
Abstract
Zebrafish is gaining increased popularity as a model organism to study stem cell biology. It also is widely used as model system to visualize human leukemic stem cells. However, xenotransplantation of primary human stem/progenitor cells has not been described. Here, we use casper pigmentation mutant fish that are transparent crossed to fli-GFP transgenic fish as recipients of red labeled human CD34+ cells. We have investigated various conditions and protocols with the aim to monitor and visualize the fate of transplanted human CD34+ cells. We here report successful use of casper mutant zebrafish embryos for the direct monitoring of human hematopoietic stem cell transplantation, differentiation, and trafficking in vivo.
Introduction
H
To track the in vivo fate of the transplanted cells, advanced techniques such as positron emission tomography, single photon emission computed tomography, magnetic resonance imaging, and bioluminescence imaging mostly with luciferase, have been used [5]. However, such techniques are invasive, have limited sensitivity, sometimes require anesthesia and surgery, and are difficult to perform over the time frames required to study HSC biology. However, real-time imaging of transplanted cells in vivo has become possible using zebrafish as a model system.
The use of zebrafish (Danio rerio) as a model has gained much popularity, in particular that of casper multiple pigment mutant that remains transparent throughout adulthood allowing easy monitoring of developmental processes and cell trafficking in vivo [6]. In using the zebrafish model, several potential issues should be considered. First, the different maintenance temperatures (zebrafish usually at 28°, human cells at 37°), difference in organ and vessel size, lack of available antibodies, and lack of obvious transplantation routes. Some of these hurdles have been overcome using human tumor cells and cell lines, for instance fish can be kept at 33°–34° [7] and the lack of antibodies can be overcome by genetic tools or for fish blood cells by fluorescence-activated cell sorting (FACS) scatter profiles [8,9]. Obvious advantages such as ease of visualizing transplanted cells in situ, permeability of fish to small molecules added to the aquarium water, and the relatively low number of cells required for transplantation, compared to typical murine experiments, make efforts to generate “humanized” fish worthwhile.
Moreover, because the genes and signaling pathways that control hematopoiesis are highly conserved between zebrafish and mammals [10,11] and the zebrafish adaptive immune system is not functional within the first 4 weeks postfertilization, we investigated whether zebrafish embryos could be used as a xenotransplantation model.
We here report successful use of casper mutant zebrafish embryos for the direct monitoring of human HSCT, differentiation, and trafficking in vivo.
Materials and Methods
Human CD34+ cells
Human CD34+ cells were isolated from umbilical cord blood (UCB) samples. Mononuclear cells of UCB were isolated by Ficoll [Leiden University Medical Center (LUMC) pharmacy] density centrifugation, washed, and stored in liquid nitrogen until further use. Single UCB units (HSCT) or 5–10 combined UCB units (CD34+ transplantations) were selected, thawed, and selected for CD34+ progenitors using the CD34 Microbead Kit (Miltenyi Biotec GmbH) according to the manufacturer's protocol. Purity of the CD34 selection was verified by flowcytometry and exceeded 95%. Experimental procedures were approved by the Ethical Committee on Animal Experiments of the LUMC. The UCB samples used in this study were collected at the Diakonessenhuis Hospital (Leiden, the Netherlands) after informed consent was given by the parents. The Joint Medical Ethical Committee of LUMC and Diakonessenhuis Hospital approved this procedure (P14.078).
Zebrafish husbandry
Zebrafish were handled in compliance with the local animal welfare regulations and maintained according to standard protocols (
Zebrafish transplantation
U937 human monocytic leukemic cells or cord blood-derived CD34+ cells were intravenously injected in the duct of Cuvier or yolk sac (posterior cardinal vein) in 48 h-old Casper Fli-GFP zebrafish. The fluorescently labeled HSCs were followed using stereo and confocal microscopy. Upon transplantation of human cells, the fish were transferred from aquaria at 28°C–34°C. Retro-orbital transplantation was performed as described by Zon and coworkers [12].
