated intravenously with PBS, 2 × 107 pfu of rAd/IL-21, 1 × 105 MSCs, or 1 × 105 IL-21/MSCs. To determine the tumor-free survival, mice were monitored for development of hind-leg paralysis and any visible tumor manifestations (Fig. 3). The control group that received PBS showed a tumor incidence rate of 100%, in which all mice developed hind-leg paralysis and tumor outgrowths, indicating the lethality of the disease. Mice treated with rAd/IL-21 or MSCs did not show improved tumor-free survival, and all mice developed tumors by day 40. In contrast, IL-21/MSC-treated mice showed delayed clinical tumor growth, and three mice remained tumor-free for more than 80 days.
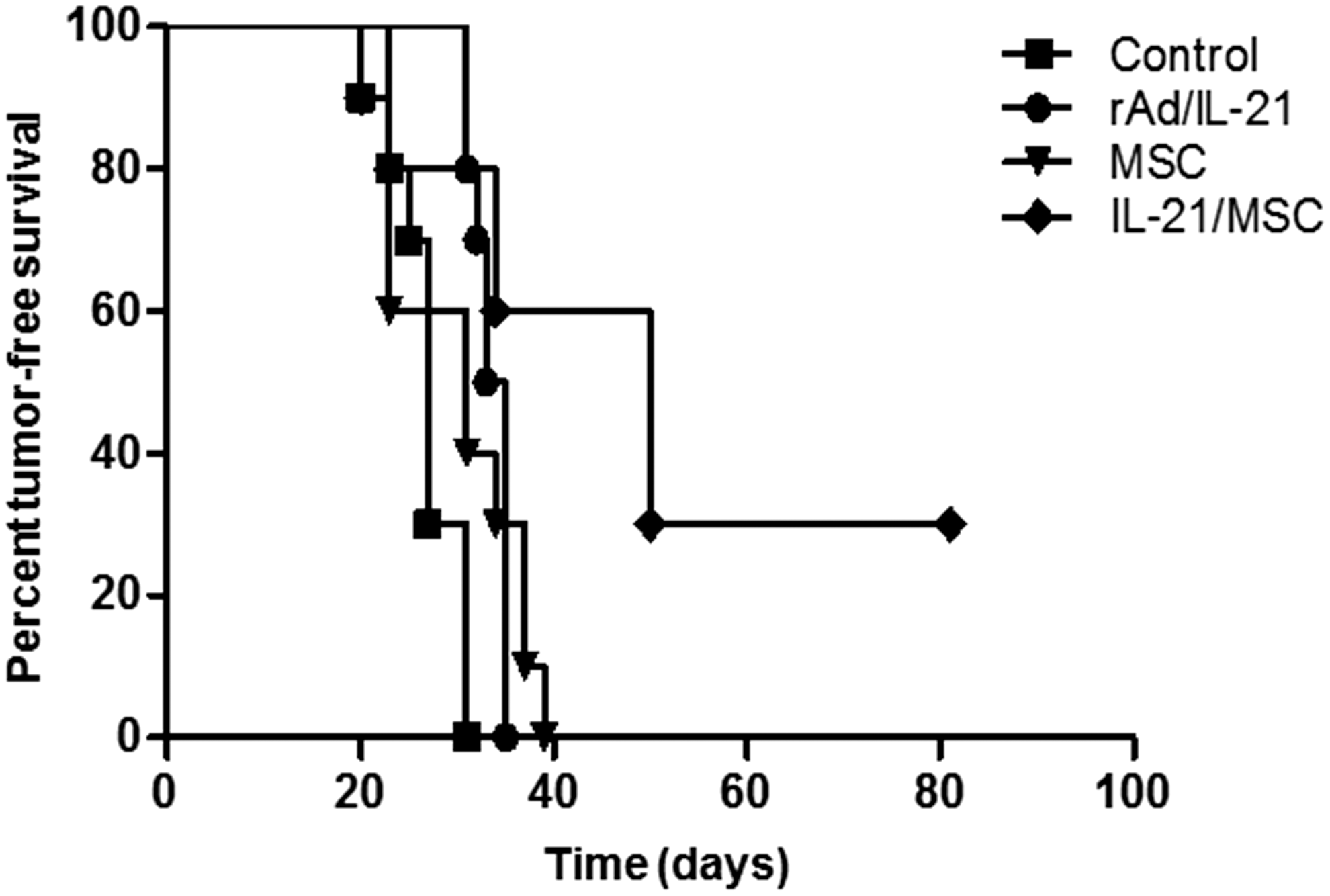
Improved tumor-free survival induced by IL-21/MSCs in a disseminated B-cell lymphoma model. At 1 week following intravenous injection of A20 lymphoma cells, mice were treated with phosphate-buffered saline (PBS) (control, n = 10), 2 × 107 plaque-forming units of recombinant adenoviral interleukin-21 (rAd/IL-21, n = 10), 1 × 105 MSCs (n = 10), or 1 × 105 IL-21/MSCs (n = 10) for 4 weeks at 1-week intervals. Mice were monitored daily for the presence of visible tumors or hind-leg paralysis. Significantly improved tumor-free survival was seen in tumor-bearing mice treated with IL-21/MSCs.
