
Abstract
Recent studies suggest that mature somatic cells can be reprogrammed to become induced pluripotent stem cells by overexpressing specific transcription factors or microRNAs (miRNAs). Theoretically, this technique could provide a wide array of cells for therapeutics. However, the process of redifferentiation after cell reprogramming to pluripotency is inefficient and time restricted. We proposed that the differentiation of somatic cells into specific cells of another germ layer can be induced and accelerated with appropriate miRNAs and culture conditions. In human fibroblasts, we found that overexpression of pluripotency stem cell-specific miRNA-302/367 cluster, together with two other neuron-specific miRNAs (miRNA-9/9* and miRNA-124) induced fibroblasts conversion into neurons. The cells assumed neuron morphology, were positive for several neuron markers, and exhibited neuronal membrane potential feature. Moreover, concentrated expression of synaptic markers were observed in these cells in vitro and in vivo in nude mice brain, suggesting possible connectivity. To achieve efficient reprogramming, miRNA-302/367 cluster, miRNA-9/9*, and miRNA-124 were all required. The combination of the proved pluripotency-inducing miRNA-302/367 cluster and cell-specific miRNAs provides a unique strategy for one-step cellular conversion that could have important implications for studies of neuron development and neurological disease therapy.
Introduction
D
Recently, a promising alternative strategy for direct reprogramming/conversion is the use of microRNAs (miRNAs). The repressive effect of miRNAs on gene expression can be powerful as one miRNA may target multiple pathways simultaneously. Another advantage of miRNA is that chemically synthesized miRNA mimics can be easily administered to cells through lipid-based transfection and exhibit low toxicity in animal models. Moreover, the small size of miRNA allows the packing of multiple transcripts in the same delivery vector to increase both reprogramming efficiency and functional homogeneity of reprogrammed cells.
Cell-specific miRNAs were natural candidates for reprogramming. Among them, the miRNA-302/367 cluster (miR-302/367), including five mature miRNAs (miRNAs-302a/b/c/d and miRNA-367) on chromosome 4 (Fig. 1a, b), is important in the maintenance and self-renewal of ES cells. Studies showed that overexpression of this cluster in somatic cells can efficiently induce reprogramming to pluripotency [11,12]. For somatic cells, muscle-specific miRNAs-1, -133, -208, and -499 combination, without exogenous transcription factor, has been reported to reprogram mouse fibroblasts into cardiomyocyte-like cells both in vitro and in vivo [11,12]. Highly expressed neuron-specific miRNAs-9/9* and -124 (miR-9/124) have been shown to play important roles in the development of the nervous system, especially neural stem cells [13,14]. The expression of these two miRNAs can specifically induce ES cells to become neurons. Coexpression of neuron-specific transcription factors Neurod2, Ascl1, and Myt1l (DAM) can function synergistically with miR-9/124 to induce human fibroblasts into cells with enhanced neuronal function [15]. Recently, it was reported that repression of polypyrimidine tract-binding protein, which regulates miRNA-124, was sufficient to induce fibroblasts into functional neurons [16].
