
Abstract
Mesenchymal stem cells (MSCs) exhibit a potent immunomodulatory capacity and have been applied to treat diseases such as graft versus host disease and severe autoimmune diseases. However, the mechanism underlying their immunosuppressive effect is not yet completely understood. Here, we investigated the role of the CD73/adenosine pathway in immune modulation by MSCs using a mouse model of experimental autoimmune uveitis (EAU). Moreover, we examined the in vitro modulatory effect of MSCs mediated through the CD73/adenosine pathway in human and mouse T cells. We found that the severity of EAU was significantly attenuated by MSCs; however, most therapeutic effects of MSCs were lost by pretreatment with a CD73 inhibitor. The inhibitory mechanism of MSCs might be contributed by CD73 on MSCs that cooperated with CD39 and CD73 on activated T cells to produce adenosine, resulting in inhibition of T-cell proliferation. Furthermore, MSCs increased the expression of CD73 on CD4+ T cells, and transforming growth factor-β1 (TGF-β1) was the only tested cytokine that contributed to upregulation of CD73. Hence, our study demonstrates that the CD73/adenosine pathway involves the immunomodulatory function of MSCs in autoimmune responses.
Introduction
M
A large amount of adenosine triphosphate (ATP) is released into the extracellular space upon cell damage by inflammation or tissue injury, which evokes proinflammatory effects. It has been recently found that CD39, an ectonucleotidase, hydrolyzes ATP/adenosine diphosphate (ADP) to adenosine monophosphate (AMP), and CD73, another ectonucleotidase, can convert the AMP to adenosine, which conversely mediates anti-inflammatory effects on diverse immune cells [9 –11]. This shift from an ATP-induced proinflammatory microenvironment to an adenosine-driven, anti-inflammatory milieu plays strategic roles in modulating the purinergic signals transmitted to immune cells [9 –11]. Recent in vitro studies indicate that coexpression of CD39/CD73 and adenosine generation mediates the suppression of T-cell proliferation and effector functions of Th17 cells by MSCs [12,13]. Chen et al. recently demonstrated that MSCs derived from human gingiva suppress collagen-induced arthritis depending partially on CD39/CD73 expression in MSCs [14].
CD73 is highly expressed on the MSC membrane and is an important surface marker of MSCs. However, in contrast to previous reports, we have not detected significant CD39 expression on human MSCs (hMSCs) or mouse MSCs (mMSCs) in our laboratory. T cells also express CD39 and CD73, and CD73 expression is downregulated after T-cell activation. It is currently unknown whether MSCs exert immunosuppressive effects with low CD39 expression, and the mechanism of CD73 in the therapeutic efficacy of MSCs in autoimmune diseases is unclear, especially how CD73 on MSCs and CD39/CD73 on T cells cooperate under inflammatory conditions. In this study, we used an experimental autoimmune uveitis (EAU) model to further clarify the therapeutic role and immunomodulatory mechanism mediated by CD73 on MSCs. We found that mouse adipose tissue-derived MSCs (mAD-MSCs) significantly attenuate EAU in mice through the CD73/adenosine pathway. Both CD39 and CD73 expressed on T cells jointly engaged this pathway, and CD73 expression on T cells was upregulated by MSCs, which might further enhance autoimmune suppression. Our findings imply an additional and novel mechanism that participates in the immunomodulatory function of MSCs for the treatment of uveitis and possibly other autoimmune diseases, which might provide a new target for improving the immunosuppressive potential of MSCs by upregulating their CD73 expression.
Materials and Methods
Animals
C57BL/6 mice (B6, 99 female aged 8–10 weeks old for EAU induction and 35 male mice aged 12–14 weeks old for mMSC isolation) were purchased from Vital River (Beijing, China) and maintained under pathogen-free conditions. All animal experiments were approved by the Laboratory Animal Care and Use Committee of Tianjin Medical University and conformed to the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research.