On day 41 after tumor inoculation, 2 weeks after the last treatment, mice from each group were sacrificed. Tumor-bearing mice showed tumor outgrowths in various tissues, including liver, spleen, axillary lymph nodes, mammary glands, and ovaries (data not shown). Liver tissues were removed and assessed for weight and number of metastases. We observed enlarged livers with tumor nodules in the PBS-treated control group (13.2 ± 0.8). The livers of IL-21/MSC-treated mice resembled those of normal mice; there was an absence of visible tumor nodules (P < 0.05, compared with the control and MSC-treated group, and P < 0.01, compared with rAD/IL-21-treated group; Fig. 4A–C). In addition, histological examination of the liver sections displayed a lymphoblastic morphology in the PBS-treated control mice. The liver sections of rAd/IL-21 and MSC-treated mice showed fewer tumor cells and signs of necrosis compared with PBS-treated mice. However, IL-21/MSC-treated mice exhibited a complete absence of tumors in the liver (Fig. 4D). Similarly, immunohistochemical analysis demonstrated significantly reduced cell proliferation in the IL-21/MSC-treated group compared with the control group (Fig. 4E). These results suggest that IL-21/MSC treatment effectively prevents the formation of tumor nodules in the liver.

Effects of systemically administered IL-21/MSCs on inhibition of liver metastases in vivo. Following 4 weekly intravenous administrations of PBS (control), rAd/IL-21, MSCs, or IL-21/MSCs, mice were sacrificed on day 41 after tumor cell inoculation. IL-21/MSC-treated mice showed reduced numbers of tumor nodules present in the liver
Therapeutic effect of IL-21/MSCs is associated with local delivery of IL-21
To determine whether IL-21/MSCs could deliver IL-21 locally to a tumor site, we assessed the concentrations of IL-21 in the liver using liver homogenate samples collected on day 41 after tumor inoculation. Results indicated elevated levels of IL-21 in the IL-21/MSC-treated mice (738 ± 112 pg/mL), while the IL-21 concentration was reduced in the control group (408 ± 29.5 pg/mL; P < 0.05; Fig. 5A). Furthermore, we examined the expression of IL-21 in the spleen using qPCR. IL-21 expression was increased significantly in the spleen (P < 0.05; Fig. 5B). These results provide indirect evidence of MSCs homing to tumor sites, including the liver and spleen. To directly track homing of MSCs to tumor sites, we labeled IL-21/MSCs with PKH26 red fluorescent dye and administered them to lymphoma-bearing mice. Animals were sacrificed, and spleen and liver tissues were harvested at different time points following the administration of MSCs. PKH-26-labeled IL-21/MSCs were not noticeable 24 h following intravenous infusion. However, the presence of IL-21/MSCs in both liver and spleens of tumor-bearing mice became evident 1 week after infusion.