Flow cytometry
Cells were incubated with PKH26 (No. PKH26GL; Sigma-Aldrich) for 2 min and washed extensively following directions of the manufacturer. For CD34 and CD38 staining, erythrocytes in the cord blood samples were lysed using an isotonic NH4Cl buffer (LUMC apothecary), after which the cells were stained with the indicated antibodies (anti-CD34, anti-CD38; Becton Dickinson) in phosphate-buffered saline (PBS)/0.2% bovine serum albumin/0.1% sodium azide buffer for 30 min on ice. Cells were subsequently analyzed using a FACS Canto II (Becton Dickinson). For stainings of human cells with antibodies, cells were stained with antibodies against the human markers CD3, CD14, CD34, CD38, and CD45 using standard protocols, as described [13] using intravenous immunoglobulins to block Fc receptors on myeloid cells. For adult fish, fish were incubated for 10 min in 0.5% warm trypsin in unconditioned medium and single cell suspensions were prepared by passing the fish over a cell strainer.
Stromal cocultures
Zebrafish stromal cells were generated by FACS sorting of Fli+ endothelial cells (Fli-GFP+) that were cultured in 24-well plates in RPMI1640 medium supplemented with 10% fetal calf serum. These cells or conditioned medium thereof were used in a transwell system to study migration of human cells toward zebrafish stromal cells. Transwell migration assays were performed in Transwells of 6.5 mm diameter with 5 μm pore size polycarbonate filters (Costar) coated with 10 μg/mL fibronectin (Sigma-Aldrich) in duplicate. Human CXCL12 was used as positive control.
Results
Human hematopoietic stem/progenitor cells survive up to 6 days after transplantation in zebrafish embryos
The adaptive immune system of the zebrafish is not functional within the first 4 weeks postfertilization. We have used this immune tolerant window to inject human 50–500 CD34+ UCB-derived hematopoietic stem/progenitor cells (HSPC) intravenously in of 48 h-old embryos. Labeling of the HSPC with the live fluorescent dye PKH26 allowed live imaging of the transplanted cells via fluorescent (confocal) microscopy. In combination with the use of transparent casper zebrafish that express GFP in their endothelial cells (Fli:GFP casper) tissue distribution of the fluorescent HSPC could be determined.
