
Abstract
The timing and biological events associated with germ cell specification in chickens have not been determined yet. In this study, we report the origin of primordial germ cells (PGCs) and germ plasm dynamics through investigation of the expression of the chicken homolog of deleted in azoospermia-like (cDAZL) gene during germ cell specification. Asymmetric localization of germ plasm in the center of oocytes from preovulatory follicle stages leads to PGCs being formed in the center. During cleavage stages, DAZL expression pattern changes from a subcellular localization to a diffuse form before and after zygotic genome activation. Meanwhile, PGCs exhibit transcriptional active status during their specification. In addition, knockdown studies of cDAZL, which result in reduced proliferation, aberrant gene expression profiles, and PGC apoptosis in vitro, suggest its possible roles for PGC formation in chicken. In conclusion, DAZL expression reveals formation and initial positioning of PGCs in chickens.
Introduction
T
Importantly, components in the germ plasm play diverse roles in maintaining germ cell integrity, such as the induction of germ cell fate [10], migration, and viability [2], and germ cell proliferation and differentiation [11]. In the induction mode, germ cells are induced by signals from neighboring somatic cells during gastrulation, as studied in Mus musculus [12] and Ambystoma mexicanum [13]. Therefore, the presence of germ plasm from oocytes to cleavage-stage embryos is a crucial criterion to distinguish the mechanism of germ cell specification.
Primordial germ cells (PGCs) in avian species were initially identified in the germinal crescent region of Hamburger and Hamilton (HH) stage 4–10 embryos after the formation of the primitive streak and were thought to originate from the hypoblast based on their location [14]. PGCs in the germinal crescent were identified based on their morphological characteristics, such as a large cell size compared with the surrounding somatic cells, and a large amount of glycogen granules in the cytoplasm. The presence of a large amount of glycogen granules in PGCs permits the use of periodic acid-Schiff (PAS) solution to stain the cells [15].
Eyal-Giladi et al. reported the epiblastic origin of PGCs in a chick–quail chimera study [16]. Based on this report, further studies have focused on the mode of induction of PGC specification around Eyal-Giladi and Kochav (EGK) stage X [17,18]. These data revealed that only the central region, and not the marginal zone or the area opaca, of the blastoderm at EGK.X can give rise to PGCs [17,19]. Chicken PGCs had been well accepted to arise around EGK stage X in response to inductive signals because no evidence of the presence of germ plasm or PGCs has been detected during the cleavage stages, even in quail [20]. However, no available marker had existed to detect PGCs or the germ plasm, except PAS staining, when the study by Ginsberg et al. (1989) was performed.
After a chicken vasa homolog (CVH) was isolated, chicken PGCs could be traced backward to the initial stages of development [21]. Remarkably, CVH protein was colocalized with spectrin and mitochondrial clouds in growing oocytes, which indicates the presence of germ plasm in chickens. CVH was also localized to cleavage furrows and was restricted to only six to eight cells of the 300-cell stage embryo in a patch-like structure that resembled the pattern of germ plasm inheritance in zebrafish [22]. The above studies suggested that chicken germ cells might be specified by a maternally inherited determinant (preformation mode). Therefore, to completely understand the mechanism of germ cell specification in chickens, additional studies should be performed using reliable germ cell-specific markers, transplantation studies, and gene expression patterns during the intrauterine stages.
Deleted in azoospermia-like (DAZL), which encodes an RNA-binding protein, is a functional gene expressed in germ line cells. Its functions in germ cells have been well studied in vertebrate species and include meiotic progression [23] and maintaining the pluripotency of germ cells [24]. A previous study reported germ cell-specific expression of chicken DAZL from the embryonic to adult stages in chickens [25]. However, there is no report on DAZL expression during germ cell specification. In the present study, we investigated the expression of chicken DAZL and transcriptional status of PGCs during the intrauterine stages to analyze germ cell specification in chickens and to determine the timing and biological events during germ cell specification in chickens.
Materials and Methods
Experimental animals
White Leghorn (WL) hens (35–50 weeks old) were used to collect oocytes and intrauterine eggs. Chickens were managed according to our standard operation protocols, and all experimental procedures were approved by the Institutional Animal Care and Use Committee, Seoul National University, before the experiments were performed (SNU-070823-5). Transgenic (TG) chickens expressing green fluorescent protein (GFP) under the control of the CMV promoter were derived in our previous study [26]. GFP_TG embryos were produced by mating GFP_TG males with wild-type females.
Collection of intrauterine eggs, oocytes, and early embryos
The egg-laying time of WL hens was recorded and intrauterine eggs from EGK stages I–X were harvested using an abdominal massage technique [27]. Briefly, the abdomen was pushed gently until the shell gland was exposed; the surface of the shell gland expanded when an egg was located there for eggshell formation. After expansion of the surface of the shell gland, massaging was used to move the egg gently toward the cloaca until the intrauterine egg was released. To collect oocytes, three WL hens were killed and the ovary and the follicles were collected. The follicles were then divided into two stages based on their size: large white follicles and large yellow follicles with <0.5 and 3.5–4.5 cm diameters, respectively [28].
To collect early embryos, WL eggs were incubated with intermittent rocking at 37°C–38°C under 60%–70% relative humidity until sample collection. HH stage 4–11 embryos were collected for whole-mount in situ hybridization [29].
Staging of chicken embryos
To distinguish the developmental stages of chicken embryos, in this study, two kinds of conventional staging systems were used; one is EGK stages [27] and the other is HH stages [29]. The former covers intrauterine development until just after oviposition, contains EGK stages from I to XIV, and is classified by residence time in the uterus (total 20 h). The latter covers all embryonic development after oviposition until hatching, contains HH stages from 1 to 45, and is classified by incubation time and days (total 20 days). In brief, the stages before EGK stage X indicate intrauterine embryos and those after, including EGK stage X, indicate postovipositional embryos.