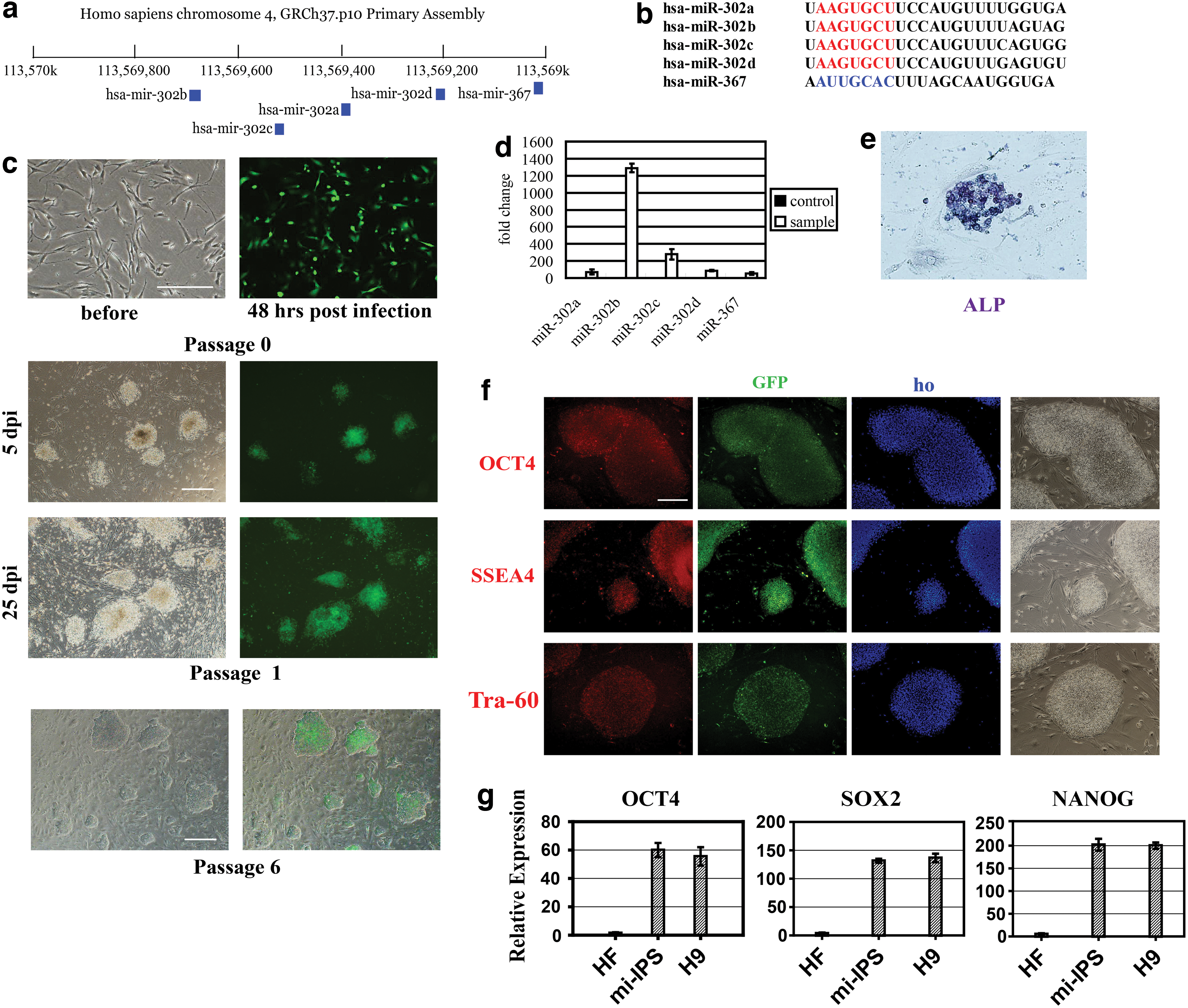
Induced pluripotent stem cells (iPSC) reprogramming, but not neuron conversion, after expression of miR-302/367 in human fibroblasts.
In the present study, it was hypothesized that the expression of the miR-302/367 functions in synergy with miR-9/124 to reprogram fibroblasts into neuronal cells [17]. Results showed that fibroblasts expressing miR-9/124 could reprogram into neurons, although at very low efficiency (∼4%). The expression of miR-302/367 alone did not result in neuron formation from fibroblast. However, coexpression of these miRNAs synergistically promoted fibroblasts reprogramming into neurons at much higher efficiency (∼33%). The converted cells assumed typical neuronal morphology, expressed neuron-specific proteins, formed synaptic connection with neighboring cells, and exhibited electrophysiological features, including action potentials, spontaneous and evoked excitatory and inhibitory synaptic activity.
To verify that treated human skin fibroblasts could convert into neuron and survive in vivo, we injected 5 days postlentiviral infection cells into animal brains, asking (1) do these cells have the same reprogramming capability in vivo as in vitro, (2) can the reprogrammed neurons establish connectivity in the brain. Thirty days after injection, GFP-tagged cells with neuron morphology can be observed in nude mice pallium. Immunohistochemistry study showed that they expressed neuron-specific markers and positive for presynaptic protein synapsin-1 in cluster on soma and processes. These results proved that the infected cells can be reprogrammed into neurons in vivo and showed signs of connections with surrounding wild-type neurons.
Our study showed for the first time that miRNA 302/367 cluster not only played important roles in iPSC reprogramming, but also facilitated direct conversion between somatic cells mediated by other factors. This conclusion may inspire its application in other cell types. In addition, our method of induction of functional neurons from human fibroblast solely by overexpressing miRNAs may reveal a new possibility of clinical future.
Materials and Methods
Fibroblast isolation, cell culture, and molecular cloning
Human fibroblasts were isolated from human cutaneous tissue cultured by tissue patch. Fibroblasts were expanded for four to six passages before using and maintained in 10% fetal bovine serum (FBS), 89% DMEM/F12, and 1% nonessential amino acids. The day before lentiviral infection, fibroblasts were seeded at 1 × 105/cm2 density onto a 24-well plate. Next day, cells were infected with filtered viral supernatants at 1 × 108 TU/mL in the presence of polybrene (5 μg/mL) overnight. Fresh media were replaced 3 days after infection. Cells were digested by trypsin, and cultured in a six-well plate coated with poly-L-lysine and laminin, maintained in media containing 1% N2 supplement, 2% B27 supplement, 48% neurobasal medium, 48% DMEM/F12, 1% nonessential amino acids, and 5 nM 2-mercaptoethanol. Two weeks later, cells were treated with 2.5 μg/mL cytarabine to inhibit the gliocyte from growing.