Isolation and characterization of adipose tissue-derived MSCs
Inguinal and celiac adipose tissues obtained aseptically from male mice were digested with 0.075% type I collagenase (Sigma-Aldrich, St. Louis, MO), washed with phosphate-buffered saline (PBS), and then transferred to a culture flask. The cells were cultured in α-MEM (Gibco, Grand Island, NY) supplemented with 20% fetal bovine serum (Gibco), 100 U/mL penicillin, and 100 μg/mL streptomycin (Gibco) at 37°C with 5% CO2 in a humidified incubator. Passage 3–5 cells were used in experiments. For hMSCs, the adipose tissue from human breast was collected after informed consent had been obtained, under the guidelines for research involving human subjects, and with the approval of the Ethics Committee of Tianjin Medical University Eye Hospital, Eye Institute. The cells were cultured as mMSCs.
The mMSCs were characterized by their capacity to differentiate into endothelial cells, adipocytes, and osteocytes under appropriate in vitro conditions. Further characterization was based on positivity for CD29 (eBioscience), CD44 (eBioscience), CD73 (BioLegend, San Diego, CA), CD90 (BioLegend), CD105 (BioLegend), and Sca-1 (eBioscience), and negativity for hematopoietic markers CD34 (eBioscience), CD45 (BioLegend), and CD11b (eBioscience). We also detected the expression of CD39 (eBioscience).
Animal model of adoptively transferred uveitis
Induction of EAU in female mice was performed as described previously [15]. EAU was first induced in mice by active subcutaneous immunization with 150 μL of an emulsion containing 150 μg interphotoreceptor retinoid-binding protein (IRBP) 1–20 (GPTHLFQPSLVLDMA KVLLD; Sangon, Shanghai, China) and 800 μg mycobacterium tuberculosis H37Ra (Difco, Detroit, MI) in incomplete Freund's adjuvant (Sigma-Aldrich), which was distributed in more than six sites on the tail base and flank. A single dose of 200 ng PTX (Sigma-Aldrich) was injected intraperitoneally at 30 min before antigen injection.
At 13 days after immunization, a single cell suspension was prepared from the draining lymph nodes and spleens of EAU mice and applied to nylon wool columns. The nonadherent cells were collected as T cells, whereas the adherent cells were separated from the columns and served as antigen-presenting cells (APCs) after irradiation of 37 Gy. T cells (1 × 107/well) were then incubated with APCs (1 × 107/well) for 48 h in the presence of 10 μg/mL IRBP 1–20 and 10 ng/mL interleukin (IL)-23 (R&D Systems, Minneapolis, MN). The T cells were then separated by gradient centrifugation using Ficoll (GE Healthcare Life Sciences, Freiburg, Germany). For induction of adoptively transferred uveitis, the live T cells were injected intraperitoneally into naive C57BL/6 mice (5 × 106 cells/mouse).
MSC treatment protocol and clinical evaluation
The mAD-MSCs were pretreated with 100 μM alpha,beta-methylene adenosine-5′-diphosphate (APCP) (a CD73 inhibitor) (Sigma-Aldrich) overnight before injection into EAU mice. The EAU mice were randomly divided into three groups. The two mMSC-treated groups received a single intravenous injection of either 1 × 106 plain mMSCs or mMSCs pretreated with APCP at disease onset (10 or 11 days after transfer), whereas the control group received the same volume of PBS injection. Mice were examined three times a week for clinical signs of uveitis by indirect fundoscopy. The incidence and severity of inflammation were recorded in a blinded manner as grades of 0–4 according to Caspi et al. [16]. Six mice in each group were followed up for 60 days after transfer for clinical and histological evaluation and electroretinography (ERG) assessment, whereas six mice in two mMSC-treated groups and nine mice in control group were kept for 21 days before sacrifice for T-cell proliferation assay.
Histology
For histopathological evaluation, the eyeballs were collected on day 60 after transfer and embedded in paraffin after fixation and dehydration. Four-micrometer sections were cut and stained with hematoxylin and eosin. The degree of retinal inflammation and damage was assessed in a blinded manner on a scale from 0 to 4 using the criteria of Caspi et al. [16].
