
Abstract
Hematopoietic stem and progenitor cells (HPC) niche, consisting of HPC and their surrounding stromal components, is the fundamental unit for bone marrow (BM) tissue engineering. Previously, mouse BM-derived cell complexes with HPC niche unit properties called “niche-like units” were isolated and characterized. This study was aimed to evaluate the possibility of bioengineering marrow tissue in heterotypic sites using niche-like units in combination with three-dimensional scaffolds. BM niche-like units were isolated from GFP-transgenic C57BL/6 mice and seeded on electrospun poly (L-lactide) nanofiber scaffolds, which were then roll-folded and aseptically implanted into the peritoneal cavity of irradiated wild-type mice. One month after implantation, donor-derived cells were detected in peripheral blood of the recipients and contributed to restoration of all blood lineages. The transplanted bioengineered tissue histologically resembled native BM structure and was connected to the mouse systemic circulation. Long-term self-renewal was confirmed by serial transplantation into tertiary recipients. In conclusion, this study establishes a novel system for BM tissue engineering, which can be used to improve the HPC transplantation outcomes especially in cases where HPC niche is damaged and also as an in vivo model to test the effects of different factors on hematopoiesis.
Introduction
H
In previous studies we isolated and characterized unique cellular complexes from bone marrow called BM HPC niche-like units based on size fractionation [18,22]. In culture experiments confirmed BM cellular complexes were the origin of both BM-mesenchymal stem cells (BM-MSCs) and various hematopoietic lineages. Transplantation of mouse HPC niche-like units into irradiated recipients resulted in long-term multilineage reconstitution [22]. We showed that a bulk of these niche-like units is disposed as medical waste during the clinical BM transplantation [18]. In this study we aimed to isolate the niche-like units from murine BM and then culture in poly (L-lactide acid) (PLLA) nanofibrous scaffolds as a promising substrate, which is clinically approved and support the stromal cell growth and differentiation by providing structure closely mimic of natural ECM [23,24]. This three-dimensional (3D) scaffold-based culture system naturally consists of major components of HPC niche, which support HPC functions and could be intraperitoneally transplanted into irradiated mice.
Materials and Methods
Nanofiber scaffold fabrication by electrospinning
Electrospinning was used to produce nanofibrous scaffolds as reported previously [25]. Briefly, a solution of PLLA (Sigma-Aldrich, St. Louis, MO) in dichloromethane (12% w/v; Merck, Darmstadt, Germany) was placed in a 5 mL syringe and then pumped with a rate of 1 mL/h into the following extension tube connected to a 21-gauge needle. A high voltage (20 kV) was applied to the distance between the tip of the needle and a steel collector to obtain ultrafine fibers. The obtained fibers were collected and placed in vacuum to remove the residual solvent. To enhance the surface hydrophilicity of fibers and improvement of the cell adhesion and proliferation, scaffolds surface were modified using oxygen (O2)-plasma treatment [26,27]. The hydrophilicity of scaffolds was determined using the measurement of contact angle of electrospun materials by the sessile drop method with a G10 contact angle goniometer (Kruss, Hamburg, Germany) at room temperature. A water droplet was placed on the scaffold surface and the contact angle measured after 10 s. Angle measurement was repeated three times. To evaluate the mechanical properties of scaffolds, PLLA mats were cut into 10 × 60 × 0.2 mm rectangular samples and assessed using a universal testing machine (Galdabini, Cardano al Campo, Italy). Briefly, tensile testing was measured under 50 mm/min generator (44 GHz; Diener Electronics, Ebhausen, Germany). Using 0.4 mbar pressure, oxygen was introduced into the reaction chamber followed by 5 min glow discharge. The tensile modulus of the scaffolds was measured from the linear portion of the stress–strain curve.