Immunofluorescence and flow cytometry
Cells were washed with phosphate buffered saline (PBS) and fixed with 4% paraformaldehyde for 15 min, then incubated in PBS with 0.3% Triton X-100 for 10 min at room temperature. After washing three times with PBS, cells were blocked in PBS with 3% bovine serum albumin (BSA) and 2% FBS for 1 h at room temperature. Primary and secondary antibodies were diluted in blocking solution and incubated for 1 h each. Cells were washed three times with PBS between antibodies. The cell nucleus was stained with Hoechst 33342. The following antibodies were used: rabbit anti-MAP2 (1:300; Santa Cruz), mouse anti-β III Tubulin (1:200; R&D), mouse anti-GABA (1:200; Abcam), rabbit anti-NeuN (1:500; Cell Signaling), rabbit anti-doublecortin (1:500; Cell Signaling), and rabbit anti-synapsin-1 (1:200; Cell Signaling). The secondary antibodies were goat anti-rabbit or mouse IgG conjugated with Alexa 647 (1:800; Jackson Immuno).
Patch clamp recording
Recordings were performed on fibroblasts 4 weeks after infection. Recording solutions: extracellular solution (mM): NaCl 145, KCl 4, CaCl2 2, MgCl2 1, glucose 10, HEPES 10, pH 7.4, and osmolarity ∼295 mOsm; intracellular solution (mM): KOH 31.25, KCl 120, EGTA 10, MgCl2 1.75, CaCl2 5.374, HEPES 10, Na-ATP 4, pH 7.2, and osmolarity ∼285 mOsm. The extracellular solution was used within 10 days of preparation. The intracellular solution was prepared in aliquoted batches and stored at −20°C until used.
Recording solutions for evoked EPSC and IPSC: extracellular solution (mM): NaCl 140, KCl 2.5, CaCl2 2.5, MgCl2 2, NaH2PO4 1, glucose 20, HEPES 10, pH 7.4; intracellular solution (mM): CsMeS 135, CsCl 5, HEPES 10, EGTA 0.5, MgCl2 1, Mg2ATP 4, NaGTP 0.4, QX-314 5, pH 7.4 CsOH.
Cells were tested at room temperature using whole-cell patch clamp techniques. Output signals from the MultiClamp 700B amplifier was digitized using a Digidata 1440 A/D D/A board, low-pass filtered at 1 kHz. Glass micropipettes were used as recording electrode in the study. The pipettes were constructed from thin-walled glass (1.5 mm diameter, WPI) on the same day of patch-clamp experiment. With ICS filled, the pipette resistance was between 2 and 5 MΩ.
In vivo experiment
The housing facility is in keeping with the national standard Laboratory Animals–Requirement of Environment and Housing Facilities. The care of laboratory animals and the animal experimental operations conform to the PRC Administration Rule of Laboratory Animals.
Experimental animals were 3 weeks nude mice, BALB/c mice, and Sprague Dawley rats, all female. Major equipment: stereotaxic apparatus, fluorescence microscope, microtome. Reagents: 1.5% pentobarbital (5 mL/kg body weight), PBS, 4% paraformaldehyde, Triton X-100, 3% BSA. The same antibodies used on isolated cells were used on brain tissue sections.
Cells were infected with lentivirus as described above. Five days after infection, cells were digested with 0.05% Trypsin-EDTA and dispersed evenly. Animals were anesthetized and injected with 105 cells into forceps minor of the corpus callosum at the speed of 10 μL/5 min. Animals were executed 10 days–1 month later and brains were fixed in paraformaldehyde for 24 h. Samples were sliced into 10 μm–20 μm sections. Immunohistochemistry and Hematoxylin and eosin stainings were carried out.
Results
Expression of miR-302/367 caused iPSC reprogramming, but not neuron conversion
Cutaneous fibroblasts were obtained from normal human skin tissue. The fibroblast culture was free of neuron progenitors as assessed by MAP2 immunoreactivity.
A lentiviral vector expressing miR-302/367 (miRNAs-302a/b/c/d/367) under an EF-1α promoter, as well as a GFP reporter under the CMV promoter, was generated to infect human fibroblasts. Significant green fluorescence was observed in cells 48 h after infection, indicating efficient transduction of the virus (Fig. 1c). Over 98% fibroblasts counted were GFP positive with the virus multiplicity of infection of 100. Real-time quantitative polymerase chain reaction (PCR) confirmed the expression of miRNAs-302a/b/c/d and miRNA-367 (Fig. 1d). Infected fibroblasts were cultured on feeder cells with ES cells media. After 2 weeks culture, change in cell morphology was noted—clone-like cells emerged and separated from the feeder layer. Two weeks later (4 weeks postinfection) those clones were picked and passaged. These passaged clones tested positive for ALP activity and retained GFP activity, consistent with the expression of transduced miRNAs (Fig. 1e). Passaged clones also stained positive for OCT4, SSEA4, and TRA-60, markers of pluripotency (Fig. 1f). Real-time PCR for OCT4, SOX2, and NANOG further proved that cells were pluripotent (Fig. 1g).