Electroretinography
To evaluate retinal function, dark-adapted ERG was recorded (IRC, Chongqing, China) on days 25, 35, 45, and 60 after transfer. The mice were dark adapted overnight before recording, and the procedure was performed under dim red light. Ketamine (77 mg/kg) was used for systemic anesthetization and 0.5% Alcaine drops were used for ocular surface anesthetization. Pupils were dilated as described previously. For ERG recording, a gold ring electrode was placed in contact with the cornea, and the reference and ground electrodes were placed on the tail and back of the neck, respectively. The amplitude of a-waves was measured by the range between the baseline and the trough of a-waves, whereas the amplitude of b-waves was measured from the trough of a-waves to the peak of b-waves.
MSC viability test
To verify the toxicity of APCP pretreatment on mMSCs, annexinV-7AAD (BioLegend) staining followed by flow cytometry analysis was performed. MSCs were treated with APCP (100 μM) or complete medium only (control) for 12 h. Aliquots of 5 × 105 cells were then incubated with annexin V-7AAD for 15 min at room temperature. Data collection was performed on a FACSCalibur flow cytometer (BD Biosciences, San Jose, CA) and analyzed using flow cytometry software (FlowJo). The acquisition and analysis were gated on MSCs, based on characteristic properties of the cells in forward and side scatter.
T-cell proliferation assay
To assess the suppressive effects of mMSCs and APCP-pretreated mMSCs on T cells in vivo, T cells and APCs from lymph nodes and spleens of three mice from each group were prepared on day 21 post-transfer as described previously. T cells (4 × 105/well) were seeded in 96-well plates (Corning, Corning, NY) and treated with various doses of IRBP 1–20 (0, 1, 10, and 30 μg/mL) in the presence of APCs (1 × 105/well) in a total volume of 200 μL. Cells were cultured for 3 days at 37°C with 5% CO2 in RPMI 1640 (Gibco) supplemented with 10% fetal bovine serum, 2 mM glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin, and 50 μM β-mercaptoethanol (ICN Biomedicals, Irvine, CA). T-cell proliferation was then measured by a BrdU ELISA kit (Roche, Indianapolis, IN) according to the manufacturer's instructions.
To measure the direct suppressive effects of mMSCs and APCP-pretreated mMSCs on uveitic T-cell proliferation in vitro, T cells and APCs from lymph nodes and spleens of six untreated EAU mice were isolated and seeded in 96-well plates. The mMSCs or APCP-pretreated mMSCs were then added at various ratios (mMSCs/T cells: 1:5, 1:10, and 1:20). The cells were then cultured as described previously. To evaluate the effect of CD73 on T-cell proliferation, several soluble factors, including a CD39 inhibitor (100 μM sodium polyoxotungstate 1 [POM-1], Tocris Bioscience), APCP (100 μM, Sigma-Aldrich), and a selective and competitive antagonist of the A2A adenosine receptor (25 μM SCH58261, Tocris Bioscience), were added to the coculture system. Proliferation was also determined by the BrdU ELISA kit as described previously.
To further verify whether CD73 on hMSCs can exert immunosupressive effect on human peripheral blood mononuclear cells (PBMCs), we isolated PBMCs by gradient centrifugation using Ficoll. Aliquots of 2 × 105 cells were seeded in a 96-well plate precoated with anti-CD3 antibody (2 μg/mL) at 4°C for 12 h, and cocultured with hMSCs or APCP-pretreated hMSCs at various ratios (hMSCs/human PBMCs: 1:5, 1:10, and 1:20) for 72 h, under the stimulation of 2 μg/mL of soluble anti-CD28 antibody. The proliferation test was also determined by the BrdU ELISA kit as described previously.
Changes in CD73 expression of T cells
T cells and APCs were prepared from untreated EAU mice, seeded in 24-well plates, and cultured with or without 10 μg/mL IRBP 1–20. The mMSCs, recombinant human transforming growth factor-β1 (TGF-β1) (2 ng/mL; PeproTech, Rocky Hill, NJ), IL-6 (50 ng/mL; PeproTech), hepatocyte growth factor (HGF) (50 ng/mL; PeproTech), IL-10 (20 ng/mL; PeproTech), and mMSCs together with an anti-TGF-β1 (50 ng/mL; PeproTech) were also added to examine the effect of mMSCs and mMSC-secreted cytokines on the expression of CD73 in activated T cells. The CD73 expression on T cells was detected under different conditions by flow cytometry. The change of CD73 expression on human PBMCs was also detected by flow cytometry after 24 h of coculture with hMSCs, under the stimulation of anti-CD3 and anti-CD28.
