
Abstract
Diabetic trauma patients exhibit delayed postsurgical wound, bony healing, and dysregulated bone development. However, the impact of diabetes on the pathologic development of ectopic bone or heterotopic ossification (HO) following trauma is unknown. In this study, we use leptin-deficient mice as a model for type 2 diabetes to understand how post-traumatic HO development may be affected by this disease process. Male leptin-deficient (ob/ob) or wild-type (C57BL/6 background) mice aged 6–8 weeks underwent 30% total body surface area burn injury with left hind limb Achilles tenotomy. Micro-CT (μCT) imaging showed significantly lower HO volumes in diabetic mice compared with wild-type controls (0.70 vs. 7.02 mm3, P < 0.01) 9 weeks after trauma. Ob/ob mice showed evidence of HO resorption between weeks 5 and 9. Quantitative real time PCR (qRT-PCR) demonstrated high Vegfa levels in ob/ob mice, which was followed by disorganized vessel growth at 7 weeks. We noted diminished chondrogenic gene expression (SOX9) and diminished cartilage formation at 5 days and 3 weeks, respectively. Tartrate-resistant acid phosphatase stain showed increased osteoclast presence in normal native bone and pathologic ectopic bone in ob/ob mice. Our findings suggest that early diminished HO in ob/ob mice is related to diminished chondrogenic differentiation, while later bone resorption is related to osteoclast presence.
Introduction
R
Similar to normal fracture healing, HO is believed to develop through an endochondral intermediary [4,5]. Metabolic derangements, including diabetes, can result in decreased bone mineral density and impaired fracture site healing in normal bone [6 –8]. Leptin-deficient mice, which exhibit hyperglycemia and obesity similar to patients with type 2 diabetes, are known to exhibit diminished bone volume and osteoporosis [9,10]. Mesenchymal stem cells (MSCs) cultured in the presence of high-glucose exhibit diminished chondrogenesis [11]. Additionally, osteoclast upregulation may contribute to bone turnover in diabetic patients [12,13]. Understanding how diabetes affects HO could elucidate new strategies for inhibiting or limiting this debilitating condition.
We have previously described a mouse model of HO using a systemic 30% total body surface area (TBSA) partial-thickness burn with Achilles tenotomy of the hind limb [14]. Wild-type mice (C57BL/6) develop heterotopic cartilage within 3 weeks post-trauma and undergo endochondral ossification by 5 weeks. In this study, we investigate the impact of leptin deficiency, as a model of type 2 diabetes, on heterotopic bone formation using our in vivo mouse model of burn with tenotomy in ob/ob and wild-type mice and our in vitro model of adipose-derived mesenchymal stem cell (adMSC) culture [14].
Materials and Methods
Mice
Animal procedures were carried out in accordance with guidelines from the Guide for the Use and Care of Laboratory Animals: Eighth Edition, approved by the Institutional Animal Care and University Committee on the Use and Care of Animals of the University of Michigan (UCUCA PRO0001553). Young (10–12 weeks), male, leptin-deficient diabetic mice (ob/ob) [15,16] on a C57BL/6 background and wild-type C57BL/6 mice purchased from Jackson Laboratory were used for all experiments. Mice were housed three per cage at 72F (±4F) with 12 h of light exposure daily (325 lux) and fed 5LOD chow.
Burn injury and tenotomy
All mice intended for in vivo development of HO underwent 30% TBSA partial-thickness burn injury to the dorsum under 3% inhaled isoflurane [2]. A metal block was heated to 60°C in a water bath and placed over the shaved dorsum for 18 s. Following burn injury, mice received a left hind limb Achilles tenotomy using sterile surgical scissors, and the incision was closed with a single 5-0 vicryl suture. Animals were housed in UCUCA-supervised facilities, three per cage, receiving 12 h of light exposure daily, with no restrictions to diet. All mice were housed in the same facility.
Osteogenic differentiation of adMSCs
Ob/ob and wild-type mice were prepared for harvest of adMSCs [17]. Mice underwent 30% TBSA burn without tenotomy or no burn injury at all. After 2 h, mice were euthanized and adMSCs were harvested from the bilateral inguinal regions. AdMSCs were chosen as a proxy for the still unidentified mesenchymal cell responsible for HO formation [14] and have shown capacity for bone formation in vivo and in vitro [2].
MSCs were seeded in triplicate onto 6- and 12-well plates and cultured in the osteogenic differentiation medium (DMEM with 10% FBS, 1% penicillin/streptomycin, 10 mM β-glycerophosphate, 100 μg/mL ascorbic acid. Invitrogen). Early osteogenic differentiation was assessed by an alkaline phosphatase (ALP) stain after 7 days [2]. Alizarin red staining for bone mineral deposition was completed at 2 weeks [2].