To determine if these cells can be reprogrammed directly to neurons, cells infected with miR-302/367 virus were seeded onto wells coated with poly-L-lysine and laminin, and cultured in a neural medium. However, no emergence of neuron-like cells was observed in culture after 1 month. The failure of these cells to reprogram to neural cells demonstrated that complete conversion did not occur.
Expression of miR-302/367 efficiently facilitated miR-9/124-induced fibroblast reprogram into functional neuron
To test how miR-302/367 could contribute to neuron conversion, cells were divided into control group overexpressing miR-9/124 only, and experimental group overexpressing both miR-302/367 and miR-9/124. After lentiviral infection, cells were proliferated for 5 days, and then replated onto poly-L-lysine and laminin-precoated six-well plate. The medium was changed to Neurobasal medium plus N2/B27. Cells were cultured for 21 days postinfection, and observed daily (Fig. 2a). Eight batches were repeated.

Human fibroblasts reprogram to functional neurons under coexpression of miR-302/367 with miR-9/124.
Figure 2b showed that on day 7, there was no obvious change of cell morphology. On day 14, some cells changed from flat or oblate into round shape. Comparing with wild-type fibroblasts, infected cells contracted into tiny round cell body and sprouted some small dendrites. Some intermediate cells also had dendrites, but their cell bodies still remained a fibroblast shape. This might be attributed to incomplete reprogramming. On day 21, the number of cells with tiny round cell body and dendrites increased greatly. The number of incomplete reprogrammed cells (with dendrites, but oblate cell body) increased as well. Cells were photographed and counted 10 frames per well for converted/total cell numbers (Fig. 2c). The average ratio of reprogrammed cells was 5.46% (STDEV = 4.67) in control group versus 38.02% (STDEV =7.67) in the experimental group (P < 0.01). Meanwhile, these projected processes grew interlaced and formed a network (Fig. 2d). Under high magnification, it could be observed that the same cell body had both long and short processes (Fig. 2e).
To further prove that those cells with the morphological changes were true neuron, antibodies for neuron-specific markers MAP2, NeuN, TuJ1, DCX, and GABA were used for immunofluorescence staining. As shown in Fig. 3a, reprogrammed cells were positive for tested markers. Real-time PCR was performed for several markers (MAP2, TUBB3, NEFL, RBFOX3, DPF1, SYN1, GABBR1, and NES) as well as in 2, 10, and 21 dpi cells. Data showed that on 21 dpi, in cells infected with miR-302/367 plus miR-9/124, mRNA levels of tested neuronal markers were significantly higher (P < 0.05) than those in cells infected with only miR-9/124 (Fig. 3b). Western blot for MAP2 and TuJ1also supported the increase of these two proteins in miR-302/367 plus miR-9/124 over miR-9/124 alone group (Fig. 3c). MAP2 and TuJ1were also used for fluorescent-activated cell sorting on 7, 14, 21 dpi cells (Fig. 3d), and the change of numbers was statistically analyzed (three batches per time point). At 21 dpi, MAP2-positive cells were 4.07% versus 33.78%, and TuJ1-positive cells were 5.80% versus 38.90% in the control and experimental groups, respectively (Fig. 3e).

The characterization of reprogrammed neurons.
Reprogrammed cells exhibited action potential and established synaptic activity
Using whole-cell patch recording (Fig. 4a), we found that applying a series of voltage steps to the cells evoked distinct inward currents on top of the outward currents (Fig. 4b). The current–voltage (I–V) curve of inward currents (Fig. 4c, left) displayed voltage-gated sodium channel features, whereas I–V of outward currents displayed the features of voltage-gated potassium channels in the cells (Fig. 4c, right). In addition, 1 μM tetrodotoxin (TTX) completely and reversibly blocked the initial inward current, confirming that the inward current was due to voltage-gated sodium channels (Fig. 4d). Current clamp recording from a human fibroblast-derived neuron showed cell firing when depolarized (Fig. 4e).