Malachite green assay for determination of ecto-nucleotidase activity
The mMSCs were washed with phosphate-free buffer and then seeded in 96-well plates (1 × 105/well). 5′-AMP (Sigma-Aldrich) diluted in phosphate-free buffer was added to six wells at a final concentration of 50 μM, and in three wells of which we added 100 μM APCP. In blank control wells, mMSCs were incubated in phosphate-free buffer in the absence of 5′-AMP and APCP. After 30 min of incubation at room temperature, malachite green colorimetric assay was performed with a SensoLyte kit (AnaSpec, Inc., Fremont, CA). The absorbance was measured at 630 nm.
Measurement of adenosine production by high-performance liquid chromatography
To detect adenosine in cell culture supernatants, mMSCs were incubated in 24-well plates (1.5 × 105/cm2) for 1 h at 37°C in the presence of 50 μM 5′-AMP in phosphate-free incubation buffer with or without 100 μM APCP. Control wells contained mMSCs alone. Supernatants were collected, evaporated under N2, resuspended in 200 μL H2O bidest, and analyzed by high-performance liquid chromatography (HPLC). Chromatographic analysis was performed on a Shimadzu LC-MS2020 instrument. A SPD 20AV detector was used with a Venusil XBP C18 5 μm column (Bonna-Agela Technologies, Tianjin, China). Adenosine was measured isocratically with acetonitrile and water as the mobile phase at a flow rate of 0.8 mL/min. The absorbance was measured at 260 nm. The retention time was 9 min. Class VP software was used for system control and data analysis.
Statistical analysis
GraphPad Prism software (version 5.01) was used for all statistical analyses. Data are expressed as means ± standard deviation. EAU clinical scores were assessed by repeated measures analysis of variance (ANOVA) using mixed models. One-way ANOVA was applied for three or more sets of data. P < 0.05 was considered significant.
Results
MSC characterization
Flow cytometric analysis revealed that mMSCs lacked expression of hematopoietic markers CD34, CD45, and CD11b, but highly expressed MSC-associated markers such as CD29, CD44, CD73, CD90, CD105, and Sca-1 (stem cell antigen-1) (Fig. 1A). The cells could be induced to differentiate into adipocytes, osteocytes, and endothelial cells under appropriate conditions (Fig. 1B1–B4). We also detected CD39 expression on mMSCs (Fig. 1A). In addition, hMSCs from umbilical cord, adipose tissue, and bone marrow showed very low levels of CD39 expression (2.4%, 3.2%, and 2.1%, respectively).

Characterization of mMSCs.
MSC treatment ameliorates EAU through CD73 expression
Early signs of uveitis were found on day 10 or 11. Severe illness had developed from days 18 to 24, and then the inflammation diminished gradually by day 60 (Fig. 2, black curve), while leaving persistent damage of the retinal structure (Fig. 3A). A significant decrease in disease scores was observed following mMSC treatment (Fig. 2, blue curve), whereas blocking the activity of CD73 on mMSCs by APCP significantly decreased the therapeutic effect of mMSCs on the progression of EAU (Fig. 2, red curve). In line with the clinical observations, histopathological examinations of retinal sections on day 60 revealed that mMSC treatment dramatically reduced the retinal damage with less inflammatory cell infiltration. This therapeutic effect resulted in a near normal retinal structure compared with the severe retinal injury characterized by striking infiltration of inflammatory cells and retinal disorganization in the control group (Fig. 3A–C). As expected, histopathological scores were higher in the APCP–mMSC-treated group than in the mMSC-treated group, indicating that APCP pretreatment significantly attenuated the therapeutic efficacy of mMSCs (Fig. 3D).