μCT imaging and analysis
In vivo development of HO was assessed with longitudinal μCT scans at 5, 7, and 9 weeks after trauma for each mouse (n = 3 for ob/ob and for wt/wt mice) (μCT: GE Healthcare Biosciences, using 80 kVp, 80 mA, and 1,100 ms exposure). Images were reconstructed and HO volume was computed using a calibrated imaging protocol [18]. Heterotopic bone was calculated between Hounsfield units of 1,250 and 1,800. The voxel size for all scans is 48.08 μm, and the field of view is 38.47 × 38.47 cm. Scans were performed from the most distal aspect of the foot to the mid-thigh. The region of interest evaluated for HO extended from the knee to the foot.
MICROFIL imaging and analysis
In vivo collateral vessel formation was assessed using MICROFIL [19]. At week 7, ob/ob and wild-type mice (n = 3 each) were euthanized and perfused through the descending thoracic aorta MICROFIL (MV120-blue; Flow Tech Inc.) until it flowed out through the vented inferior vena cava. MICROFIL was allowed to polymerize overnight at 4°C within the vasculature. μCT imaging was performed to assess vasculature. Total MICROFIL volume was computed using a calibrated imaging protocol [20].
Histology
Before injury, or at 5 days (n = 2), 3 weeks (n = 2), and 3 months (n = 5) after burn injury/tenotomy, animals were euthanized and the tenotomized legs from mid-thigh onward were fixed, decalcified in 19% EDTA, paraffin embedded, and sectioned at 5 μm. Slides were stained with pentachrome for cartilage and ectopic bone and Tartrate-resistant acid phosphatase (TRAP) for osteoclast presence [14,21]. Immunohistochemical staining was performed with the following primary antibodies at 1:100 dilutions: anti-HIF-1α, anti-VEGF-A, anti-SOX9, and anti-CD31.
RNA collection and qRT-PCR from histologic slides
Histologic sections of the left hind limbs from burned ob/ob and wild-type mice were collected 5 days post-trauma (n = 2). Additionally, the histologic section of the left hind limb from uninjured unburned ob/ob and wild-type mice was also collected. Local soft tissue at the tenotomy site posterior to tibia/fibula was isolated at 20× magnification and digested in Proteinase K, followed by QIAzol lysis buffer (Qiagen) and chloroform extraction. mRNA was treated with DNAse Turbo (ThermoFisher Scientific) and reverse transcribed to cDNA using TaqMan Reverse Transcription Reagents (Applied Biosystems). Quantitative real time PCR was completed using SYBR Green PCR Master Mix (Applied Biosystems). Specific primers for these genes corresponded to their PrimerBank sequences. Transcript levels were standardized to internal gapdh transcript levels. All reactions were run in triplicate.
Statistical analysis
Data were analyzed using SPSS software (v21, IBM). Means and standard deviations were calculated, and statistical analysis was performed using Student's t-test. In figures, bar graphs represent means, whereas error bars represent one standard deviation. For all assays, significance was defined as P < 0.05.
Results
Ob/ob mice produce significantly less HO after injury
μCT imaging was performed at 5–9 weeks after injury to quantify total HO volume. The HO volume in ob/ob mice was significantly lower than wild-type mice at each time point with the greatest difference existing at 9 weeks. At 9 weeks, the HO volume in ob/ob mice was 0.70 mm3 (SD 0.23 mm3) versus 7.02 mm3 (SD 1.42) in wild-type mice (P < 0.01) (Fig. 1A, B).