Analysis of intrauterine embryos
Intrauterine embryos were separated from the egg using sterilized paper [30], and the shell membrane and albumen were detached from the yolk. A piece of square filter paper (Whatman, Maidstone, United Kingdom) with the hole in the center was placed over the germinal disc. After cutting around the paper containing the intrauterine embryo, it was gently turned over and transferred to saline to further remove the yolk and vitelline membrane to allow embryo collection [31]. The harvested embryos were fixed with 4% (w/v) paraformaldehyde in 1× phosphate-buffered saline (PBS), and then classified according to cleavage [27]. Unfertilized and abnormal embryos were identified according to the morphological criteria of the cleavage furrows.
Whole-mount in situ hybridization
To make hybridization probes, total RNA from magnetic-activated cell sorting (MACS)-positive PGCs at E6.5 was reverse transcribed, and the cDNA was amplified using cDAZL-specific primers (fwd: 5′-CGT CAA CAA CCT GCC AAG GA-3′ and rev: 5′-TTC TTT GCT CCC CAG GAA CC-3′, product size 540 bp), as described previously [25]. The polymerase chain reaction (PCR) products of the correct size were cloned into pGEM-T vector (Promega, Madison, WI).
After sequence verification, the recombinant plasmids containing the gene were amplified using T7- and SP6-specific primers (T7: 5′-TGT AAT ACG ACT CAC TAT AGG G-3′ and SP6: 5′-CTA TTT AGG TGA CAC TAT AGA AT-3′) to prepare the template for labeling the hybridization probes. The digoxigenin (DIG)-labeled sense and antisense cDAZL hybridization probes were transcribed in vitro using a DIG RNA labeling kit (Roche Diagnostics, Indianapolis, IN). A standard protocol in chickens was followed for whole-mount in situ hybridization [32].
Immunohistochemistry and transmission electron microscopy
The harvested whole-mount and paraffin-embedded samples were immunostained. For immunostaining, oocytes and intrauterine embryos (after deparaffinization for paraffin-embedded tissues; thickness 8 μm) were washed thrice with PBS and blocked with blocking buffer (PBS containing 5% goat serum and 1% bovine serum albumin) for 1 h at room temperature. Samples were then incubated at 4°C overnight with rabbit anti-cDAZL, anti-CVH, or anti-RNA polymerase II CTD repeat antibodies (Abcam, Cambridge, MA). The polyclonal antibodies against N-terminal peptides of cDAZL (aa 2–17) and CVH (aa 42–57) were raised in a rabbit host and their antisera were purified. After washing twice with PBS, samples were incubated with secondary antibodies labeled with phycoerythrin or fluorescein isothiocyanate (FITC; Santa Cruz Biotechnology, Santa Cruz, CA) for 4 h at room temperature. Samples were then mounted with Prolong Gold antifade reagent with 4′,6-diamidino-2-phenylindole (DAPI; Invitrogen, Carlsbad, CA) and visualized using fluorescence microscopy.
To prepare specimens for transmission electron microscopy (TEM), samples were dehydrated using a graded ethanol series, embedded in Spurr's resin, and cut on an ultramicrotome (MT-X; RMC, Tucson, AZ). Samples were then stained with 2% uranyl acetate and Reynold's lead citrate for 7 min each and observed under TEM (LIBRA 120; Carl Zeiss, Oberkochen, Germany), as described previously [33].
Knockdown of cDAZL in PGCs
For RNA interference assays, cDAZL-specific siRNAs were designed against unique regions according to siRNA design rules (ST Pharm, Seoul, Korea). Two siRNA sequences, siRNA_516 (5′-CUG CAU GUA AGA ACA CAA G-3′) and siRNA_957 (5′-UUC UUG UGA UAC AAA GGU G-3′), and a control siRNA (5′-UGC UUU AAC CAC CGC AUC C-3′), which was not scrambled cDAZL siRNA, were then synthesized and purchased from ST Pharm.
To validate the knockdown efficiency of cDAZL, siRNAs were transfected into in vitro cultured PGCs using Lipofectamine (Invitrogen), as described previously [34]. Forty-eight hours after transfection, total RNA was extracted using TRIzol reagent, and cDAZL knockdown was analyzed using real-time quantitative PCR (qPCR). Relative cDAZL expression was calculated after the threshold cycle was normalized to that of chicken glyceraldehyde 3-phosphate dehydrogenase (GAPDH). The sequences of the cDAZL-specific and GAPDH primers are shown in Supplementary Table S1 (Supplementary Data are available online at
In vitro analysis of cDAZL knockdown effects on PGCs
cDAZL siRNA_957 was selected for further in vivo and in vitro analyses. To elucidate the effect of cDAZL knockdown on PGCs in vitro, the expression of germ cell-, pluripotency-, and differentiation-related genes was examined using real-time qPCR 48 h after transfection. The sequences of all primers used are provided in Supplementary Table S1. Relative gene expression was calculated after normalization to GAPDH expression. Cell proliferation efficiency after cDAZL knockdown was quantified using WST-1 cell proliferation reagent (TaKaRa, Tokyo, Japan).
To assess apoptotic cell death after cDAZL knockdown, Annexin V/propidium iodide (PI) staining was performed and analyzed by flow cytometry. Annexin V and PI double-negative cells were considered viable, and Annexin V-positive and PI-negative (early apoptotic) cells or Annexin V- and PI-positive (late apoptotic) cells were considered apoptotic. The induction of apoptosis was confirmed using an in situ cell death detection kit and tetramethyl rhodamine (TMR) dye (Roche Diagnostics) and counterstained with DAPI (Invitrogen). Photographs of apoptotic cells were captured under a fluorescence microscope.
Statistical analysis
Statistical analysis was performed using SAS version 9.3 software (SAS Institute, Cary, NC). When a significant main effect was detected by analysis of variance (ANOVA) using SAS, the least-squares analysis was used to compare the different treatments. Differences between control and treatment groups were deemed to be significant when P < 0.05.