MSCs attenuate the clinical severity of EAU in mice through CD73. The mMSCs or mMSCs pretreated with APCP were administered at day 10 after transfer (disease onset). Overall trend of the disease course demonstrated that mMSCs significantly decreased disease scores, whereas blocking the activity of CD73 on mMSCs by APCP dramatically impaired the therapeutic effects of mMSCs on the progression of EAU. Black asterisks indicate the significant differences between control group (black curve) and mMSCs-treated group (blue curve), red asterisks for control group (black curve) and APCP–MSC-treated group (red curve), and blue asterisks for APCP–MSC-treated group (red curve) and MSC-treated group (blue curve). EAU clinical scores were assessed by repeated measures ANOVA, using mixed models. Values are expressed as the mean ± SD of six mice (12 eyes) per group. *P < 0.05. ANOVA, analysis of variance; EAU, experimental autoimmune uveitis; SD, standard deviation.
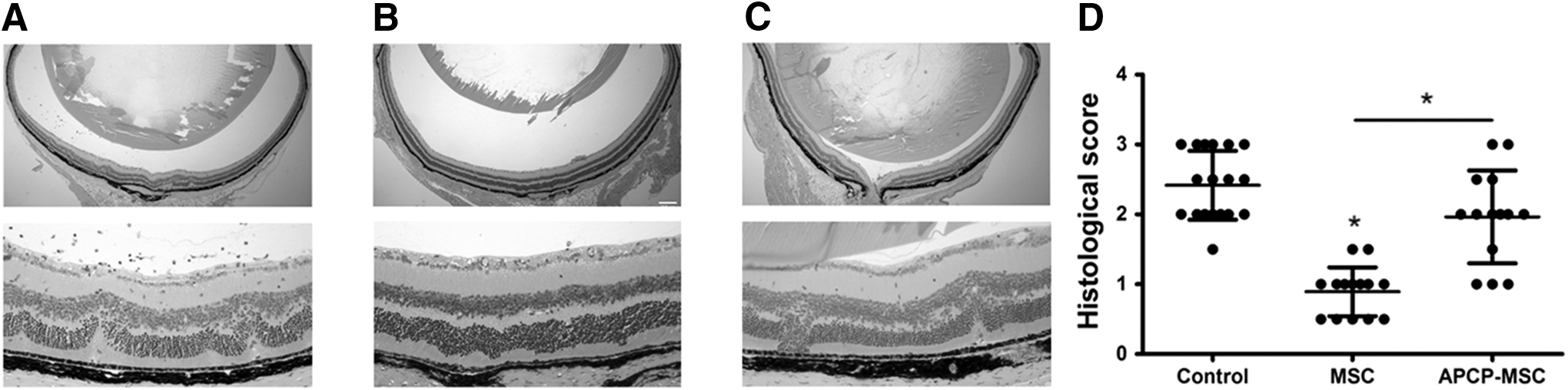
Histological changes of the retina on day 60. Sections of the retina were stained with hematoxylin and eosin, and evaluated for histological damage. Representative retinal histological features are shown. Compared with severe retinal injury characterized by striking infiltration of inflammatory cells and retinal disorganization in the control group
Both the therapeutic effect of mMSCs and the favorable role of CD73 were further confirmed by ERG evaluation. Reductions in dark-adapted a- and b-waves and oscillatory potential were detected on days 25, 35, 45, and 60 post-transfer in EAU mice (Fig. 4). The retinal function of mice in the mMSC-treated group was much better than that of mice in the control group, whereas blocking the activity of CD73 significantly reversed the protective effect of mMSCs on retinal function.