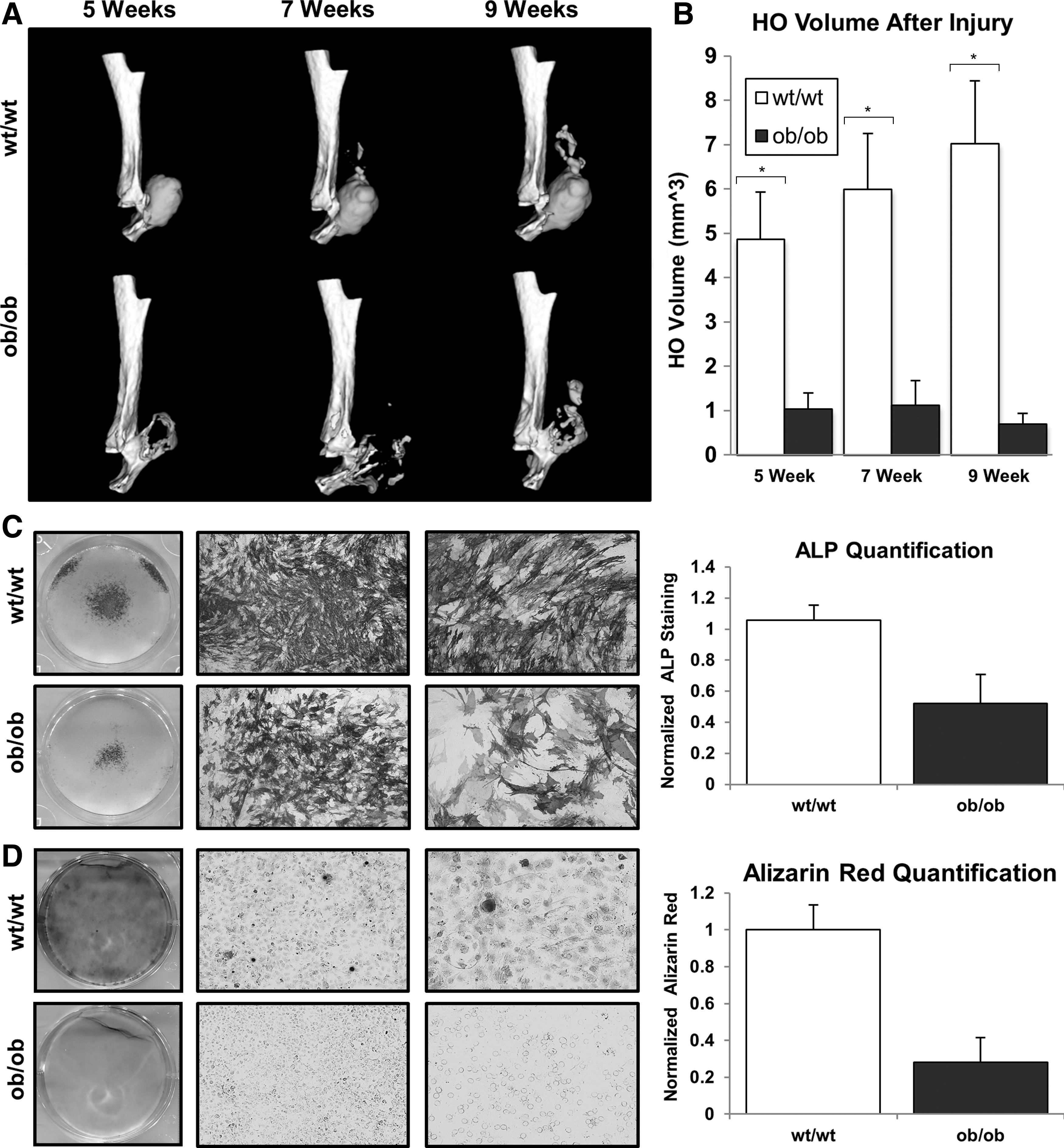
AdMSCs were harvested from ob/ob and wild-type mice 2 h after burn injury. Both ALP and alizarin red staining confirmed significantly diminished osteogenic differentiation of adMSCs from burned ob/ob mice at 7 and 14 days after initiation of osteogenic culture, respectively (Fig. 1C, D). Similarly, we noted that unburned wild-type adMSCs were more osteogenic with respect to ALP expression compared with ob/ob adMSCs (Supplementary Fig. S1; Supplementary Data are available online at
Ob/ob exhibit evidence of hyperglycemia and hyperinsulinemia
To confirm that the ob/ob mice in our study approximated type 2 diabetes, we assessed serum glucose and insulin (Supplementary Fig. S2). Using serum analysis of the specific mice in our study, we found that at both 5 days and 3 weeks after injury, the ob/ob mice had significantly higher serum glucose levels compared with wild-type mice at the same time points after injury (day 5: 700 mg/dL vs. 281 mg/dL, P < 0.05; week 3: 663 mg/dL vs. 273 mg/dL, P < 0.05). Serum insulin levels were also significantly higher in ob/ob mice compared with wild-type mice (day 5: >300 μM/mL vs. 7.3 μM/mL, P < 0.05; week 3: >300 μM/mL vs. 10.3 μM/mL, P < 0.05). Additionally, we found that the hind limbs of ob/ob mice weighed significantly more compared with wild-type mice (1.41 g vs. 0.63 g, P < 0.05). Finally, we noted that the uninjured right hind limb of ob/ob mice exhibited substantially more adipose tissue compared with the uninjured right hind limb of wild-type mice.