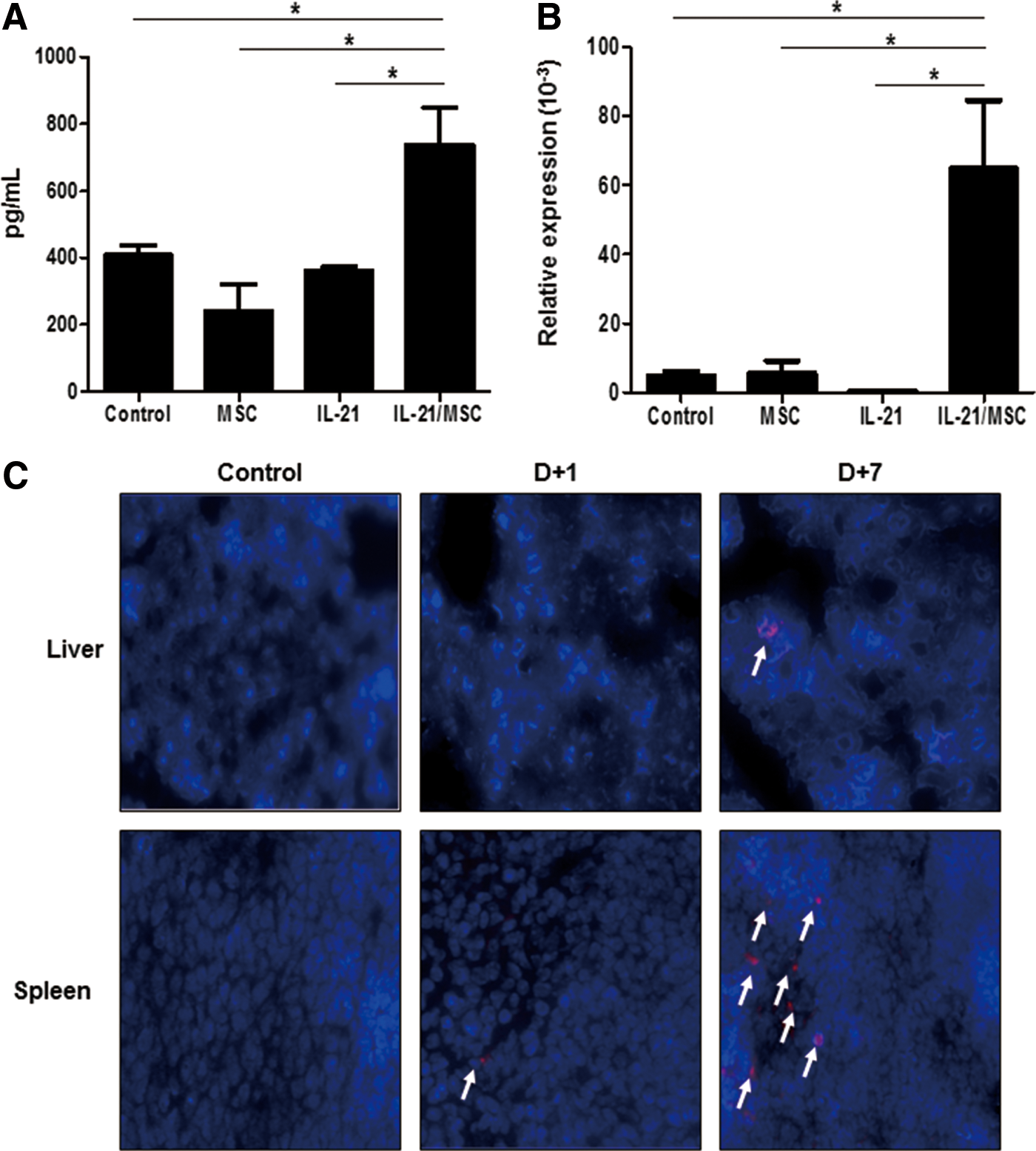
Therapeutic effects of IL-21/MSCs are associated with the local delivery of IL-21. IL-21/MSCs exhibited efficient delivery of IL-21 to tumor sites, as determined by
Effects of IL-21/MSC treatment on tumor immunity
IL-21 plays an important role in enhancing antitumor immunity through inducing effector cells, such as CD8+ T, NK, and NKT cells. We first investigated the effects of IL-21/MSCs in inducing effector immune responses in the spleen compared with MSC or rAd/IL-21 treatment alone. We observed higher levels of both IFN-γ-expressing CD4+ (2.26%, P < 0.05) and CD8+ T cells (1.86% ± 0.1%) in the IL-21/MSC-treated group (Fig. 6A, B). Furthermore, CD3-DX5+ NK cells were increased significantly following treatment with IL-21/MSCs (4.4% ± 0.04%; P < 0.05; Fig. 6A, C). In contrast, MSC- and rAD/IL-21-treated mice showed no significant change in immune cell subsets compared with untreated control mice. Granzyme B (Fig. 6D) and perforin (P < 0.05, compared with the rAd/IL-21-treated group; Fig. 6E) showed elevated levels in the splenocytes of the IL-21/MSC-treated group, suggesting functional maturation of T or NK cells, in addition to their induction.