Results
PGC-specific expression of cDAZL mRNA and protein during early germ cell development in chickens
cDAZL mRNA was expressed specifically in early migrating PGCs in chickens at HH stages 4–11. At HH stage 4, cDAZL was expressed in cells in the germinal crescent region where chicken PGCs are located (Fig. 1A). At HH stages 4–7, cDAZL-expressing PGCs migrated laterally (Fig. 1B, C). At HH stage 11, cDAZL-expressing PGCs localized near the anterior vitelline veins of the embryos (Fig. 1D). As a control, whole embryos at EGK stages I–II and HH stage 4 were hybridized with sense probes and gave no signals (Supplementary Fig. S1).
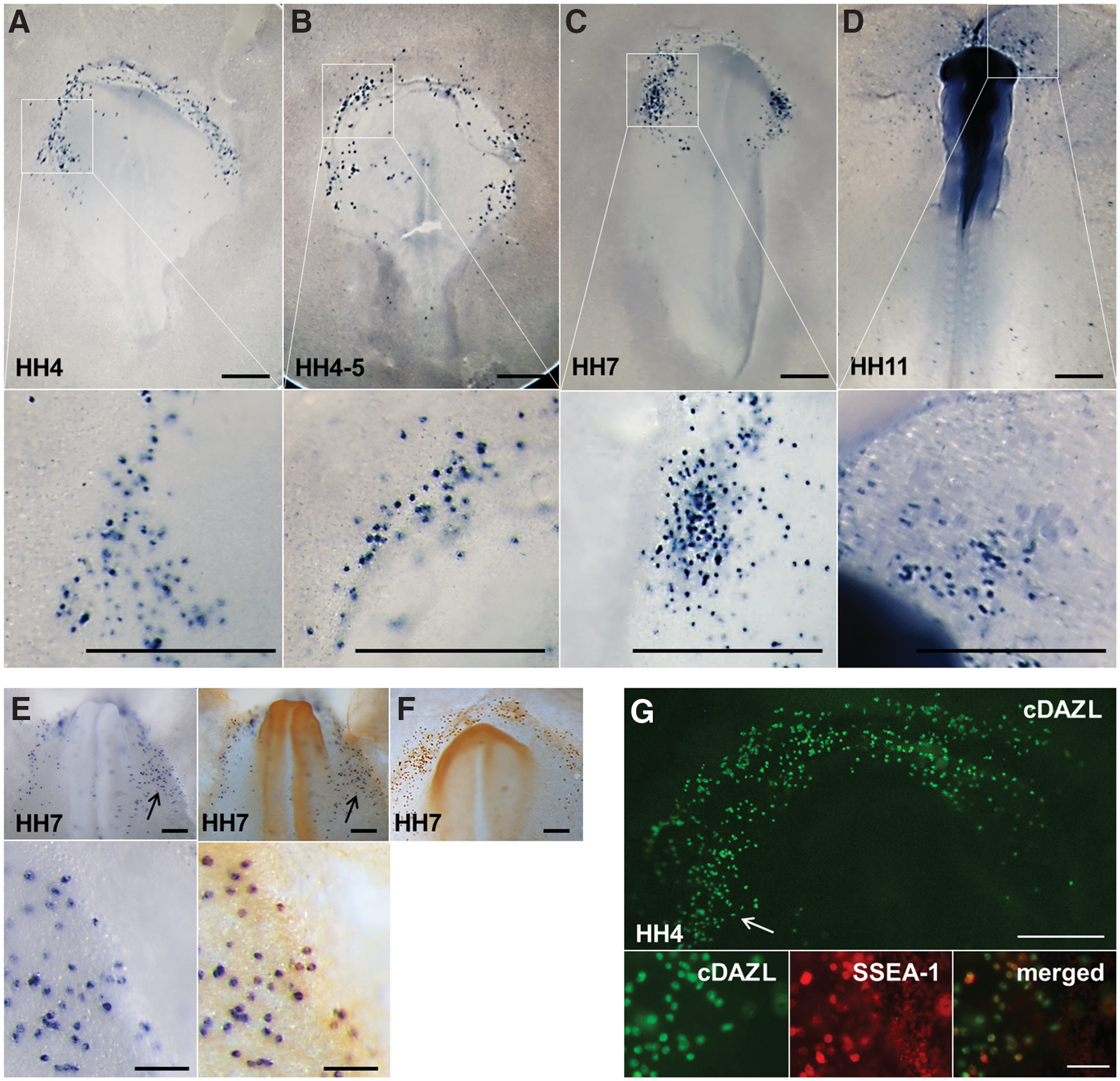
PGC-specific expression of cDAZL mRNA and protein. Whole embryos at HH stage 4
To confirm whether cDAZL-expressing cells are PGCs or not, immunostaining with CVH after in situ hybridization of cDAZL or double immunostaining with cDAZL and stage-specific embryonic antigen-1 (SSEA-1) antibody was conducted (Fig. 1E, G). As a control, one embryo was immunostained with CVH only (Fig. 1F). Most of cDAZL-positive cells were double positive for CVH or SSEA-1, indicating that cDAZL is a PGC-specific marker.
Expression of cDAZL mRNA and protein and localization during the oocyte-to-zygote transition
To investigate the details of germ plasm dynamics during the oocyte-to-zygote transition, oocytes were classified into three stages during their development; growing oocytes, which are identifiable by sectioning, oocytes from large white follicles with <0.5 cm diameter, and oocytes from large yellow follicles with 3.5–4.5 cm diameter. In addition to oocytes, zygote stage and first cleavage-stage embryos were harvested for whole-mount in situ hybridization or immunohistochemistry.
In growing oocytes, cDAZL protein showed patch-like localization in the peripheral cortex region of smaller oocytes and diffused expression in the cytoplasm of the larger oocyte (Fig. 2A). In oocytes from the large white follicles, cDAZL mRNA (Fig. 2B) and protein (Fig. 2C) were detected in the cytoplasm. In oocytes from the large yellow follicles, cDAZL mRNA (Fig. 2D) and protein (Fig. 2E) were strongly localized around the germinal vesicle (GV).