Ob/ob mice exhibit more angiogenic signaling at the trauma site compared with wild type
Mean vessel volumes (MVV) in the injured hind limb of burned diabetic and wild-type mice at 7 weeks after injury were compared (Fig. 2A). MVV in diabetic mice were significantly greater than wild type (9.47 mm3 vs. 3.22 mm3, P < 0.01). Although difficult to quantitate, the vessels in ob/ob mice are narrower and more disorganized. When adjusting for limb mass, diabetic limbs continued to have increased MVV (6.87 mm3/g vs. 5.31 mm3/g, P < 0.01) (Fig. 2B). At 3 weeks, we found more small vessels in diabetic mice based on CD31 staining (Fig. 2C). Upon staining of wild-type and ob/ob hind limbs in the absence of burn/tenotomy, we found more CD31+ vessels in wild-type mice suggesting that baseline vessel presence is elevated in uninjured wild-type mice (Supplementary Fig. 3A, B).

At 5 days post-trauma, there was similar staining for Hif1α at the hind limb of diabetic and wild-type mice (Fig. 2D). RNA transcript levels of Hif1α were higher in histologic sections of the diabetic hind limb at the injury site (Fig. 2E). A similar comparison of Vegfa levels showed more Vegfa staining in the ob/ob hind limb, again confirmed based on RNA transcript levels at 5 days post-trauma (Fig. 2F, G). We found elevated Vegfa immunostaining in uninjured wild-type mice compared with uninjured ob/ob mice, although RNA analysis did not suggest a significant difference (Supplementary Fig. 3C, D).
Decreased cartilage formation in ob/ob mice at 3 weeks and 3 months postinjury
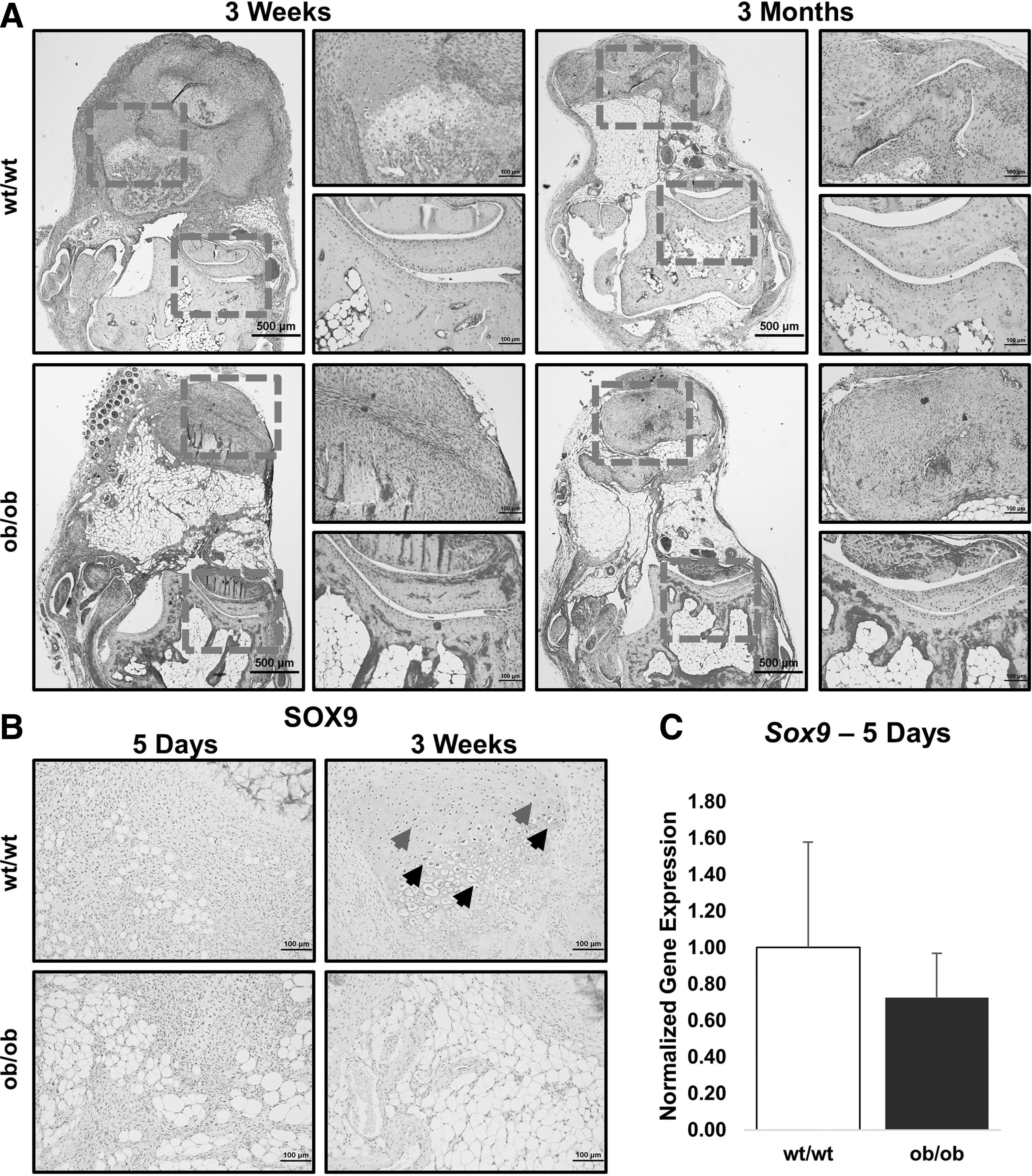
Despite increased angiogenic signaling based on the RNA transcript level, protein expression, and final vessel growth, pentachrome and SOX9 staining confirmed that ob/ob mice exhibit diminished cartilage formation. At the level of tenotomy, we found robust alcian blue staining representative of cartilage after 3 weeks in pentachrome-stained sections of wild-type mice only (Fig. 3A). Ob/ob mice exhibited less SOX9 staining compared with wild type at the site of future HO development 3 weeks after injury, coinciding with