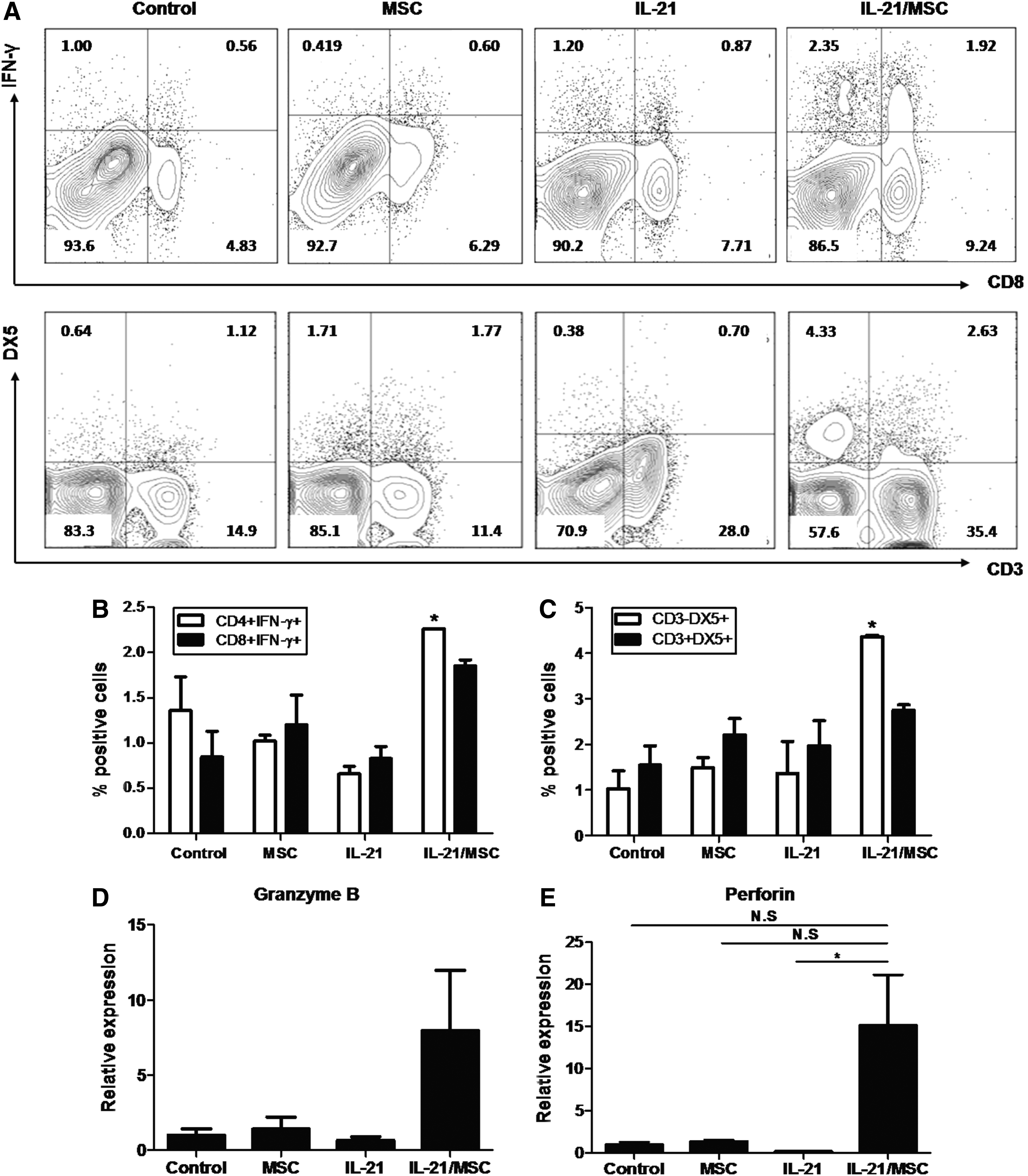
Treatment with IL-21/MSCs results in enhanced antitumor immunity.