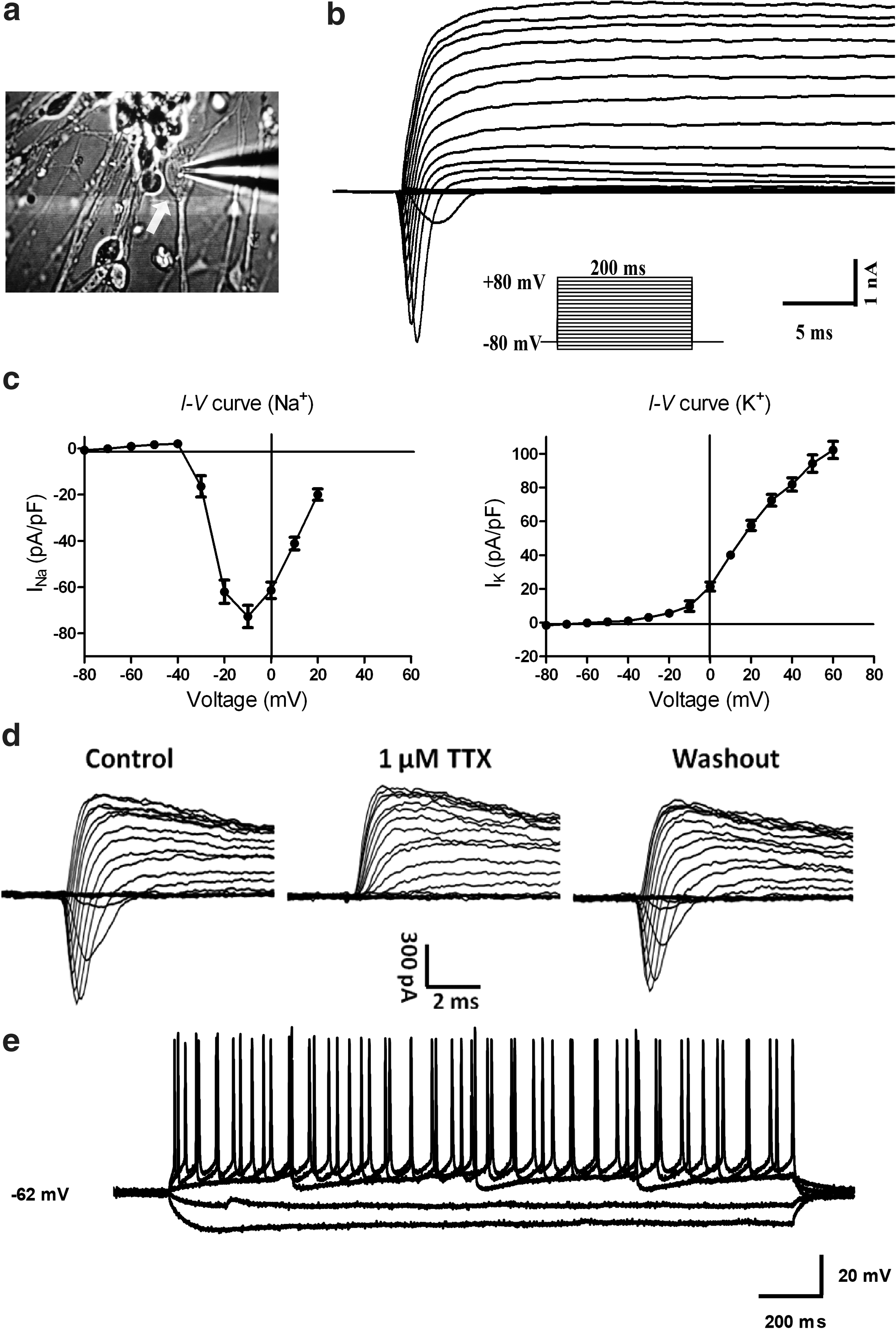
Patch clamp study on human fibroblast-derived neurons.
Real-time quantitative polymerase chain reaction (Q-PCR) data suggested that the presynaptic marker synapsin-1 was upregulated in reprogrammed cells. Immunocytochemistry study further showed its accumulated presence in cell processes (Fig. 5a), suggesting that these network-forming cells had made contact and connections. Converted cells were able to fire repetitive action potentials in response to depolarizing current injections. Moreover, some of these cells exhibited synaptic transmission similar to primary neurons. In the presence of 1 μM TTX, mini spontaneous postsynaptic currents were seen in 10 out of 14 induced cells (Fig. 5b). This mini current was completely blocked by 6,7-dinitroquinoxaline-2,3-dione (DNQX) and 2-amino-5-phosphonopentanoic acid (APV), indicating that it was mini excitatory postsynaptic current (mEPSC). Furthermore, the converted cells exhibited evoked EPSCs (blocked by DNQX plus APV, Fig. 5c) and inhibitory postsynaptic currents (IPSCs) in response to local stimulation (blocked by bicuculline, Fig. 5d).