Experiments in which CD34+ cells or U937 human erythromyeloid leukemia cells were injected in the yolk sac never led to detectable human cells in the embryo proper, as cells always remained trapped in the remnants of the yolk sac (Fig. 1). However, transplantation was successful (ie, cells left the site of injection) by injecting into the duct of Cuvier, a temporary vessel at the rostral side of the embryo also called common cardinal vein (CCV), which returns blood to the heart. Injection at this site led to reproducible and ready detection of red fluorescent human CD34+ HSPC that were visible within the cardiac tissue and in the tissue surrounding the injection site directly. Using live confocal imaging we observed that the human cells actively extravagated the blood vessels in the caudal hematopoietic region of the tail (Supplementary Movie S1; Supplementary Data are available online at

Different injection routes in zebrafish embryos lead to different engraftment. Injection of PKH26-labeled human cells in the yolk sac lead to entrapment and lack of migration into the circulation of the embryos, while injection into the duct of Cuvier lead to migration into the circulation of the fish embryo. Representative pictures are shown. For yolk sac injection-distribution experiments, over 100 embryos were analyzed in three different experiments. In the Duct of Cuvier distribution experiments, over 65 embryos were analyzed in four different experiments. Color images available online at
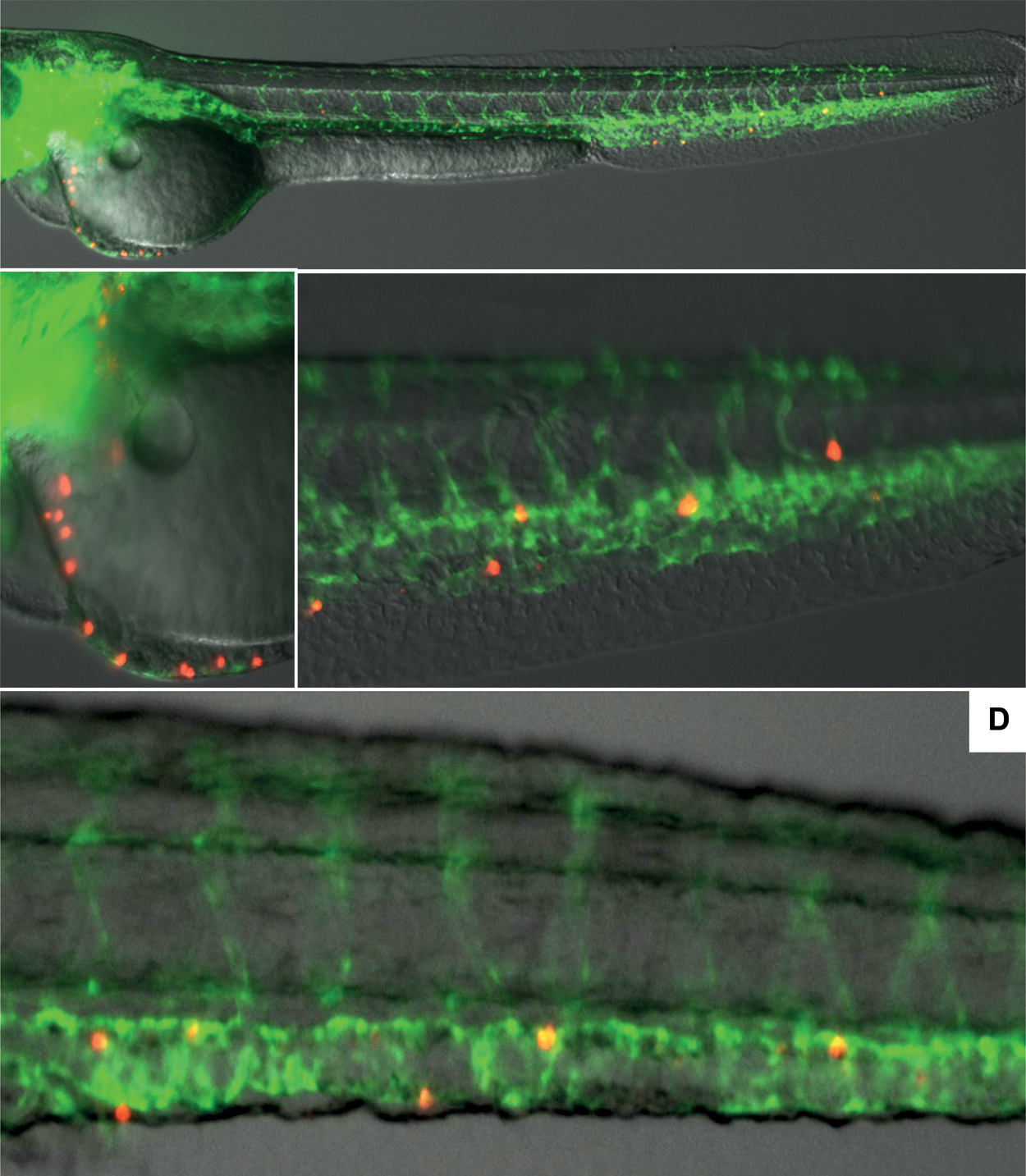
Human HSPC survive in zebrafish embryos up to 6 days after injection. Human HSPC labeled with pkh26 (red) are injected in the Duct of Cuvier of 48 h transparent Casper fish that express Fli-GFP in their endothelial cells (green). Fluorescence is visualized using a Leica fluorescent microscope. Representative pictures are shown as in Fig. 1 but now for human CD34+ cells. For yolk sac injection-distribution experiments, over 100 embryos were analyzed in three different experiments. For duct of Cuvier experiments, over 65 embryos were analyzed in four different experiments. Panel
Human HSPC respond to zebrafish stromal cell-derived factors
The chemokine CXCL12 and its receptor CXCR4 represent probably the best-studied migration pathway for HSC in vertebrates. CXCL12 induces polarization of CXCR4-expressing HSC leading to directed trafficking of the responding HSC toward CXCL12-expressing tissues.