In addition to enhancing antitumor immunity, regulation of the tumor immunosuppressive microenvironment plays a key role in controlling tumor growth. Our results showed that following IL-21/MSC treatment, CD11b+ Gr-1+ myeloid-derived suppressor cells (MDSCs) were significantly reduced in the spleen (0.216%) compared with the untreated control group (3.2% ± 0.91%; P < 0.05). However, the changes in the levels of regulatory T cells (Tregs) and IL-10-expressing CD4+ T cells were not statistically significant (Fig. 7A). Furthermore, we assessed the changes in mRNA levels of immunosuppressive molecules of the spleen, including Foxp3, TGF-β, IL-10, and IDO, associated with immune-suppressing cells (Fig. 7B). While the Foxp3 level was increased slightly in the untreated tumor-bearing mice, there was no significant difference between the treatment groups. While there was no statistical significance, we observed reduced levels of TGF-β, IL-10, and IDO in the rAd/IL-21- and IL-21/MSC-treated groups, compared with the control group, whereas the levels were augmented in the MSC-treated group. Additionally, we evaluated the expression of PDL-1 in the liver by immunohistochemistry (Fig. 7C). Untreated tumor-bearing mice showed high expression of PDL-1 directly in the tumor and hepatocytes. In the MSC- and rAd/IL-21-treated groups, there was reduced expression of PDL-1 in the tumor; however, we observed accumulation of PDL-1-expressing immune cells. In contrast, in the IL-21/MSC-treated group, the levels of both PDL-1-expressing tumor cells and immune cells were diminished markedly. These findings suggest that the therapeutic effects of IL-21/MSCs were associated with regulation of antitumor immune responses by inducing effector cells while inhibiting immune-suppressing cells at the tumor site.
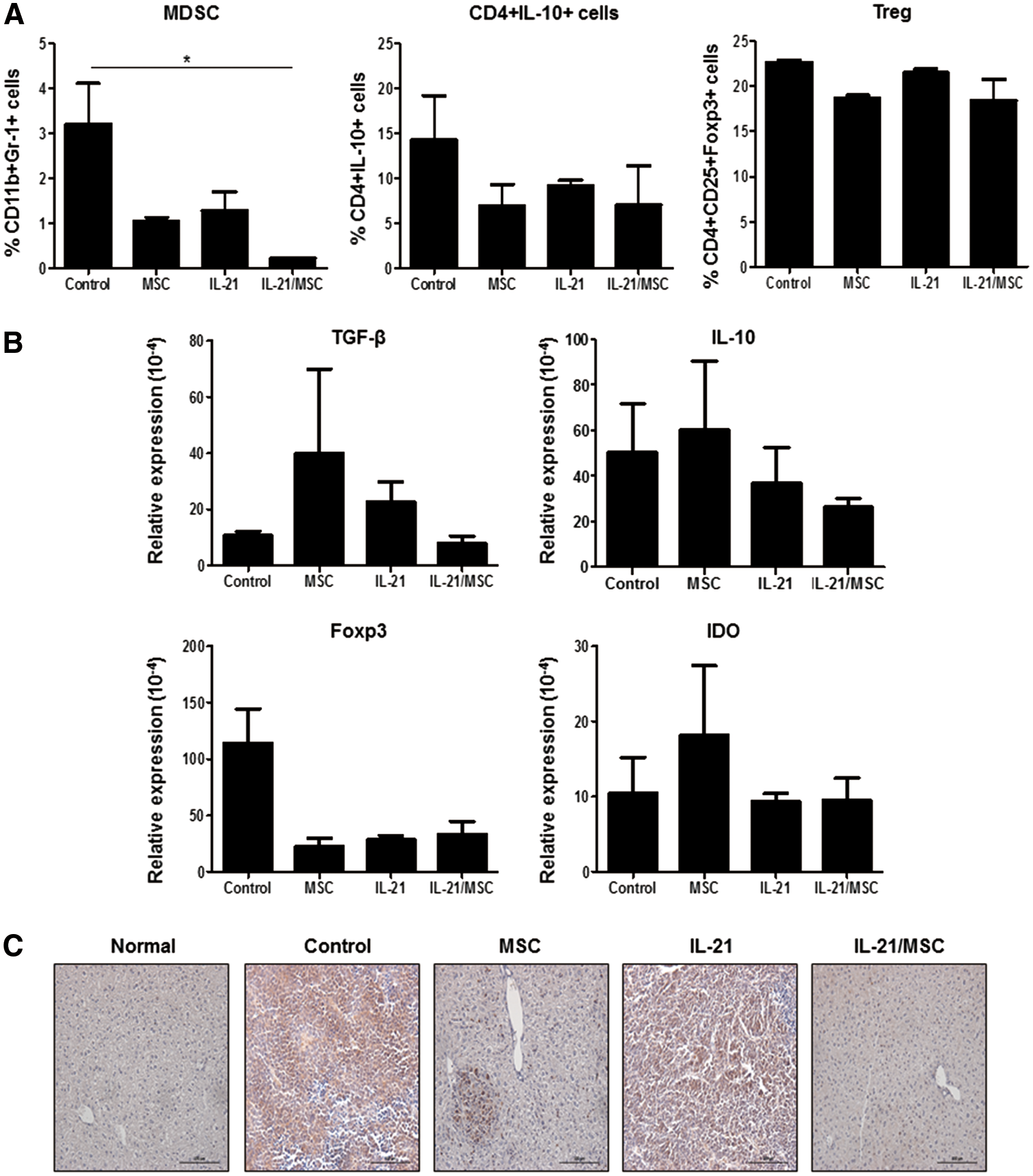
Treatment of IL-21/MSCs reduced the immunosuppressive environment.
Discussion
In this study, we demonstrated that bone marrow-derived MSCs, genetically modified to express IL-21, not only induced cell death in lymphomas in vitro but also elicited therapeutic effects in vivo in a lethal murine lymphoma model by inhibiting tumor cell proliferation, preventing tumor metastasis, and inducing antitumor immunity. Our results suggest that IL-21/MSCs maximize the antitumor effects of IL-21 therapy through (1) direct effects of IL-21 on B-cell lymphoma cells, (2) local delivery of IL-21 to tumor sites in vivo, and (3) immune modulation of the tumor environment.