areas of blue stain in our pentachrome-stained sections (Fig. 3B). RNA analysis did not indicate a significant difference in Sox9 expression early after trauma, however (Fig. 3C).
Ectopic bone has diminished collagen and osteoblast presence in ob/ob mice
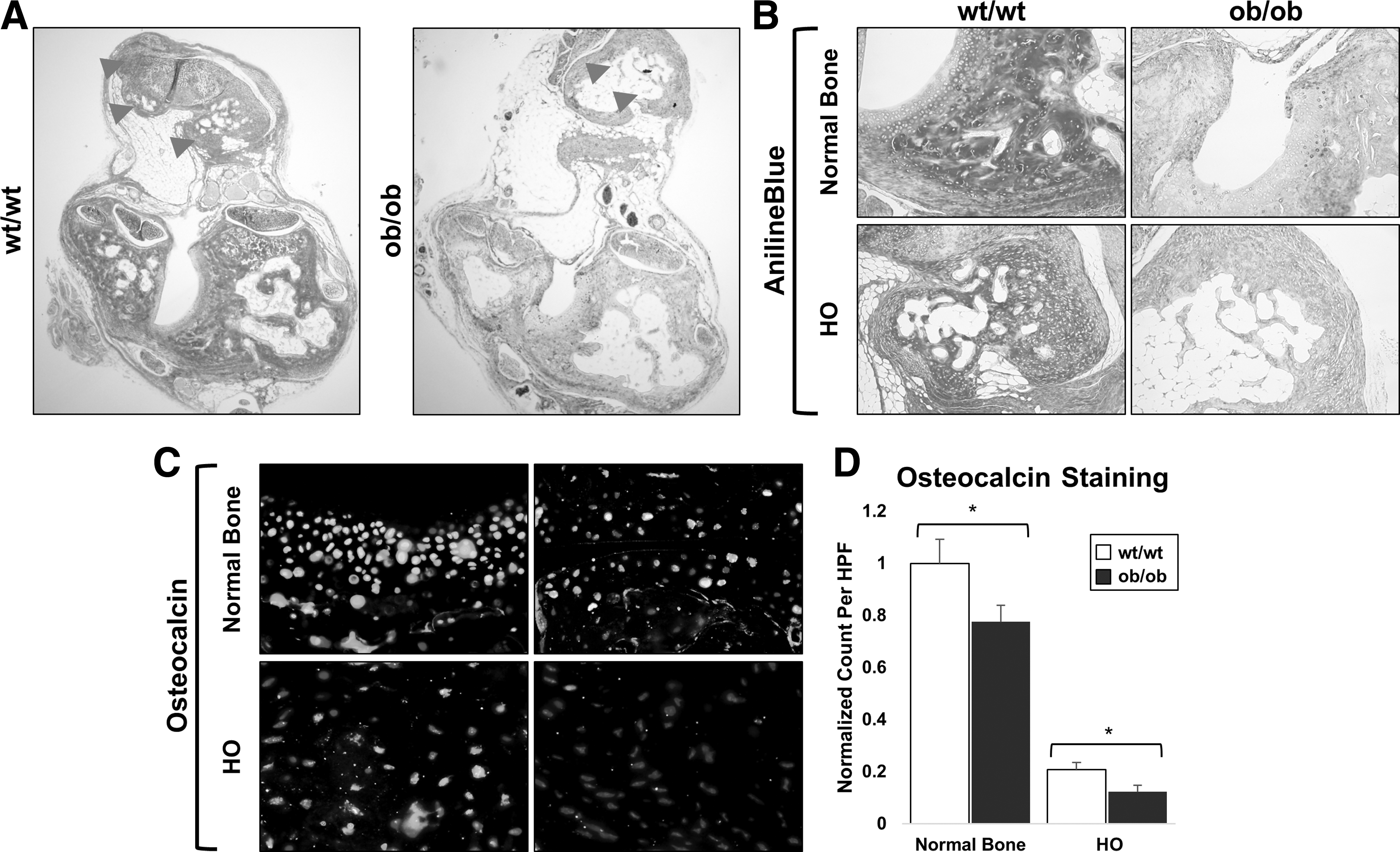
Ectopic and normal cortical bone from wild-type and ob/ob mice was stained with aniline blue. Both ectopic bone and normal cortical bone from ob/ob mice had diminished osteoid staining compared with respective bone from wild-type mice (Fig. 4). Furthermore, osteoblast presence was diminished within the ectopic bone of ob/ob mice compared with wild-type mice by osteocalcin immunofluoresence (Fig. 4).
Ectopic bone resorbs during the initial weeks after injury in ob/ob mice
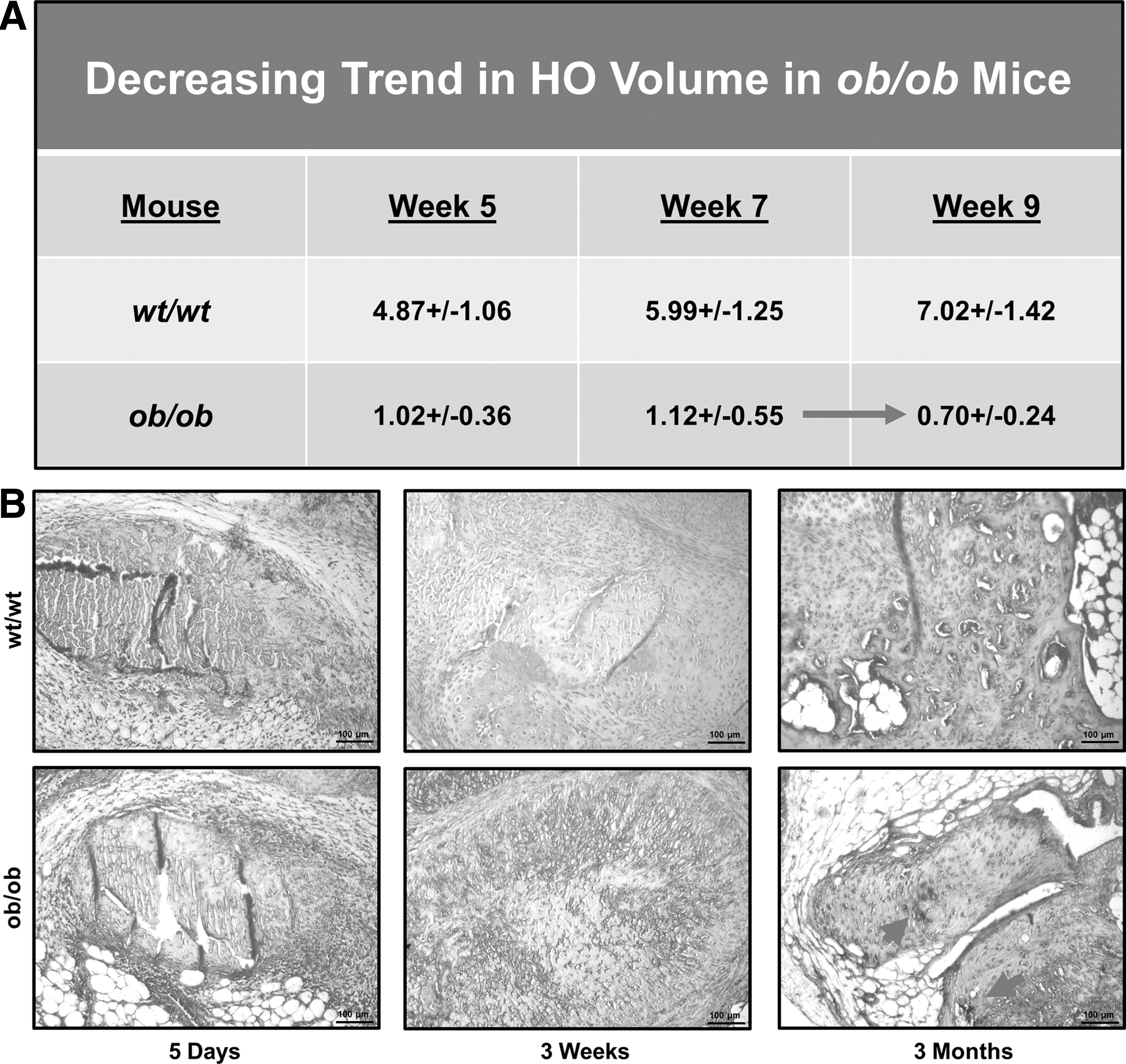
Upon further evaluation of the μCT images, we noted decreases in HO volume from weeks 5 to 7, and again from weeks 7–9, indicative of bony resorption in ob/ob mice (Fig. 5A). HO resorption was not observed in wild-type mice at any time point [14].