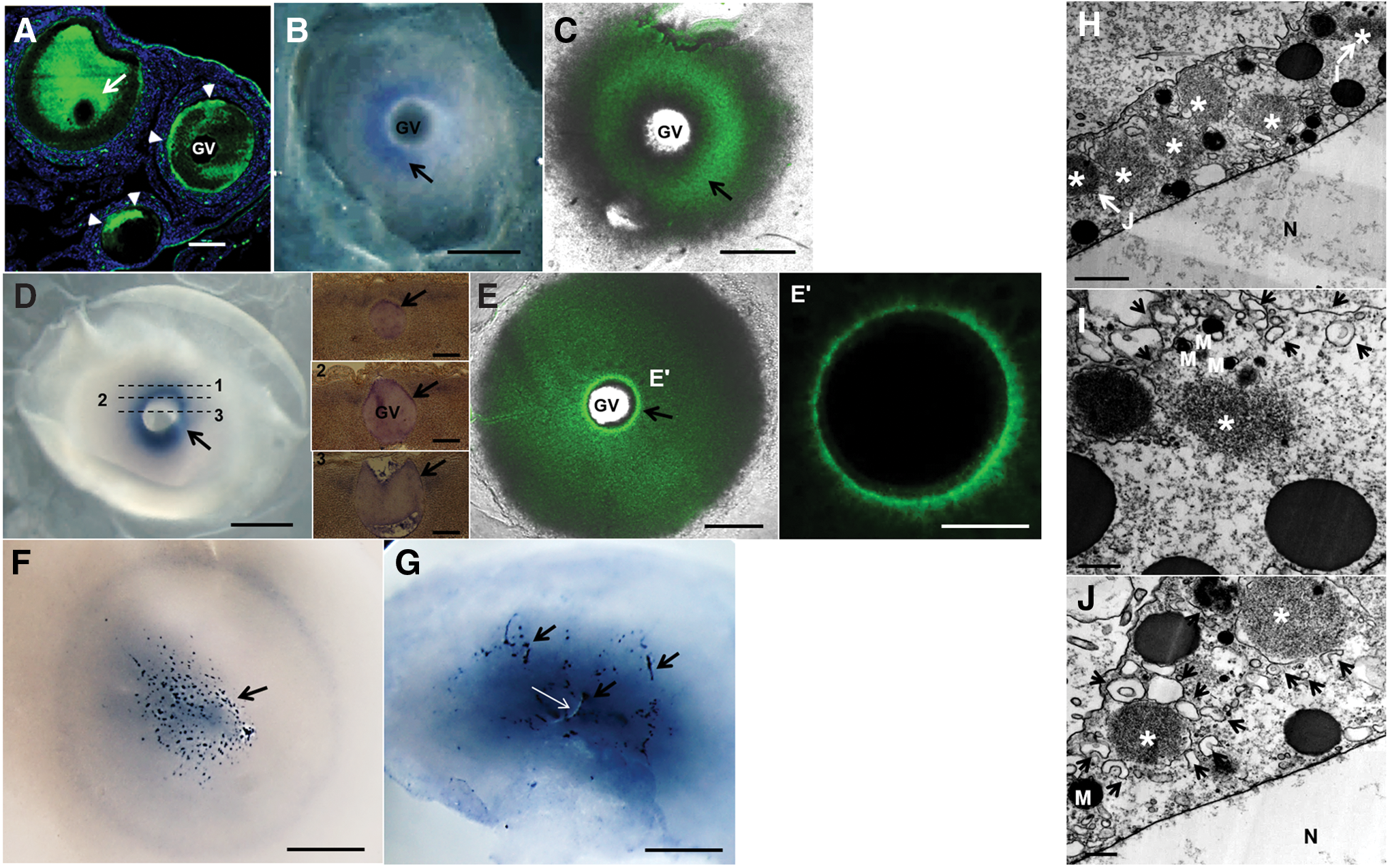
Expression of cDAZL mRNA and protein during the oocyte-to-zygote transition and the presence of electron-dense granules in the perinuclear region of oocytes.
In zygotes, granular cDAZL mRNA structures were distributed in the central region of the cytoplasm (Fig. 2F). After the beginning of the first cleavage, cDAZL mRNA was also localized in the cleavage furrow (Fig. 2G). Thin sections of the region near the GV of oocytes from the large yellow follicles revealed the presence of nonmembranous electron-dense granules (Fig. 2H). The electron-dense granules were associated with endoplasmic reticulum (Fig. 2I, J). Taken together, these data indicate that cDAZL mRNA and protein were present in the germ plasm, which was localized in the perinuclear region of oocytes.
Expression of cDAZL mRNA and protein in intrauterine-stage embryos
The localization of germ plasm and the formation of PGCs were investigated by tracing the expression of cDAZL mRNA and protein in intrauterine-stage embryos. During the initial cleavage stages (EGK stage I), cDAZL mRNA was localized in cleavage furrows (Fig. 3A). After the embryos grew with an increase in the number of cell layers, cDAZL mRNA was localized in the subcellular regions of several central cells from EGK stages III to V, which revealed the subcellular localization of germ plasm (Fig. 3B, C). Some cDAZL mRNA-containing granules divided into two daughter cells during mitosis (Fig. 3B, sectioned view).