Assessment of stem cell and scaffold interaction and in vitro cell proliferation
The biocompatibility of the synthesized scaffolds was assessed using the proliferation capacity of murine BM-derived MSC. MSC were collected as previously described [18]. Scaffold disks were prepared (5 mm diameter ×1 mm thickness) and inserted in 24-well plates. Scaffolds were sterilized with 70% ethanol overnight and then UV irradiated in a laminar flow hood for 20 min per side. Sterilized samples were incubated in standard growth medium; Dulbecco's modified Eagle medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 1% penicillin, and 1% streptomycin (all from Gibco-BRL, Grand Island, NY) overnight. Then, MSC were seeded at a density of 5 × 103/cm2 on sterilized scaffolds and cultured in DMEM supplemented with 10% FBS under standard culture conditions. To measure cell viability, after 1, 3, 5, and 7 days of culture, medium was removed and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Sigma, Munich, Germany) solution was added at a final concentration of 5 mg/mL. After 4 h incubation, MTT solution was removed and the formed formazan crystals were dissolved in DMSO (Merck). The optical density was evaluated at 570 nm in reference to 630 nm with spectrophotometer (BioTek, Winooski, VT). All experiments were repeated three times. To investigate scaffolds surface morphology and cell's interactions, acellular and seeded scaffolds (as described above) were analyzed by scanning electron microscopy (SEM) as reported previously [25]. Briefly, scaffolds were fixed in 2.5% glutaraldehyde for 2 h at 4°C. After dehydrating using a gradient of ethanol, scaffolds were dried and coated with gold, and finally observed under SEM (VEGA II; TESCAN, Brno, Czech Republic).
Animals
Eight to 10-week-old female C57BL/6 (Pasteur Institute, Tehran, Iran) and syngeneic GFP-transgenic male mice (kindly provided by Dr. M. Okabe Osaka University, Osaka, Japan) were used in BM tissue engineering studies. All Animal Care and experiments were according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals and approved by the Tehran University of Medical Sciences Animal Ethics Committee, Tehran, Iran.
Isolation of BM HPC niche-like units
Male GFP transgenic mice were sacrificed by cervical dislocation. Both tibia and femurs were removed and the marrow was exposed by distal end removal. Niche-like units were collected as described previously [22]. Briefly, the bones were inserted into adapted centrifuge tubes and centrifuged for 1 min at 600 g. The cell pellets were suspended in heparinized phosphate buffered saline (PBS; Gibco-BRL), and filtered through a 20 μm cell strainer (a double layered 40 μm nylon mesh; BD Bioscience, San Jose, CA). The niche-like units were collected from the surface of the strainers and washed with PBS.
Construction and implantation of engineered BM tissue
Twenty-four hours before implantation, the recipient mice were exposed to a single dose of 8.2 Gy total body irradiation. This dose was chosen based on preliminary experiments as appropriate dose for BM ablation [22]. The niche-like units derived from two femurs and tibias of each donor mouse were directly seeded on a sterile 1 × 1.5 cm PLLA scaffold mat. The seeded scaffolds were then roll-folded and aseptically implanted into the peritoneal cavity of the irradiated C57BL/6 female wild-type mice under anesthetic conditions (n = 12). For control group, the scaffolds were implanted without cells (n = 12).
To assess the self-renewal potential of transplanted cells, serial transplantation was performed. In brief, at day 45 upon transplantation the engineered bone marrow tissue (EBMT) from first transplanted mice were harvested and transplanted into the peritoneal cavity of second irradiated C57BL/6 mice. After 45 days, the EBMT from second transplanted mice were harvested and transplanted into the peritoneal cavity of the third irradiated C57BL/6 mice. Next, the EBMT from tertiary transplant were harvested and evaluated.
Flow cytometry, histological examination, and multilineage differentiation
These experiments were performed as described previously [22,28].
Data presentation
All data are presented as mean ± standard deviation. All in vitro experiments were repeated at least three times.
Results
The fabricated PLLA nanofiber scaffolds support MSC proliferation
SEM analysis confirmed producing fibers with a porous and uniform morphological appearance and an average fiber diameter of 879 ± 274 nm (Fig. 1A). The contact angle of PLLA scaffolds was demonstrated to be 115° ± 9.3 before plasma modification while decreased to zero after plasma treatment. The tensile strength and elongation at break were 1.74 ± 0.37 MPa and 52% ± 7.5% respectively (Fig. 1B). To assess the biocompatibility of electrospun scaffolds MSC were seeded on scaffolds. SEM observation showed MSC were exhibited a typical fattened polygonal morphology with strong attachment on the surface (Fig. 1C). In addition, MTT assay was performed and described the gradual increase in MSC cell survival after 7 days in vitro culture (Fig. 1D).