Increased osteoclast presence in heterotopic bone from ob/ob mice
Tartrate-resistant acid phosphatase (TRAP) staining of ob/ob and wild-type mice was performed at 5 days, 3 weeks, and 3 months post-trauma to assess osteoclast presence. While wild-type HO had no evidence of osteoclast activity, we found robust osteoclast presence at the 3-month time point in HO from ob/ob mice (Fig. 5B). This osteoclastogenesis is not seen even if wild-type mice are followed out to 15 weeks demonstrating dysregulated bone homeostasis in our ob/ob mice.
Discussion
In this study, we use leptin-deficient mice (ob/ob) as a model of type 2 diabetes to determine the impact on HO following trauma. HO is a condition characterized by ectopic bony lesions within soft tissues. This condition often develops in patients with severe trauma such as large surface area burns, blast injuries, or surgical procedures. As a result, patients develop chronic pain, open wounds, restricted range of motion at joints, and nerve impingement [1 –3]. Our interest in studying HO in the context of a model of type 2 diabetes was driven by previous studies demonstrating increased fracture risk and impaired fracture healing in diabetic patients and animal models, as well as less incidence of HO in diabetes patients [6 –8]. No studies have examined the impact of diabetes on the production of pathologic heterotopic bone post-trauma. By identifying the impact of a pathologic metabolic condition on pathologic bone formation, we may begin to identify potential targets to prevent or limit HO in patients.
Our model of post-traumatic HO reliably produces radiographically evident HO as early as 5 weeks after injury in wild-type mice and closely resembles patients at risk for HO—those with large burns and/or significant trauma. By using a leptin-deficient model (ob/ob), we focused on a model of diabetes closely resembling the metabolic syndrome experienced by type 2 diabetic patients, which has been shown to closely resemble pathologic wound healing in diabetic patients [22]. Serum glucose and insulin analyses of the mice included in our study confirm that these mice demonstrate some of the components of type 2 diabetes.
We found that ob/ob mice produce significantly less HO compared with wild type. Furthermore, HO, which forms initially in the ob/ob mouse, slowly resorbs over time. Overall, our study demonstrates that leptin deficiency may limit pathologic bone formation.
In vitro osteogenic differentiation
Our findings indicate that the osteogenic differentiation of adMSCs derived from burned wild-type mice is elevated compared with adMSCs derived from burned ob/ob mice. However, our finding that burn injury actually caused a decrease in the osteogenic differentiation of adMSCs from ob/ob mice was interesting, given previous reports that burn injury increases osteogenic differentiation of wild-type adMSCs [2]. This suggests that burn injury may actually decrease the osteogenic capacity of mesenchymal cells, which may be responsible for HO in ob/ob mice.
Diabetes, angiogenesis, and pathologic HO
Hif1α induces Vegfa expression, which directs vessel growth. We noted slightly increased Hif1α transcript and staining in the ob/ob mouse hind limb. We also found elevated Vegfa transcript and staining in the ob/ob hind limb, indicative of robust angiogenic signaling. Our findings were confirmed by the presence of robust angiogenesis at 7 weeks post-trauma in the ob/ob mouse. Therefore, decreased HO volume seen early in leptin-deficient mice is likely not related to decreased angiogenic signaling. Importantly, this difference was not present at baseline when comparing uninjured wild-type and ob/ob mice; rather, it appeared that there was a trend toward increased VEGF expression in wild-type mice. We did note that vessels in ob/ob mice were narrower and more disorganized. Whether this contributes to diminished HO is difficult to confirm.