We therefore examined the chemoattraction of human HSPC toward zebrafish stromal and endothelial cell-derived factors by conditioning medium by overnight culture and placing this medium in the lower compartment of a Transwell. Fig. 3 shows that zebrafish cell-derived factors attract human HSPC when compared to unconditioned medium. However, the addition of AMD3100, which was used to block CXCR4-mediated interactions, did not alter the zebrafish-cell-derived factor-mediated chemoattraction of HSPC, whereas it prevented the attraction toward human CXCL12 almost completely. Preliminary experiments using the Rho A kinase inhibitor Y27632 also show no differences in transwell migration, as do experiments after transplantation in vivo into Fli-GFP transgenic fish that lack functional CXCL12a. This indicates that either factors other than CXCL12 mediate transmigration of human HSCs or that the drugs cannot efficiently interfere with the fish derived signals that target CXCR4. Nevertheless, conserved fish signals are capable of inducing migration of human HSCs, a prerequisite for using zebrafish as a xenotransplantation model for human CD34+ cells.
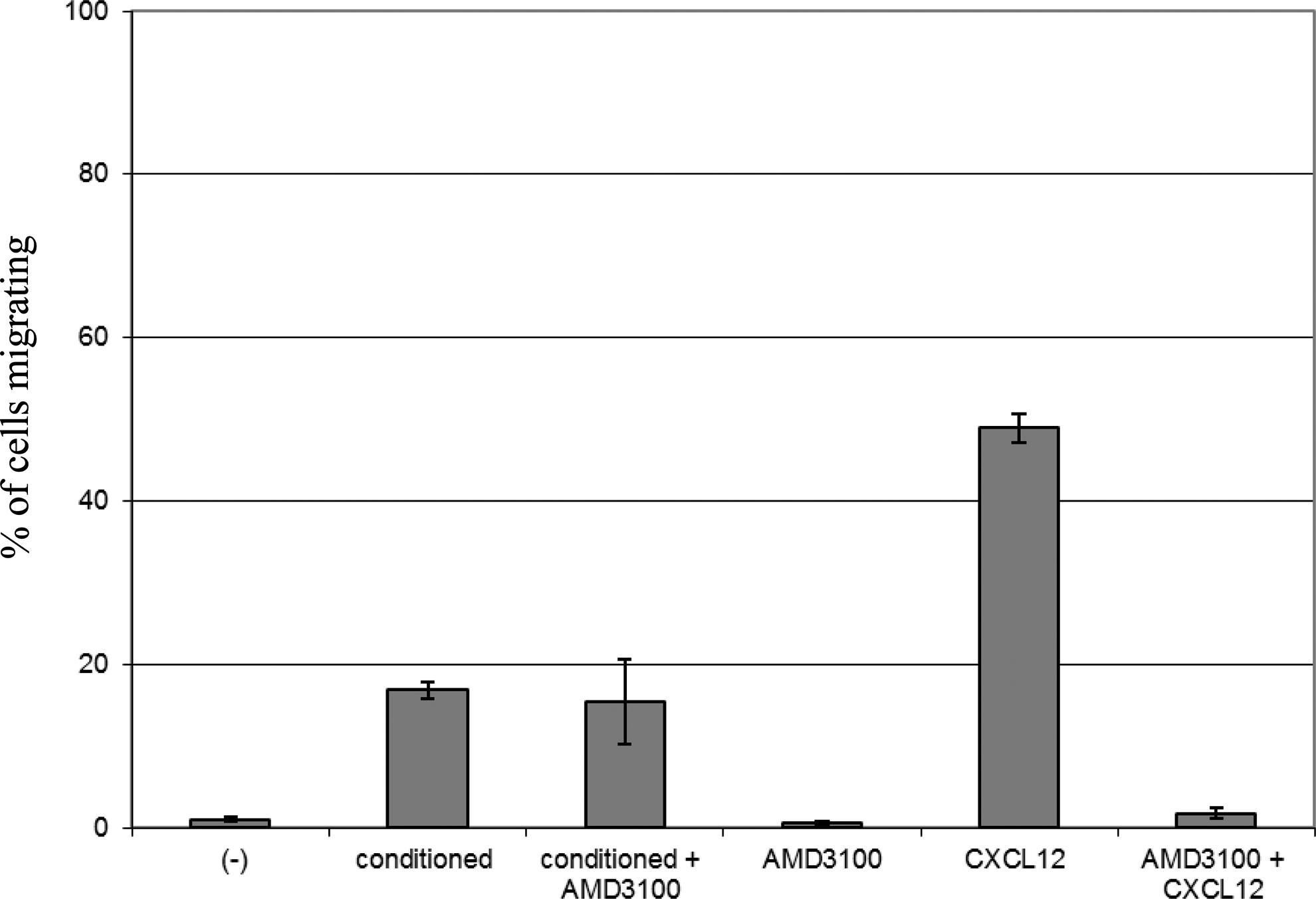
Human HSPC respond to zebrafish stromal cell-derived factors. Human HSPC migrate are added to a 5 μm pore size insert of a Transwell and allowed to migrate to the lower compartment of the Transwell, which contains migration medium or migration medium condition with zebrafish stromal cell-derived factors. After 3 h cells are harvested from the lower compartment and counted. The percentage of migrated cells is calculated as compared to the number of cells that is added to the insert. Data from 12 embryos each in two different experiments were analyzed.