Previously, clinical evaluations of IL-21 focused mainly on melanoma and renal cell carcinoma, in which a related cytokine, IL-2, failed to show therapeutic benefits. The use of IL-21 to treat lymphoid malignancies emerged due to the expression of IL-21R on lymphocytes, but not on tumor cells of carcinomas and melanomas [31]. Following the first report of the direct apoptotic effects of IL-21 on chronic lymphocytic leukemia cells [32], the effects of IL-21 have been examined, especially in B-cell neoplasms, including follicular lymphoma [33,34], chronic lymphocytic leukemia [32,35], and diffuse large B-cell lymphoma [36]. In these studies, it was noted that majority of primary lymphoma cell samples expressed IL-21R and were sensitive to IL-21 treatment. The direct antitumor effects of IL-21 on these malignancies were associated with the induction of apoptosis through the activation of caspases-3, -8, and -9 [33], reduction in the antiapoptotic protein BCl-2 [36,37], and upregulation of the proapoptotic protein Bim [35] through the JAK/STAT pathway. However, the effects of IL-21 differ depending on the neoplastic cell type. In contrast to B-cell neoplasms, IL-21 showed proliferative effects in adult T-cell lymphoma [38], cutaneous T-cell lymphoma [39], and anaplastic large-cell lymphoma cells [40]. While the exact mechanisms remain unclear, the promotion of cell survival in most T-cell-derived lymphoma subtypes is mediated by the JAK3/STAT3 pathway, which induces proliferation and protects cells from apoptosis. Thus, it is suggested that IL-21 may provide a better therapeutic approach for B-cell-derived malignancies versus other lymphoid neoplasms. In the present study, we observed the expression of IL-21R on the surface of murine B-cell lymphoma A20 cells and the treatment of recombinant IL-21 protein inhibited cell growth moderately and induced apoptosis in vitro. Further studies are needed to directly compare the effects of IL-21 in different neoplastic cell types and address the signaling pathways involved.
Despite encouraging in vitro results suggesting potent antitumor effects of IL-21 against lymphomas, the clinical application of IL-21 in lymphoma patients has been limited. Recently, the administration of recombinant IL-21 in combination with the anti-CD20 monoclonal antibody rituximab has been investigated for the treatment of low-grade B-cell lymphoproliferative malignancies. The safety of recombinant IL-21 was observed; however, only 3 of 19 patients showed complete responses to the combined treatment [16]. Furthermore, there is no clinical study reporting the use of IL-21 as a monotherapy in lymphoma. It has been suggested in clinical studies of metastatic melanoma and renal cell carcinoma patients that IL-21 as a single agent is insufficient. Like most biological agents, we believe that low efficacy of IL-21 in vivo may be associated with restricted delivery and low site specificity following systemic administration. Thus, we combined the migratory properties of MSCs and IL-21 therapy to produce IL-21-expressing MSCs for targeted treatment. While murine MSCs express low levels of molecules involved in the SDF-1/CXCR4 interaction for tumor tropism, MSCs also express a broad range of chemokine receptors, including CXCR2, CXCR3, CXCR6, CCR6, and CCR9 [41], that may play roles in MSC migration toward tumors. To evaluate the effects of IL-21/MSCs in vivo, we inoculated A20 cells intravenously in mice to develop a disseminated lymphoma model, similar to the clinical situation. We observed hepatosplenomegaly in the tumor-bearing mice, which was previously demonstrated through bioluminescence imaging [42]. IL-21/MSC treatment prolonged tumor-free survival by inhibiting metastasis and tumor cell proliferation in the liver. This therapeutic benefit was associated with the local presence of IL-21 in the liver and spleen, indicating direct delivery of IL-21 to major tumor sites through IL-21/MSCs. Fluorescence-labeled IL-21/MSCs were detected for up to 7 days in the livers and spleens of tumor-bearing mice, indicating prolonged presence and, thus, enhanced antitumor effects of IL-21 at tumor sites. It has been suggested in previous reports that the half-life of circulating IL-21 following systemic administration lasted up to 3 h. We suggest that the prolonged life span of IL-21, through IL-21/MSCs, may allow sufficient time for IL-21 to elicit its therapeutic effects.
In addition to direct apoptotic effects, the main purpose of IL-21 in cancer therapy is the promotion of antitumor immunity. IL-21 stimulates the proliferation and cytotoxicity of CD8+ T cells and also induces the maturation and enhanced cytotoxicity of NK and NKT cells [3]. The ability of IL-21 to enhance NK cell activity supports combination strategies using tumor-targeted monoclonal antibodies, such as rituximab, in B-cell lymphoma [16] for enhanced NK cell-mediated antibody-dependent cellular cytotoxicity. Our results also demonstrate that IL-21/MSCs are effective in activating effector antitumor responses, including IFN-γ-expressing CD4+, CD8+ T cells, NK, and NKT cells. In contrast, these activated effector cells, especially NK cells, may render MSCs susceptible to cell-mediated cytotoxicity. In particular, the low surface expression of MHC class I molecules on MSCs has been shown to increase their susceptibility to NK cell-mediated killing [43,44], and thus, IL-21/MSCs may induce increased NK cell-mediated killing. However, whether the administered MSCs are killed by NK cells in vivo remains unclear. Thus, future molecular imaging studies are needed to provide insights into the cell survival of MSCs and IL-21/MSCs following administration.