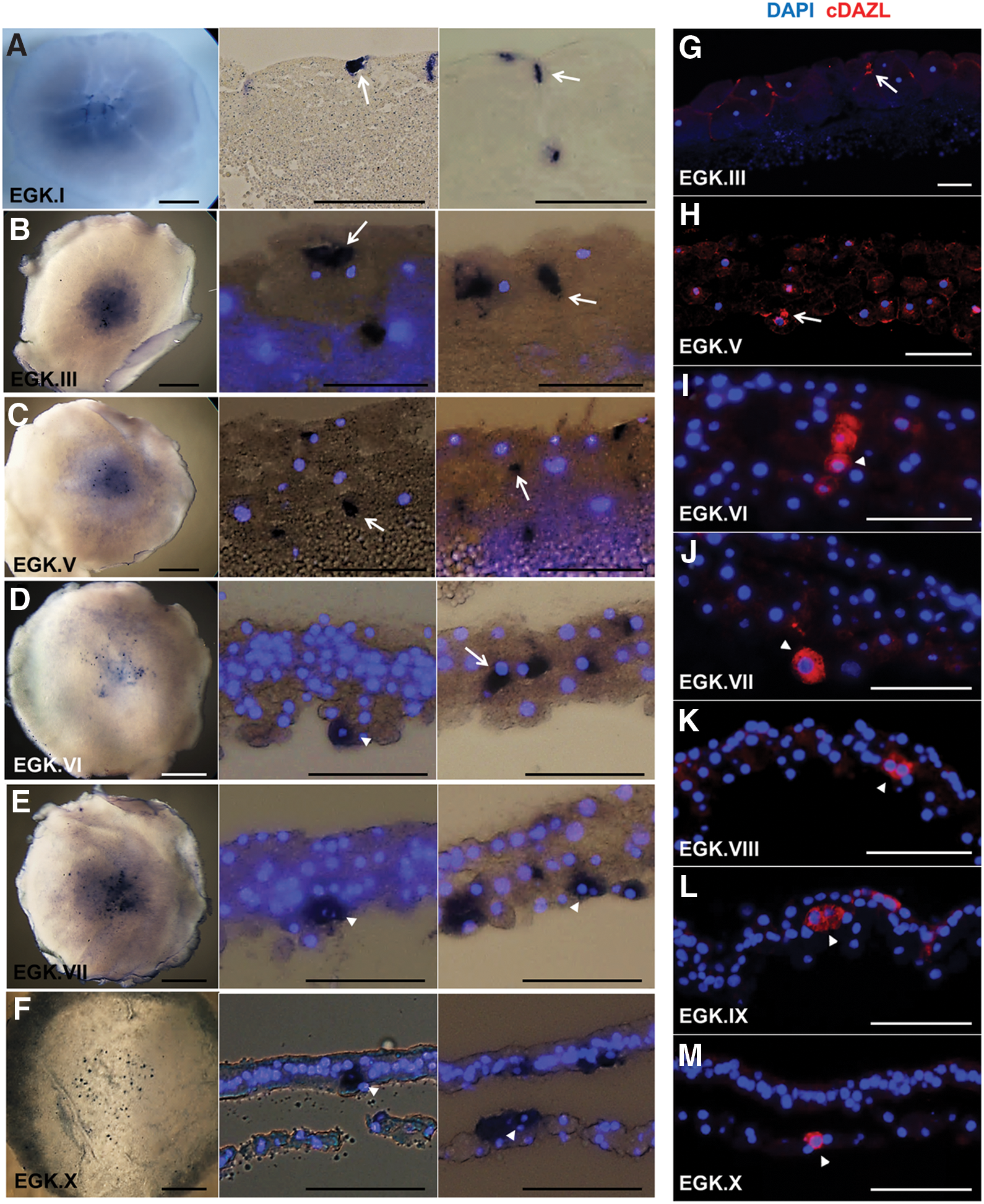
Expression dynamics of cDAZL mRNA and protein in intrauterine embryos. Approximate cell numbers are 1–3,000 during EGK stages I–V, 3,000–30,000 during EGK stages V–VII, and 30,000–60,000 during EGK stages VII–X.
From EGK stages VI to X, cDAZL mRNA was expressed strongly in the cytoplasm of several PGCs (Fig. 3D–F), suggesting the initiation of germ line-specific transcription. In addition, some cDAZL mRNA was localized between dividing cells (Fig. 3E, left sectioned view). During the intrauterine stages, cells containing germ granules or expressing cDAZL in the cytoplasm were present anywhere among the cell layers from the top to the bottom. From the initial cleavage stage (EGK stage I) to oviposition (EGK stage X), the expression of cDAZL mRNA in cleavage furrows, subcellular region, and cytoplasm was restricted to the area pellucida, where PGCs predominantly reside.
In addition, the number of PGCs was counted during their specification using whole sections (10 μm thickness) of each embryo after whole-mount in situ hybridization. The mean numbers of precursor PGCs (pPGCs) and/or PGCs at EGK stages V, VI, VII, and VIII were 49 ± 5.6 (n = 5), 60 ± 18.8 (n = 4), 82.2 ± 15.0 (n = 5), and 72 ± 11.2 (n = 3), respectively (Table 1).
Cells had a granular structure (EGK stages V–VI) or diffuse expression (EGK stages VI–VIII) of cDAZL mRNA.
EGK, Eyal-Giladi and Kochav; PGC, primordial germ cells.
To investigate the timing of the appearance of PGCs, anti-cDAZL antibody staining was performed using sectioned intrauterine embryos of EGK stages III–X. At EGK stage III, cDAZL protein was localized in the cleavage furrows (Fig. 3G). At EGK stage V, the subcellular localization of cDAZL protein was observed in a small number of centrally localized cells (Fig. 3H). From EGK stages VI to X, cDAZL was expressed strongly in the cytoplasm of several cells, which may have indicated PGCs (Fig. 3I–M). In addition, cDAZL-expressing PGCs were located anywhere among the cell layers in these stages, and several cells revealed a cluster, indicating possible mitosis.
Initiation of ZGA in chickens and maternal origin of cDAZL in early chick embryos
To identify whether the expression of cDAZL in cleavage-stage embryos originated maternally or was newly made by zygotic genome activation (ZGA), transgenic chicken expressing GFP (GFP_TG) under the control of the CMV promoter was analyzed. The GFP expression of cleavage-stage embryos that were born after mating TG males with wild-type female chickens was analyzed (Supplementary Fig. S2). Weak GFP expression was observed in a few central cells from late EGK stage II, which became strong in many central cells at EGK stage IV (Supplementary Fig. S2). At EGK stage X, whole embryonic cells from the center to the peripheral area expressed GFP (Supplementary Fig. S2).