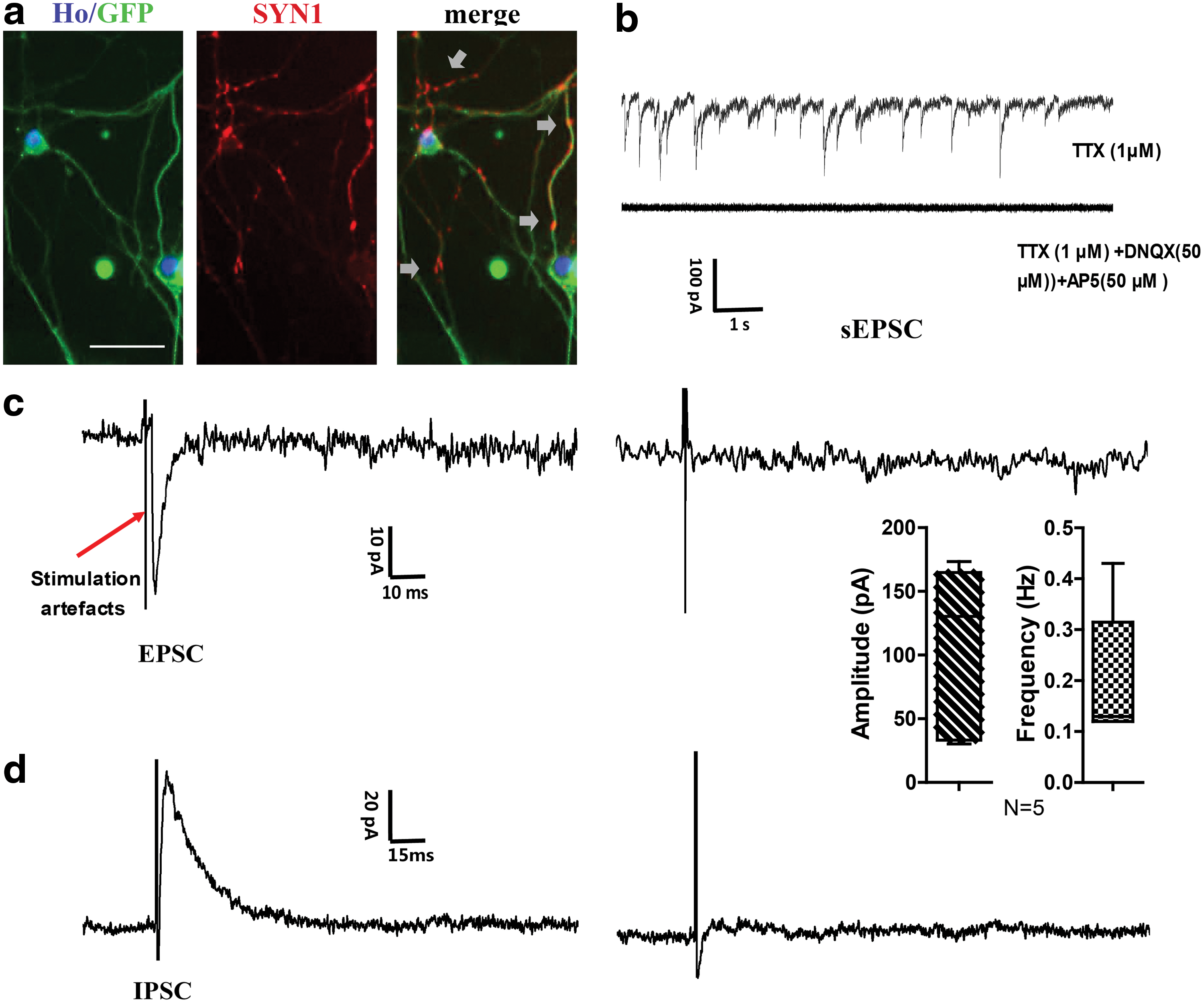
Patch clamp study on human fibroblast-derived neuron network.
Human fibroblasts can be reprogrammed to neurons and showed signs of connections in nude mice brain
Six batches of 5-day postlentiviral infection cells were injected into the hippocampus (Fig. 6a) of 10 nude mice, 10 BALB/c mice, and 4 Sprague Dawley rats (all female). Brains were sampled 7 days, 14 days, and 1 month postinjection. Frozen sections at 10–20 μm were obtained for staining. In normal mice and rats, the injected cells were barely observed, possibly been eliminated, and metabolized by their immune system. However, injected cells survived in the nude mice brain. The exogenous GFP-positive cells, located at the pallium of the brain (Fig. 6b), was in accord with the natural neuron location. It indicated that the cell may have directional growth and transfer. The reprogramming process cost at least 21 days in vitro. While in vivo, GFP-positive cells with neuron morphology were only observed in 30 dpi brains (Fig. 6c), but not in 7 or 14 dpi brains. Therefore, the reprogramming time span was accordant for both in vitro and in vivo. Immunohistochemistry study showed that these GFP-positive cells expressed certain neuronal markers, such as TuJ1 (Fig. 6d), MAP2, DCX, and NeuN (Fig. 6e). Synapsin-1, a synase marker, was also found in some GFP-positive cells, suggesting that these exogenous cells were building up synaptic connection with surrounding wild-type neurons (Fig. 6e).