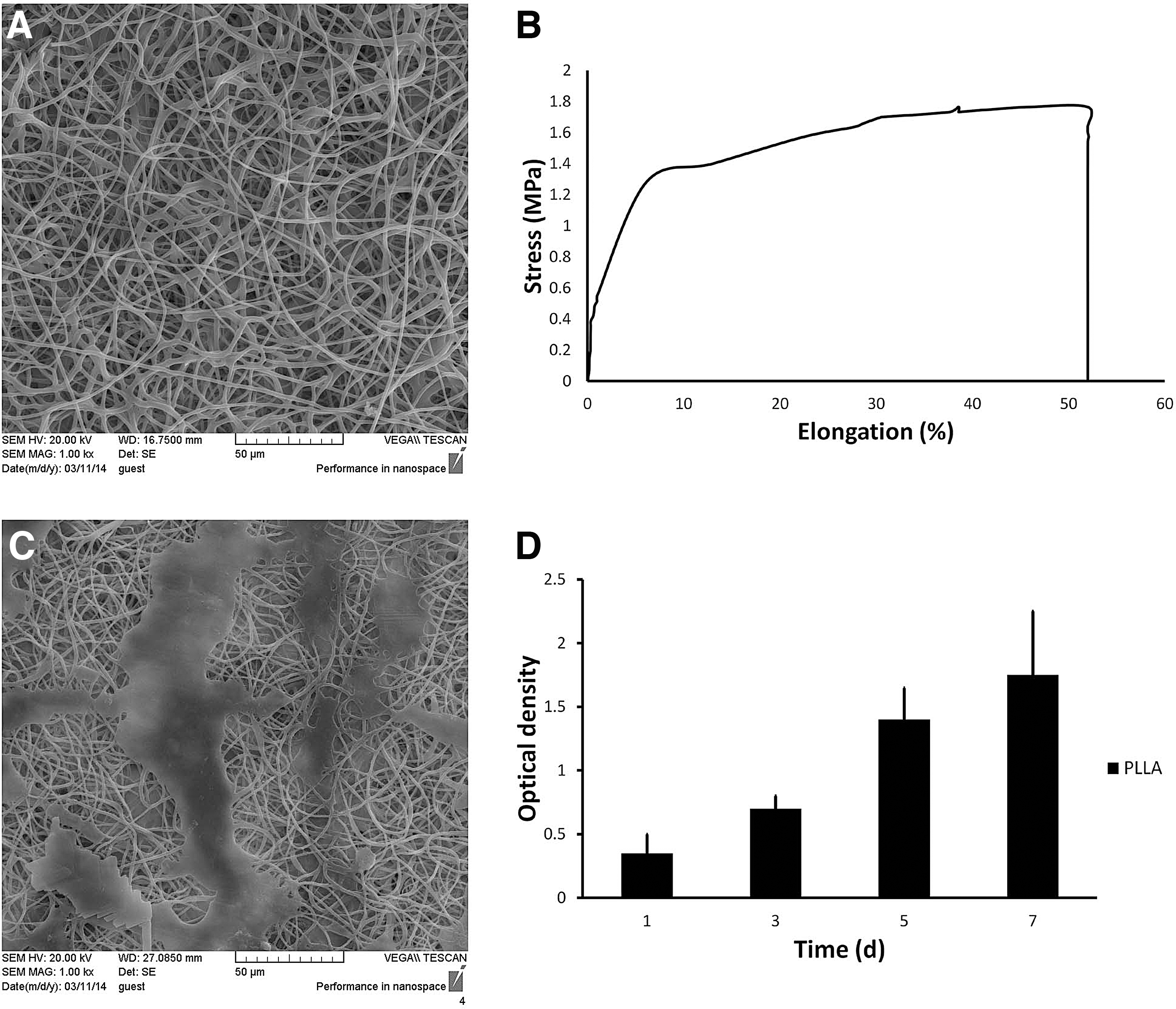
Characterization of fabricated PLLA scaffolds. Scanning electron microscopy revealed the uniform porous structure of the scaffold
The EBMT contributes in reconstitution of different blood lineages in the irradiated recipient mice
HPC niche-like units were isolated from the BM of GFP-transgenic mice using a size filtration method and were seeded on PLLA scaffolds, which were then roll-filled and transplanted into peritoneal cavity of irradiated syngeneic mice. Mice were then sacrificed for the assessment of blood lineages reconstitution and histopathologic examination (Fig. 2).

Schematic design for generation of EBMT in a heterotopic site. Hematopoietic stem and progenitor cells niche-like units were isolated from GFP-transgenic mice using a size filtration method and seeded on PLLA electrospun nanofiber scaffolds. These scaffolds were then roll-folded and transplanted to irradiated wild-type syngeneic mice. After 1 month, the recipient mice were sacrificed and the bioengineered tissue was subjected to histopathological examinations, and the contribution of transplanted cells in reconstitution of hematopietic system was assessed by cytofluorometric analysis of peripheral blood. BM, bone marrow; EBMT, engineered bone marrow tissues.