Because there was the possibility of unequal GFP expression among early cell types under the CMV promoter, we also investigated the phosphorylation of RNA polymerase II (p-pol II) in early embryos (Supplementary Fig. S2). The p-pol II showed a very similar expression pattern to that of GFP in TG chick embryos during the cleavage stages (Supplementary Fig. S2).
To confirm the presence of cDAZL mRNA before ZGA, the expression of cDAZL was analyzed in unfertilized oocytes collected 7 h after the oviposition of previous eggs. cDAZL mRNA was dispersed in the central region as a granular structure, similar to fertilized embryos (Supplementary Fig. S3), which revealed that the initial localization of cDAZL mRNA was maternally inherited. Therefore, PGCs might emerge from germ plasm containing cDAZL mRNA that is maternally inherited.
Transcriptional status of intrauterine embryos and PGCs
To determine whether intrauterine embryos and PGCs are transcriptionally active or inactive, phosphorylation status of RNA polymerase II (p-pol II) was investigated. During EGK stages IV–X, expression of p-pol II showed a dynamic pattern (Fig. 4A). At EGK stage IV, central cells except peripheral cells were positive for p-pol II, and most of the cells at EGK stage V were p-pol II positive, indicating that ZGA occurs from the center to the periphery. Expression of p-pol II was reduced in the upper layers of EGK stage VI, and most of the cells were negative for p-pol II at EGK stage VIII, indicating gradual decrease of p-pol II from upper layers to lower layers. P-pol II-positive cells started to increase again from EGK stage IX, and most of the cells at EGK stage X became positive for p-pol II.

Phosphorylation of RNA polymerase II in intrauterine embryos and PGCs during their specification. Intrauterine embryos of EGK stages IV–X were sectioned and immunostained with antiphosphorylated RNA polymerase II (p-pol II) only
Expression of p-pol II in PGCs during EGK stages VI–X was then investigated (Fig. 4B). At EGK stage VI, PGCs were positive for p-pol II. At EGK stage VII, most of the cells, including PGCs, showed weak or negative expression of p-pol II (Fig. 4A, B). At EGK.VIII, most of preblastodermal cells, including PGCs, were negative for p-pol II. At EGK.X, PGCs and other somatic cells reacquired p-pol II (Fig. 4A, B). Taken together, PGCs exhibit transcriptionally active status during intrauterine stages at least from EGK stage VI (Fig. 4).
Effect of cDAZL knockdown on PGCs in vitro
To assess the function of cDAZL in PGCs, RNA interference using siRNA-mediated knockdown was performed. For cDAZL knockdown, two candidate siRNAs, siRNA_516 and siRNA_957, were designed. Forty-eight hours after transfection of in vitro cultured PGCs with the siRNAs, the cDAZL expression levels in control and knockdown PGCs were assessed by qPCR to determine the knockdown efficiency. Compared with the control, siRNA_516 suppressed 49% of cDAZL expression, whereas siRNA_957 caused a more marked (73%) suppression of cDAZL expression (Fig. 5A). Therefore, siRNA_957 was used in all subsequent analyses.
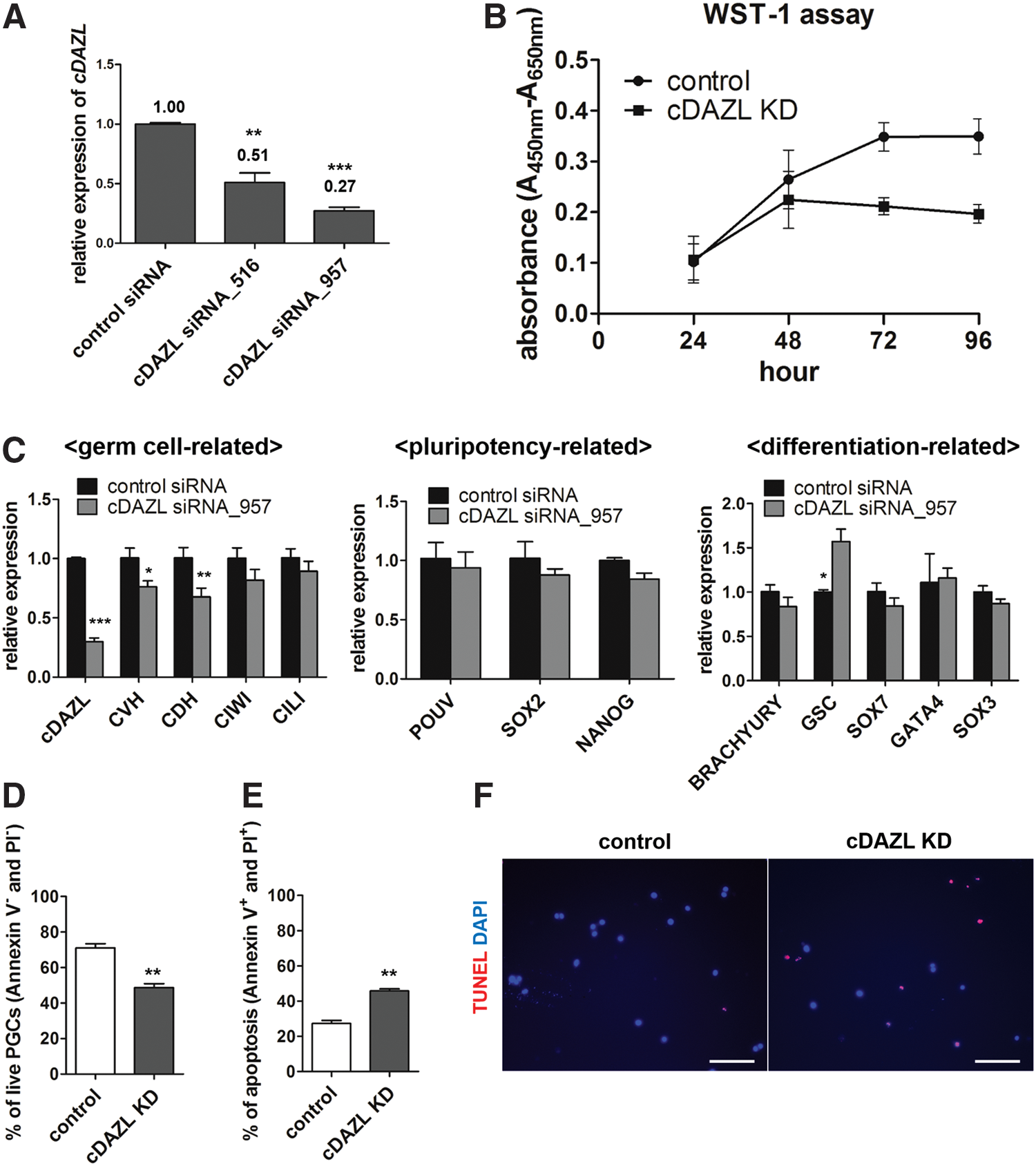
Effects of cDAZL knockdown on PGC integrity in vivo and in vitro.