In addition, overcoming the regulatory barriers of the immunosuppressive tumor microenvironment is crucial for designing an effective cancer therapy to further promote effector cell activity. While there is some evidence indicating that IL-21 plays a role in the suppression of Treg activity and associated IL-10 levels [45,46], the mechanism of action of IL-21 in inhibiting tumor-mediated immune suppression is still unclear. The effects of IL-21 on immunosuppressive cells, including MDSCs and Tregs, have recently been described in metastatic melanoma [47]. IL-21/MSCs were also able to inhibit immunosuppressive cells, especially MDSCs, and cytokines, including TGF-β and IDO, of the lymphoma microenvironment. Taken together, IL-21 shows potential for restoring balance between activating and regulatory immune responses in lymphoma.
The use of MSC-based gene therapies in cancer is, thus, promising, but the discrepancies among studies as to whether MSCs themselves provide pro- or antitumor effects remain an issue. Immunosuppressive effects of MSCs, through direct cell-to-cell contact and secretion of soluble factors [48], may provide a supportive tumor environment through induction of immune-suppressing cell populations [49], tumor vascular formation [50,51], and epithelial–mesenchymal transition [52]. However, there is also contrasting evidence that MSCs show antitumor effects when injected together with tumor cells. While the underlying mechanism remains to be determined, it is presumed that MSCs downregulate certain signaling pathways, including the Akt [53] and Wnt pathways [54], through cell-to-cell contact and secretion of the dickkopf protein, respectively. In this study, we observed a significant reduction in A20 cell growth by nontransduced MSC culture supernatants in vitro (Fig. 2) versus rIL-21 alone. We showed that nontransduced MSCs produce low levels of antitumor cytokines, such as IFN-β and IFN-γ, which may contribute to the inhibitory effects of MSCs on lymphoma cells. Previously, it was reported that MSCs cultured at a high density secrete higher levels of IFN-α and IFN-β, which contribute to tumor cell growth inhibition [55]. It is unclear, however, whether the upregulation of IFN-α by IL-21/MSCs in our study was due to plating MSCs at a high density for adenovirus transduction or due to overexpression of IL-21 by the cells. Nonetheless, secreted factors present in MSCs may support apoptosis-induced cell death, and the addition of IL-21 through gene transduction in MSCs may have elicited an additive effect. Additionally, it has been reported that MSCs interfere with proliferation of cells of lymphoid origin through cell cycle arrest [56]; however, we did not observe such changes in the cell cycle by MSC treatment. In contrast, MSCs were still ineffective in vivo, suggesting that the immunosuppressive nature of MSCs may override the antitumor potential in a tumor progressive environment or that cell-to-cell interactions within the tumor microenvironment may play a critical role. Similar discrepancies between in vitro and in vivo results of MSCs in lymphoma models have been observed in other studies [56,57].
In conclusion, we report potent therapeutic effects of IL-21/MSCs in a disseminated murine lymphoma model. Among various antitumor genes, IL-21 has multifaceted beneficial effects against lymphoma, including apoptosis-induced cell death, immune activation, and modulation of the tumor microenvironment. MSC-mediated delivery of IL-21 has the potential to maximize the efficacy of treatment in vivo, and thus, we suggest IL-21/MSCs as a novel immunotherapeutic approach in lymphoma. However, further studies are needed to evaluate the effects of IL-21/MSC using human MSCs in a humanized mouse model for future clinical applications. The potential use of third-party MSCs and their translational potential also need to be considered thoroughly.
Footnotes
Acknowledgments
This work was supported by a grant (HI14C3417) from the Korea Healthcare Technology R&D Project, the Ministry for Health, Welfare, and Family Affairs, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.