Human HSPC transplanted into zebrafish differentiate along the myeloid lineage
We monitored the differentiation status of injected human cells by optimizing flow cytometric analysis of single cell suspensions prepared from whole zebrafish embryos at different time points postinjection. In the current experimental setup it is not possible to exactly define the number of HSPC that are injected in each embryo. Based on the injection volume and velocity used, the injected cell number is estimated at 50–500 cells per embryo. Using dual expression of the fluorescent label and human CD45 cell surface protein, we have been able to detect injected human cells up to 6 days postinjection using single cell suspensions from 10 pooled embryos (Fig. 4, histogram). Very few of these human cells maintained the expression of surface CD34 (Fig. 3, left dot plots) and most human cells were of monocytic/myeloid origin based on the expression of cell surface CD14 (Fig. 4, right dot plots). Expression of the T-lymphocyte-associated molecule CD3 was not observed (Fig. 4, middle dot plots).
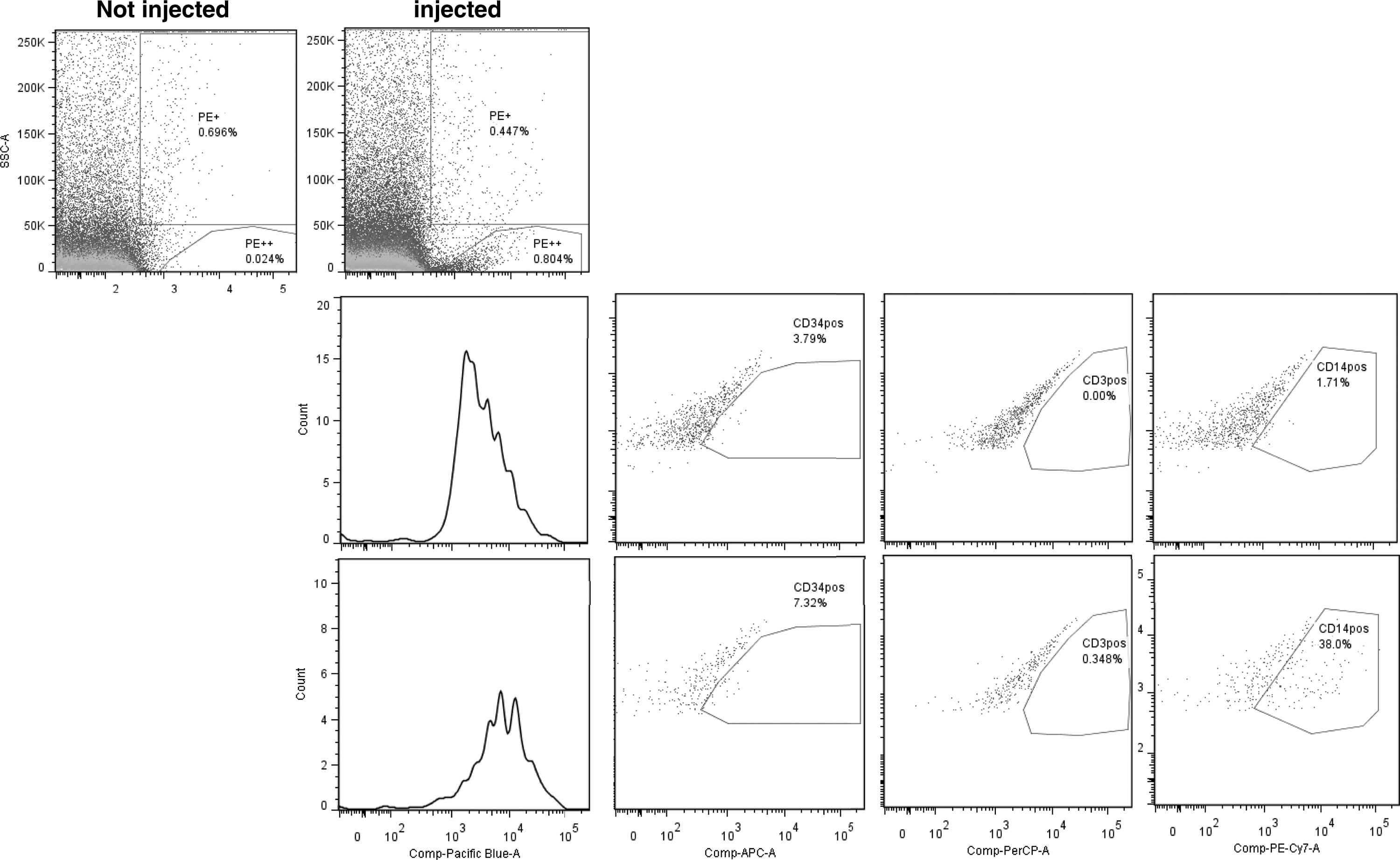
Human HSPC transplanted into zebrafish differentiate along the myeloid lineage. Zebrafish transplanted with human pkh26-labeled HSPC were sacrificed 6 days postinjection. Single cell suspensions were prepared and stained with different antihuman-specific conjugated antibodies. pkh26 cells were detected in the PE-channel (left upper dot plot) and the human origin of these cells was established by the expression of human CD45 (histogram and dot plots lower row). Most human cells expressed CD14 (right dot plot lower row). Representative data from two independent experiments are shown, harvested either after 6 days in the first experiment (n = 8) or after 2 (n = 4) and 6 days (n = 4) in the second experiment.