One month after transplantation of BM HPC niche-like units, blood samples were obtained from six mice for cytofluorimetric analysis, which revealed that 21% ± 7% of total nucleated peripheral blood cells were GFP-positive and therefore donor-derived (Fig. 3A). Also, staining for lineage-specific markers showed that 24% ± 5% of CD3+ (T lymphocytes marker), 17% ± 3% of CD11b (Mac-1, macrophage marker), 19% ± 4% Gr1+ (myeloid marker), and 48% ± 6% of B220 (B lymphocyte marker) were derived from the transplanted EBMT (Fig. 3B).
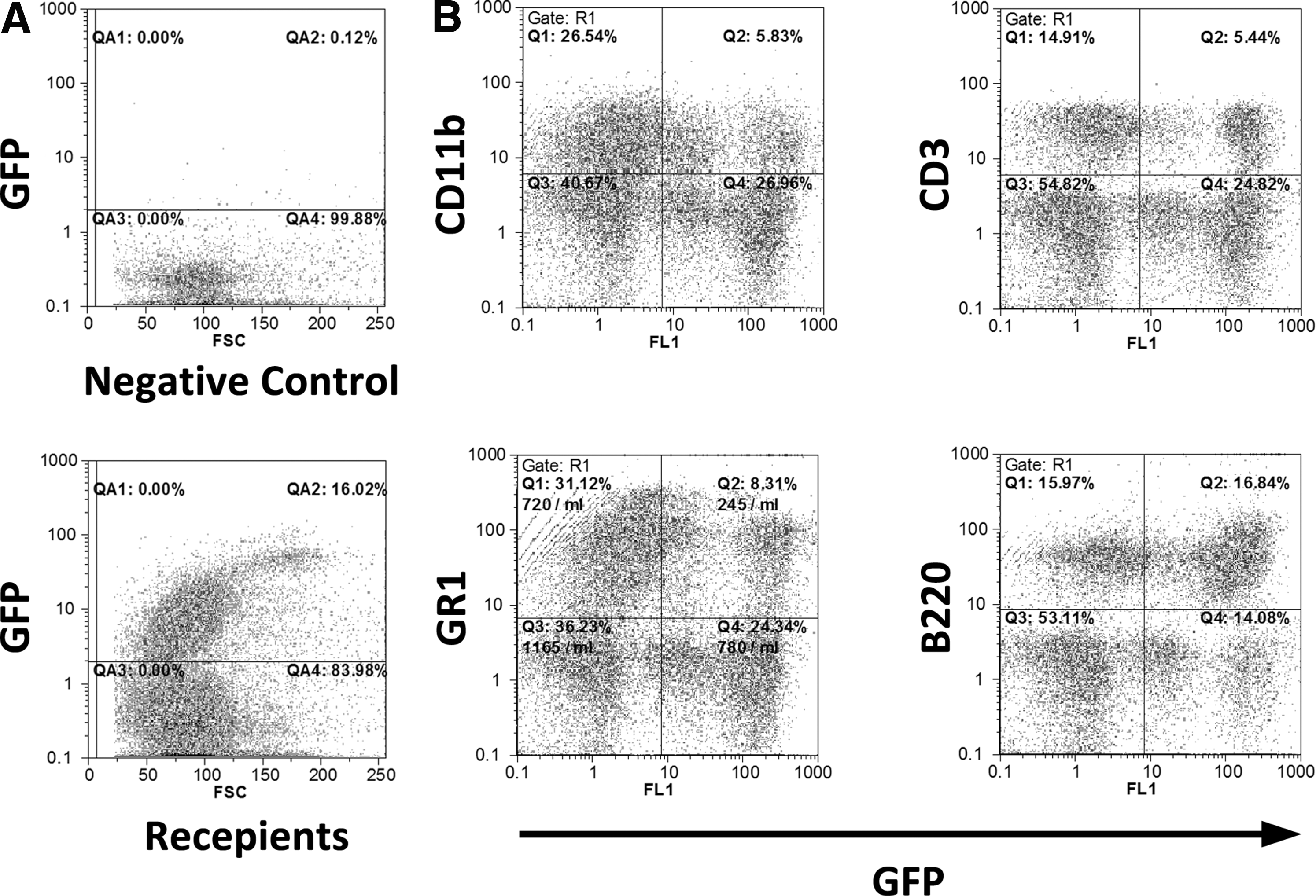
The transplanted EBMT creates all blood lineages. Niche-like units harvested from GFP-transgenic mice were seeded on scaffolds and transplanted to wild-type syngeneic irradiated mice. Representative cytofuorometric graphs for the assessments of GFP-positive cells in peripheral blood of recipients are shown. One month after transplantation, 21% ± 7% of nucleated peripheral blood cells were GFP-positive
The transplanted EBMT resembles native BM structure
After 1 month of transplantation, the recipient mice were sacrificed and peritoneal cavities were explored. Grossly, EBMT resembles a solid inner-brown cylinder surrounded and penetrated with the omentum vasculature (Fig. 4A, B). In histologic examinations, outer and inner layers were evident in EBMT. The outer layer