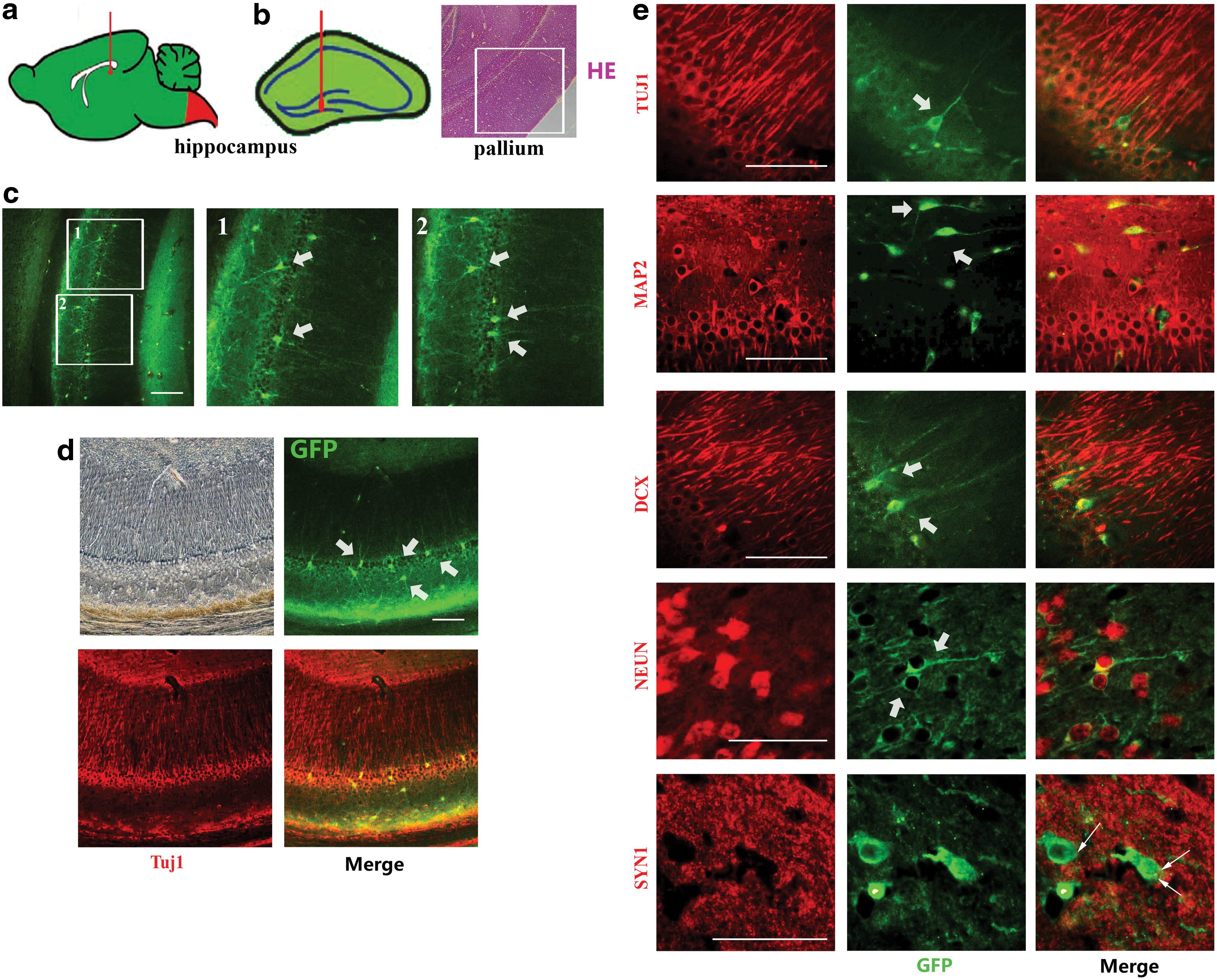
In vivo study of human fibroblast-derived neurons in nude mice brain.
These results convincingly proved that these lentiviral-infected fibroblasts can be induced to convert into neurons in the microenvironment in vivo and establish synaptic connections in nude mice brain. However, it was very rare to observe GFP-positive cells in normal mice and rat brain groups. We speculate it may be because exogenous cells were cleared by natural metabolism or immune system.