Human HSPC can be injected in adult zebrafish using retro-orbital injection
The results described above indicate that human CD34+ cells can be transplanted, survive, migrate, and differentiate when injected into zebra fish embryos. However, for a successful xenograft model adult fish would be required, to allow follow-up for extended periods of time. Ideally in analogy with xenografted mice, the host is immune deficient and indeed fish to fish transplantation have been conducted in c-myb bloodless fish [14] using whole kidney marrow. The use of adult fish immediately necessitates the question of the delivery route for the human cells. Having observed that certain injection routes (eg, yolk sac) are unsuccessful even in embryos, we focused on retro-orbital injections in adult wild-type fish as a first step to investigate whether xenotransplantation would be feasible.
First, we tested whether human cells can be intravenously transferred by retro-orbital injection in adult zebrafish. Therefore, human leukemic myeloid U937 cells were labeled with PKH-26 and injected in anesthetized 9-week-old adult c-myb heterogeneous fish. Using flow cytometry we were able to show that labeled U937 cells can be detected in single cell suspensions of the head region, but not in single cell suspensions of the tail region, when measured 2 h after injection. However, labeled U937 cells were easily detected in both regions when measured 24 h after injection (Fig. 5). In addition, adult fish transplanted with PBS, head kidney cells or human HSPC all survived (at least up to 13 weeks of age), regardless of the transplant.