was mainly consisted of fascia, adipose tissue, with capillaries and dispersed giant multinucleated histiocytes (Fig. 4C, D). The inner layer was hyper-cellular and obviously resembled native BM with sinusoidal blood networks, hematopoietic islands, and adipose tissue (Fig. 4E). These hematopoietic islands consisting of megakaryocytes and myeloid cells signify the formation and activation of hematopoietic tissue (Fig. 4F). Interestingly, in the inner layer the formation of bone trabecula was also noted as assessed by hematoxylin and eosin and Von Kossa staining (Fig. 4G, H, respectively). According to Masson's trichrom staining, fibrosis was detected neither in the inner nor in the outer layer and only a thin sheet of fibrotic tissue was surrounding the folded scaffolds (Fig. 4I). In addition, EBMT sections were subjected to immunohistochemistry for GFP. Immunostaining described presence of donor-derived cells in the newly formed tissue in recipient mice (Fig. 4J, K). Remarkably, after 5 months the trabecular bones completely disappeared and the active hematopoietic marrow was mainly replaced by adipose tissue with only a minority of cells remaining in the central parts (Fig. 4L).
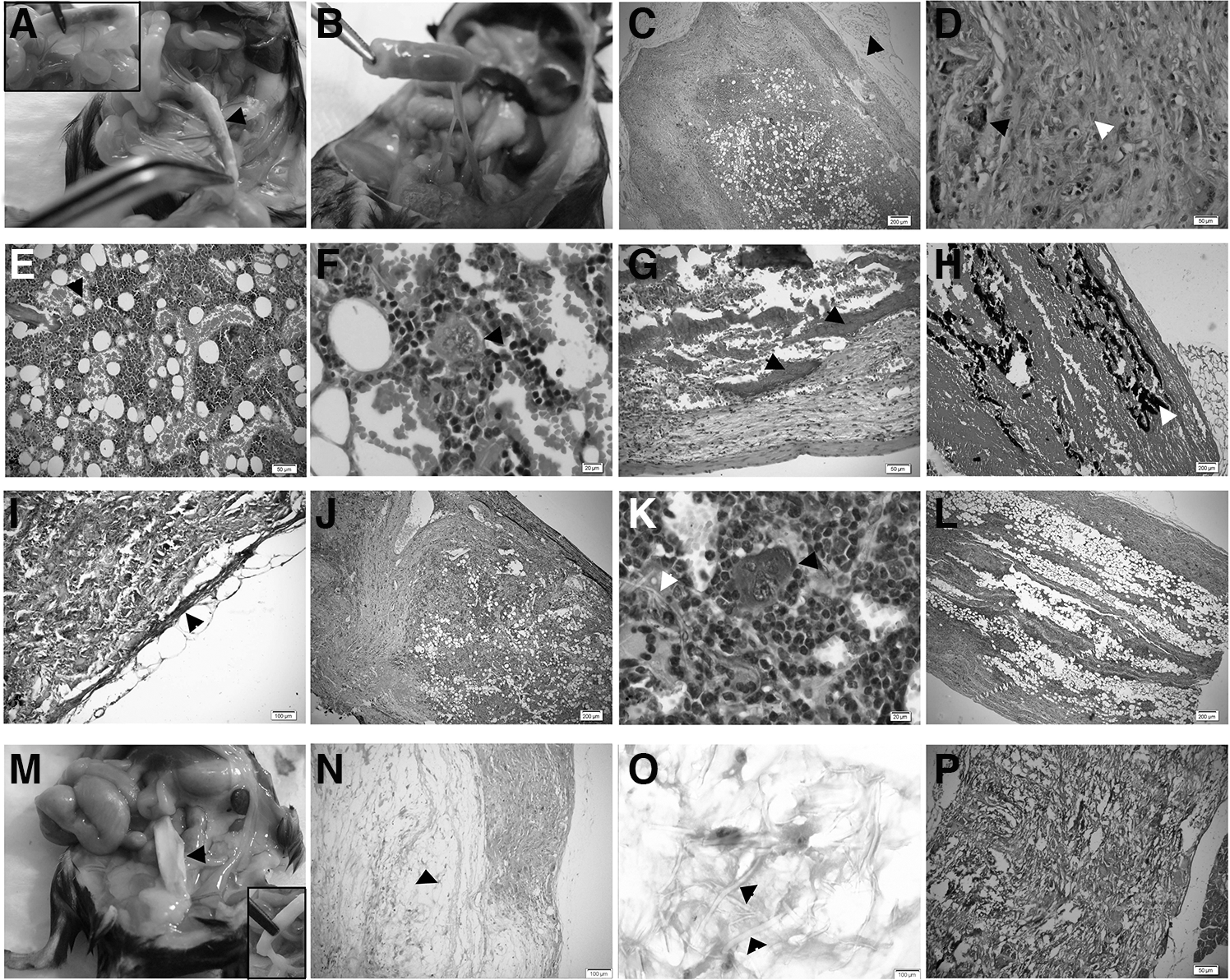
The bioengineered BM generates a heterotopic blood organ with marrow-like structure in the recipients.