To assess the effect of cDAZL knockdown on PGCs in vitro, control siRNA and siRNA_957 were transfected into cultured PGCs. WST-1 assays were performed 96 h after transfection to investigate PGC proliferation in vitro. The proliferation of cDAZL knockdown PGCs decreased continuously after 48 h (Fig. 5B).
To investigate the effects of cDAZL knockdown on PGC gene expression, qPCR was used to assess the expression of germ cell-, pluripotency-, and differentiation-related genes in PGCs 48 h after cDAZL knockdown. Among the germ cell-related genes, CVH and chicken dead end homolog (CDH) expression levels decreased significantly after cDAZL knockdown. Among the differentiation-related genes, goosecoid homeobox (GSC) significantly increased after cDAZL knockdown. However, no significant changes were observed in the expression of pluripotency-related genes (Fig. 5C).
We next assessed the effect of cDAZL knockdown on the viability and apoptosis of in vitro cultured PGCs. First, Annexin V/PI double staining was performed. Compared with the control, cDAZL knockdown significantly decreased the proportion of cells that stained double negative for Annexin V and PI (live cells; Fig. 5D), and an increased proportion of PGCs stained double positive for Annexin V and PI (dead cells; Fig. 5E). In addition, a significant apoptotic signal, in the form of TMR red, was detected in cDAZL knockdown PGCs compared with the control (Fig. 5F). Taken together, these data suggest that cDAZL knockdown reduced the proliferation and viability of PGCs and also caused aberrant gene expression patterns.
Discussion
During the specification of PGCs, several germ cell-specific genes, including NANOS, VASA, PIWI, and DAZL, are expressed and play a role in maintaining germ cell competency among various species [5,6,35]. DAZL, a germ line-specific gene, functions as an RNA-binding protein in various species [36] and also has important roles in meiosis [23,37] and maintaining the pluripotency of germ cells [24]. In addition, cDAZL is expressed specifically in germ cells from embryonic to adult stages in chickens [25].
Unlike in mammals, in which DAZL expression starts from postmigratory stages of PGCs, the present study found that cDAZL is expressed specifically in PGCs during their migration from EGK stage X (the prestreak stage) to HH stage 11, which suggests the possibility of cDAZL expression before oviposition. In addition, cDAZL-expressing cells were positive for CVH or SSEA-1, which is one of the PGC-specific markers in chickens [38]. Therefore, we selected DAZL as a marker to trace the origin of PGCs in chickens. Because DAZL is present in the germ plasm of some vertebrates [35,39], we hypothesized that chicken DAZL is localized in the germ plasm and that germ cells are specified by germ plasm.
The presence of germ plasm structures and the asymmetric localization of germ plasm-related genes in oocytes and cleavage-stage embryos are important criteria to determine the mode of germ cell specification and the initial localization of PGCs. In D. melanogaster, only posterior pole cells, containing pole plasm that is assembled during oogenesis, will give rise to germ line cells, and the pole plasm is a necessary and sufficient condition for becoming PGCs [7,40]. In C. elegans, electron-dense granules named P granules are localized into the posterior region during zygotic formation, and Z2 and Z3 cells that harbor P granules become PGCs, although P granules are a necessary, but not sufficient, condition for becoming PGCs [8,41].
In chickens, CVH protein is localized in the cortex region of growing oocytes, and then in cleavage furrows [21]. However, how germ plasm moves from the peripheral cortex in growing oocytes to the central cleavage furrows in embryos was unknown [21]. First, we investigated cDAZL protein expression in growing oocytes and found that cDAZL protein was localized in the peripheral cortex region of growing oocytes as a patch-like structure and showed diffused expression in the cytoplasm in the larger oocytes, that is, a similar localization pattern when compared with CVH protein [21].
However, in the oocytes from the large yellow follicles, which are preovulatory follicles with 3.5–4.5 cm diameters, cDAZL mRNA and protein were localized in the central perinuclear region in the buildup to ovulation and were maintained during zygote formation, which is addressed first in this study. Electron-dense granules with numerous SER and mitochondria were also present near the GV of the oocytes. In addition, cDAZL localization was maintained in the central region during the cleavage progress.
These results can explain the previous reports that identified a central position for PGCs after oviposition [17,19]. Taken together, asymmetric localization of germ plasm in the center of oocytes from preovulatory follicle stages leads to PGCs being formed in the center later on, as pore plasm in D. melanogaster and P granules in C. elegans do.
We identified the kinetics of DAZL gene expression during the cleavage stages. During intrauterine embryo development, DAZL was localized in cleavage furrows during initial cleavage progression (EGK stages I–III) and in the subcellular region during further cleavage (EGK stages IV–VI), then finally diffused in the cytoplasm after EGK stages VI–VII. Notably, the subcellular localization of DAZL disappeared and was replaced by diffuse expression in the cytoplasm from EGK stage VI, which was thought to be the initiation of germ line-specific transcription. Therefore, we suggest that pPGCs containing germ plasm give rise to PGCs at EGK stages VI–VII with initiation of germ line-specific transcription in chickens.