Discussion
Recent studies discovered that miR-9/124 instructed compositional changes of SWI/SNF-like BAF chromatin-remodeling complexes, a process important for neuronal differentiation and function [4]. However, overexpression of miR-9/124 alone in human fibroblasts only resulted in low percentage of MAP2-positive cells (less than 5%). By adding three neurogenic transcription factors, NEUROD2, ASCL1, and MYT1L (DAM), which are important for converting mouse embryonic fibroblasts into functionally mature neurons, ∼80% fibroblasts converted into MAP2-positive cells. This result was seen in both human neonatal foreskin fibroblast and adult dermal fibroblast.
MiR-302/367 was formerly known to function in iPSC reprogramming. Recent research has shown that overexpression of miR-302/367 can induce somatic cell reprogramming efficiently [18]. It was reported that miR-302/367 reprogramming was more efficient than OSKM reprogramming both in mouse and human fibroblasts, and miR367 expression was required for miR-302/367 iPSC reprogramming.
In the present study, the neural-specific miRNAs, miR-9/124, converted fibroblasts into neurons with low efficiency (<4.07% MAP2-positive cells), consistent with reported data (<5% [15]). Furthermore, coexpression of miR-302/367 with miR-9/124 resulted in a more efficient conversion (∼33.78%, Fig. 3e).
However, unlike miR-302/367 alone infected and ES medium-fed fibroblasts (Fig. 1c), typical iPSC clones were not seen in the coinfected cells (Fig. 2b) during culture. These results suggested that complete reprogramming (ie, iPSC formation) may not be necessary for conversion in all cases. A likely explanation is that the overexpression of the miRNA-302/367 cluster leaves fibroblasts in a relatively unstable intermediate state that maintains pluripotency. The resulting cells have certain differentiation potential and, therefore, can be effectively converted to neurons under neuron-specific miRNAs such as miR-9/9* and -124.
Reports have confirmed that miR-302/367 is abundant in ES cells, and that its expression inhibits a series of epigenetic factors, such as AOF1/2, methyl-CpG-binding proteins 1 and 2, and Dnmt1. MiR-302/367 can induce extensive demethylation of DNA, which promotes activation of Oct4 and Sox2, both are important to maintain stem cell pluripotency [19]. Genomes of cells that express miR-302/367 tend to be in an active state, with greatly increased plasticity. Therefore, when provided a suitable induction environment, cells are susceptible to being converted. Compared to the harsh conditions required by complete reprogramming and the extremely low efficiency of iPSC clones, the induction efficiency of coexpressing neural miRNAs is much higher. Studies have confirmed that the expression of transcription factors for reprogramming can play a similar role in determining cell fate after iPSC formation. Expression of OSKM factors complemented by appropriate cytokine signaling in mouse fibroblasts can also yield similar results, further supporting this hypothesis.
Since no significant emergence of neural stem cells or neural precursor cells was observed in our study, early steps in neural biogenesis may have been bypassed. Rosette formations, which are the typical structure of the neural precursor cells, were not observed, with the expression of both miR-302/367 and neural miRNAs. Although further studies are needed to confirm to what extent neural precursor cells were developed during the conversion, this phenomenon showed that the conversion bypassed the complete reprogramming process to neural precursors.
The characterization of the reprogrammed cells included their neuron-like morphology, expression of neuronal markers, as well as electrophysiological studies. The presence of sodium channels was confirmed and action potential can be invoked on these cells. They exhibited spontaneous, evoked excitatory and inhibitory synaptic activities, implying formation of functional connectivity-like neural circle. Furthermore, consistent with the Q-PCR results, the presynaptic marker synapsin-1 was found on reprogrammed cells, indicating the formation of synapses and signal transduction between cells.
The in vitro study proved that our genetically modified cells can convert into cells with certain functioning characteristics of neurons. Theoretically, intrinsic micro environment, including extracellular matrix, intercellular material transfer, signal transmission, and small molecules supplement, would be more friendly for cell differentiation. Our in vivo study showed that injected cells survived and had direction, as most GFP-positive cells were sited at the pallium, their corresponding locus. Presence of neuronal markers in these cells confirmed their specificity. Moreover, cluster appearance of synapsin-1 suggested that reprogrammed cells were attempting to establish synaptical connections and possible communication with surrounding native neurons. In conclusion, our findings have proved that these infected cells have reprogramming capability both in vivo and in vitro.
Although we have not established animal models for behavioral study and future clinical application requires further studies, the approach described in this study provides a conversion pathway from somatic cells to nerve cells by expressing miRNAs, and it offers a new vision that other types of cells might also be obtained through an intermediate state with a specific pathway of transdifferentiation.
Footnotes
Author Disclosure Statement
No competing financial interests exist.