The scaffold in control mice, receiving scaffolds without HPC niche-like units, showed a hollow white appearance connected to omentum with fat and fascia at both ends without significant connection to the recipient vasculature at 1 month after transplantation (Fig. 4M). On microscopic examination, the empty scaffolds also exhibited a two layer structure with mucouid degeneration and very low cell density in the central layer. The outer layer contained few cells, some of which were histiocytes and some inflammatory cells (Fig. 4N, O). The fibrous network of the scaffolds was clearly visible throughout (Fig. 4P) and there was no evidence of hematopoiesis. Also, trabecular bones were not detected in alizarin red or Von Kossa staining (data not shown).
Serial transplantation experiment was performed to assess the self-renewal capacity of aggregates on EBMT (Supplementary Fig. S1A; Supplementary Data are available online at
Discussion
In this study, we have shown that EBMT composed of BM HPC niche-like units and PLLA scaffolds generates a tissue that is structurally very similar to native BM and contributes in generation of different blood cell lineages in the irradiated recipients. This bioengineered tissue also stimulates the growth of peritoneal vessels, which penetrate to the newly formed tissue and become interconnected with sinusoidal vessels. This is probably due to the angiogenic factors that may secrete by native niche-like units as it was not detected in the control mice receiving scaffolds alone. It was previously shown, forming these new vessels and connection to systemic circulation is pivotal for tissue engineering strategies [29].
Histologic examination showed formation of hematopoietic islands, trabecular bones, and the network of sinusoidal vessels in the EBMT. In addition, serial transplantation experiments demonstrated that hematopoiesis could be continuously induced upon secondary and tertiary transplantation in recipient mice where recipient's hematopoiesis was impaired. However, in long-term and upon transplantation into irradiated mice the transplanted tissues were replaced by fibrotic and fat tissue, this could be explained by contribution of recipient hematopoietic system in hematopoiesis, which could efficiently compensate the hematopoiesis needs.
Previous results showed that transplantation of niche-like units without scaffolds produces a lymphoid organ [22] while here implantation with nanofibrous scaffolds described generation of such a well-established marrow-like structures and implies the crucial role of PLLA scaffolds. Indeed, the fabricated scaffold provides a suitable matrix for attachment, proliferation, and appropriate differentiation of seeded cell complexes. It has been described that PLLA scaffold has osteoinductive and osteoconductive properties [23,24,30 –32], which can induce MSC that were located inside the niche-like units into osteogenic lineage and form a bone layer, which is similar to trabecular bone layer in vivo and is essential for normal hematopoiesis. Furthermore, it is well documented that osteoblasts directly or indirectly can regulate HSC quiescence and activation in vivo [33] or in 3D in vitro model [34].
An advantage of current strategy of BM tissue engineering is that HPC were not harvested as single cells from the donor but were in association with their natural microenvironment, which more accurately mimic the 3D complexity of HSC microenvironment. Together, this study proposes an effective humanized mouse model, which allows activation of HSC in a 3D model after transplantation in an almost native structure. The current system also could be used to test compound efficacy in vivo, and investigate the impact of different factors on hematopoiesis.
Footnotes
Acknowledgment
This study was supported by a grant from SABZ Biomedicals Science-Based Company, Tehran, Iran.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.