Germ line-specific transcription, which starts after germ cell specification in C. elegans [42] and D. melanogaster [43] and starts during germ cell specification in mice [44], is closely related with transcriptional repression in germ cells [45]. In the preformation mode, maternally inherited C. elegans PIE-1 or D. melanogaster Pgc proteins block global transcription of mRNA in PGCs by inhibiting p-pol II [46,47]. In the induction mode of mice, however, there are no maternally inherited molecules for global transcriptional repression. Instead, Blimp1 (B-lymphocyte-induced maturation protein 1) is expressed from E6.25 and inhibits transcription of a specific set of genes for somatic lineages [48] during germ cell specification in mice. In this regard, change in transcriptional status of PGCs during their specification is one of the conserved mechanisms among species.
In our results, chicken PGCs exhibited transcriptionally active status during their specification, showing positive staining of p-pol II in their nuclei. Although they were transiently negative for p-pol II at EGK stage VIII, it is not specific to PGCs, and neighboring somatic cells were also negative at that time. Our results show that chicken PGCs are transcriptionally active during their specification as early as at EGK stage VI, which might cause the diffused expression pattern of DAZL mRNA and protein. Therefore, to address mechanisms of transcriptional changes in chicken PGCs, related gene expression in these early stages should be investigated in further studies.
Previous studies reported that avian germ cells originate from epiblasts, and not hypoblasts [16,49]. At EGK stage X, the area pellucida has the epiblast in the upper layer and the primary hypoblast in the lower layer; primary hypoblasts were thought to be derived by polyingression from the epiblast [50].
However, in the current study, PGCs were consistently in both the upper and lower layers, at least from EGK stages VI to VII (5–6 cell layer thickness), until EGK stage X (1–2 cell layer thickness). In addition, serial sections from EGK stage VII to EGK stage X revealed that the initial segregation of the primary hypoblast might already start at EGK stage VII, which suggests that most upper layers are future epiblasts and that the lower layers are future hypoblasts at EGK stage VII.
Therefore, PGCs are likely already present in both the epiblasts and hypoblasts in prestreak-stage embryos. The expression pattern of lineage-specific genes during intrauterine embryo development should be investigated to clarify when the epiblastic and hypoblastic lineages are specified.
Although cDAZL was continuously present from initial development, its origin remains unclear. Therefore, we investigated the expression of cDAZL in unfertilized oocytes and the timing of ZGA in chickens. One previous study demonstrated that exogenous DNA began to be expressed in the central cells of EGK stage IV [51], but details on ZGA were not reported. In our study with GFP transgenic embryos, the unsynchronized expression of GFP started from late EGK stage II in the central cells. Because the unubiquitous GFP expression could be driven by the CMV promoter, we also investigated p-pol II expression during the intrauterine stages and obtained similar results as reported in our previous study [52].
These data suggest that ZGA in chickens first occurs in the central cells between EGK stages II and III, and then expands toward the periphery; it does not occur simultaneously in all cells. Therefore, these results, including expression of cDAZL mRNA in unfertilized oocytes and intrauterine embryos and the timing of ZGA, suggest that cDAZL mRNA as well as protein is maternally inherited from the germ plasm, which comes from oocytes rather than zygote transcription.
Its expression pattern suggests that cDAZL has important functions in PGCs. In mice, Dazl is first expressed in germ cells at embryonic day 11.5, but is not expressed during germ cell specification [53]. Although mRNA and protein of the Dazl or Vasa gene are expressed in oocytes and cleavage embryos in mice and humans, there is no subcellular localization as a germ plasm [54 –56]. Meanwhile, our study demonstrated that cDAZL was maternally inherited, present as a germ plasm, and its expression was specific to PGCs in chickens.
This suggests that the mechanism of action of DAZL differs between mice and chickens, even though it is highly conserved among vertebrate species [25]. However, we could not exclude the role of signaling or induction from neighboring somatic cells since we do not know what causes changes of DAZL expression pattern at EGK stage VI. One possible candidate is neighboring somatic cells, which are transcriptionally active at the same stage. In mice, DAZL knockout has no effect on PGCs during their specification or before their settlement into gonads, even though it leads to reduced germ cell numbers, germ cell apoptosis, and infertility after embryonic day 12.5 [24,53].
To verify a potential role of DAZL in chickens, we knocked down cDAZL in vitro. cDAZL knockdown in PGCs caused decreased proliferation, aberrant expression of germ cell-related genes, and increased apoptosis, which suggests that DAZL is required for maintaining the integrity of PGCs. However, to clarify a role of DAZL especially for germ cell specification in chickens differing from mammals, functional studies with cleavage-stage embryos should be conducted.
In conclusion, we revealed the origin of PGCs and germ plasm dynamics during germ cell specification in chickens (Fig. 6). Considering the timing of ZGA in chickens, the DAZL expression pattern changing from a subcellular localization (pPGC) to a diffuse form (PGC) seems to depend on the initiation of zygotic expression. In addition, PGCs were transcriptionally active during their specification. In addition, unlike previous studies, our results demonstrated that chicken PGCs are formed before epiblast formation, and then incorporate into epiblasts or hypoblasts at EGK stage X.

Schematic diagram of germ plasm dynamics and PGC formation in chickens. Green indicates the expression or localization of the germ line-specific gene such as cDAZL. During the oocyte-to-zygote transition, germ granules are localized around the GV of oocytes from preovulatory yellow follicles and are distributed in the center of the zygote. During the cleavage stages, germ granules are localized in cleavage furrows at earlier stages and in a subcellular region of pPGCs after the formation of preblastodermal cells. Germ line-specific transcription was revealed by the diffuse cytoplasmic expression in PGCs. PGCs were present randomly in the central region of an embryo regardless of the cell layer. pPGCs, precursor PGCs. Color images available online at
Footnotes
Acknowledgments
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIP) (no. 2015R1A3A2033826) and Bio-industry Technology Development Program (IPET312060-5), Ministry for Food, Agriculture, Forestry and Fisheries, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.