Endochondral ossification and leptin deficiency
Normal bone healing occurs through endochondral ossification. During this process, undifferentiated MSCs condense at the site of future bone growth. These MSCs differentiate into chondrocytes, which hypertrophy and produce matrix proteins [23,24]. In this study, we chose to use MSCs derived from the inguinal fat pad to study differences in osteogenic potential between wild-type and ob/ob mice. We chose to study these cells as opposed to osteoblasts from wild-type and ob/ob mice because we are interested in understanding how leptin deficiency alters osteogenic potential of HO precursor cells and not of mature osteoblasts.
Osteoblasts and vessels invade the surrounding matrix, forming bone. SOX9 is a transcription factor known to play a central role in chondrogenic signaling. Incidentally, Hif1α also regulates SOX9 expression. Although Hif1α levels were elevated in ob/ob mice, and there was more downstream proangiogenic signaling (Vegfa) in ob/ob mice, SOX9 levels were actually diminished in the ob/ob mice at both 5 days and 3 weeks after injury. This suggests that Hif1α was unable to induce SOX9 expression with the same efficiency in ob/ob mice as in wild-type mice. Pentachrome staining at 3 weeks confirmed diminished cartilage production in the ob/ob mouse compared with wild type. These findings strongly suggest that the diminished chondrogenic differentiation contributes to diminished HO as early as 3 weeks post-trauma.
Next, we performed aniline blue and osteocalcin immunofluorescent staining of ob/ob and wild-type HO sections at 3 months after trauma. We found that aniline blue staining was much deeper in the wild-type mice for both HO and normal bone compared with the HO or normal bone of ob/ob mice. The fact that even normal bone appears to have diminished osteoid in the ob/ob mice compared with wild-type mice suggests that the effect of leptin deficiency on normal bone may be similar to its effect on developing HO (Fig. 4). Furthermore, these results are consistent with our pentachrome stain, which showed more red stain indicative of immature collagen deposition. In addition, HO from wild-type mice had more osteocalcin staining on immunofluorescence, suggesting more osteoblast presence within wild-type HO.
Increased osteoclast presence in ob/ob mice
Between 7 and 9 weeks, ob/ob mice showed decreased ectopic bone volume over time, in stark contrast to wild-type mice. Previous studies demonstrated that osteoclast function is upregulated in the setting of diabetes, prompting our interest in osteoclast presence in our model [12,13]. Osteoclast activity is one mechanism believed to contribute to osteoporosis in diabetic patients. We stained sections of HO in ob/ob and wild-type mice with TRAP to assess for osteoclast presence. We noted robust osteoclast presence at 3 weeks in ob/ob mice with persistent activity at 11 weeks postinjury. We noted no osteoclast presence in the HO of wild-type mice. This finding suggests a mechanism by which HO is resorbed in the ob/ob mouse model of diabetes.
Limitations
In this study, we use leptin-deficient ob/ob mice as a model for type 2 diabetes. Serum from ob/ob and wild-type mice in our study confirms that the ob/ob mice have significantly higher glucose and insulin levels, similar to type 2 diabetes. However, it is also possible that leptin deficiency itself is responsible for decreased HO in the ob/ob mouse. To study this, it may be required to determine whether leptin deficiency is sufficient to decrease HO in the absence of hyperglycemia or hyperinsulinemia. Previously, it has been shown that ob/ob mice without hyperglycemia have a very low rate of bony turnover.
Conclusions
We demonstrate that pathologic bone formation after trauma, or HO, is limited in the setting of diabetes. Our study using the ob/ob mouse model suggests that this may be secondary to impaired early SOX9 activation and chondrogenic differentiation, followed by late bone resorption by osteoclasts. Diabetic mouse models serve as a promising tool to study targets for the prevention or treatment of HO.
Footnotes
Acknowledgments
The authors thank the Department of Radiology at The University of Michigan for the use of The Center for Molecular Imaging and the Tumor Imaging Core, which are supported, in part, by the NIH grant P30 CA046592. B.L. received funding from NIH/NIGMS-K08GM109105-0, Plastic Surgery Foundation National Endowment Award, the Association for Academic Surgery Roslyn Award and the Research & Education Foundation Scholarship, American Association for the Surgery of Trauma Research & Education Foundation Scholarship, DOD: W81XWH-14-DMRDP-CRMRP-NMSIRA, and American Association of Plastic Surgery Research. S.A. funded by the NIH LRP and Coller Society. S.L. funded by HHMI fellowship.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.