Human HSPC can be injected into adult zebrafish using retro-orbital injection.
Discussion
To the best of or knowledge we here report for the first time successful use of zebrafish for xenotransplantation of primary human stem/progenitor cells. A large number of articles have described use of leukemic or other cancer cells for xenotransplantation in zebrafish (reviewed in Konantz et al. [7]) following the seminal work by Lee et al. who showed transplantation of human melanoma into zebra fish [15]. Human cells have mostly been transplanted, by injection into the yolk sac. Pruvot et al. have used this approach fairly successfully for human leukemic cells [16], but similar to our observation injection in the yolk sac of normal CD34+ cells only resulted in detection of human cells for 1 day. The invasive nature of cancer cells may have contributed to successful transplantation in these and other reports [17,18], as in our hands nontransformed primary cells remained trapped at the boundary of yolk to embryo proper and never entered the embryonic circulation. Key to our successful approach appears to be the injection into the duct of Cuvier, the CCV that leads to the hearts sinus venosus.
We here show that injected human HSPC can be easily monitored in live transparent zebrafish embryos when labeled with a red fluorescent dye. HSPC actively crossed the endothelial layer of blood vessels shortly after intravenous injection.
Based on the Transwell assay, the transplanted human HSPC appear to respond to zebrafish-derived chemotactic cues. Human cells were detected up to 6 days postinjection, either by live imaging or flow cytometric analysis, which indicates responsiveness to zebrafish-derived cues supporting human cell survival. The nature of such signals wants further investigation. Moreover, flow cytometric analysis using human-specific antibodies showed differentiation of injected HSPC toward the myeloid lineage, suggesting zebrafish-to-human cross-reactivity of zebrafish-derived growth factors. That human cells can respond to developmental clues in zebrafish embryos is not unprecedented. Orlova et al. showed that iPSC-derived human pericytes and endothelial cells integrated into developing vasculature as xenografts in zebrafish [19].
For a successful xenotransplantation model, akin to immune-deficient mice, human HSPC should be transferred into adult immune permissive zebrafish lines. As a first approach we here show that human CD34+ cells can be transferred into adult fish using retro-orbital injection. To be able to show human hematopoiesis in zebrafish we wanted to use fish that lack c-myb, which have been successfully used for fish to fish transplantations using whole kidney marrow stem cells [14]. Unfortunately, in these pilot experiments fish homozygous for the c-myb mutation did not survive the required polymerase chain reaction-based genotyping that precedes the transplantation procedure. Therefore, other lines that are genetically marked wish florescent markers, for instance rag knock-in mutant zebrafish, as recently described [20], would be useful in this regard. In addition, analogous to xenotransplantations in immune-deficient mice, various lines with variable degrees of immune deficiencies should be tried, for instance, fish with mutations in the il2g gene, as is similar to human X-SCID, could be proven useful. Moreover, immune-permissive fish with transgenic expression of human cytokines could also be very useful in the future.
The experiments reported here are merely a first step toward use of zebrafish as a xenotransplantation model. While therefore preliminary in nature, we report on these findings as stepping stone for other investigators to build on and develop fully “humanized” fish.
In summary, we here report that successful transplantation of human CD34+ HSPC is feasible, allowing trafficking and differentiation to be studied in vivo. Given the powerful use of zebrafish for finding evolutionary conserved factors that support hematopoiesis [21,22] our findings may help establishing full xenotransplantation in zebrafish. Such models would allow the visualization of microenvironmental clues acting on HSCs in the niche, which in turn could be exploited for clinical use in HSC expansion protocols.
Footnotes
Acknowledgments
We thank Davy de Witt (University of Leiden) for zebrafish maintenance and Claudia Tulotta (University of Leiden) for the CXCL12a−/− mutant fish. Cmyb mutants were kindly provided by Isabell Hess and Michael Schorpp (Max Planck Institute of Immunobiology and Epigenetics, Freiburg, Germany). This work was supported in part by the Netherlands Institute for Regenerative Medicine (NIRM; grant no. FES0908).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.