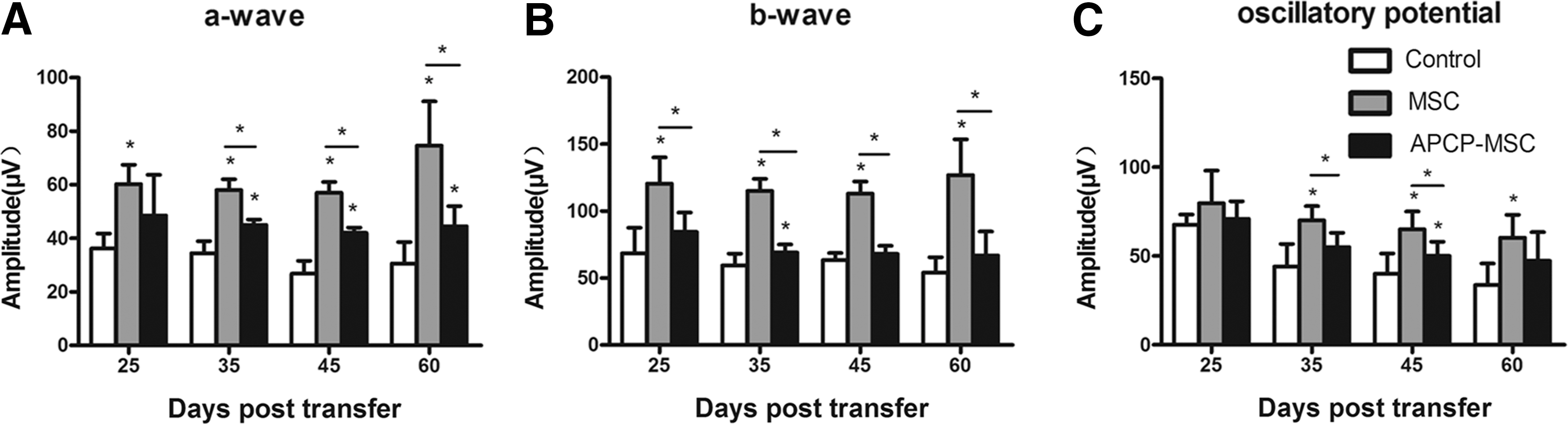
ERG responses after dark adaptation. The retinal function of EAU mice was monitored and followed up at the indicated time points by ERG. a-Wave
Modulation of T-cell proliferation by AD-MSCs through CD73
Before exploring the effect of CD73 on T-cell proliferation, we tested whether APCP is toxic to mMSCs by annexin V-7AAD staining. As shown in Fig. 5A, mMSC viability was not impaired by APCP pretreatment. In T-cell proliferation experiments, the proliferation indexes of T cells from control mice were positively correlated with the increasing concentrations of IRBP 1–20 (Fig. 5A). The mMSC treatment resulted in lower proliferation of T cells, whereas the proliferative potential was partially recovered in T cells following APCP-pretreated mMSC treatment. To verify the direct immunosuppressive effect of mMSCs on uveitic T cells in response to antigen stimulation in vitro, we cocultured these cells in the presence of 30 μg/mL IRBP 1–20 and found that mMSCs (mMSCs/T cells: 1:5) significantly inhibited the proliferation of T cells (Fig. 5B, column C), whereas APCP-pretreated mMSCs had a significantly lower suppressive effect on T-cell proliferation (Fig. 5B, column D).

MSCs modulate T-cell proliferation through CD73. Human and mouse T-cell proliferation assays were performed as described in the Materials and Methods section.
To further investigate the role of the CD39/CD73/adenosine pathway in T-cell proliferation, we repeated the proliferation experiment under different conditions. A burst of T-cell proliferation was observed when APCP was directly added to the T-cell culture system without mMSCs (Fig. 5B, column E), suggesting that CD73 on T cells also plays an important role in the inhibition of T-cell proliferation. Most reports consider the A2A adenosine receptor to be related to immunosuppression. When SCH58261 (an A2A adenosine receptor antagonist) was added to the T-cell culture system without mMSCs, a remarkable T-cell response similar to that mediated by APCP was observed by blockade of adenosine from binding to its immunosuppressive receptor (Fig. 5B, column G). Although CD39 was not highly expressed on our cultured MSCs, it was expressed on activated human PBMCs and mouse T cells (Fig. 5C, D). Similar to SCH58261 and APCP treatment, POM-1 (CD39 inhibitor) treatment also led to potent T-cell activation (Fig. 5B, column I). When T cells were cocultured with mMSCs under the latter three conditions (columns F, H, and J in Fig. 5B), the proliferative ability of T cells was decreased but still high. These results indicated that the inhibitory capacity of mMSCs was, in part, dependent on the function of CD73. By collaborating with CD39 on T cells, CD73 on both mMSCs and T cells participated in the suppression of T-cell proliferation, which might be mediated by adenosine through the A2A receptor. The suppressive ability of mMSCs was not fully restrained after CD39–CD73–adenosine–A2A pathway blockade, probably because of other immunosuppressive mechanisms.
The inhibitory effect of CD73 in hMSCs on human PBMCs was also clarified by T-cell proliferation experiments in vitro. The hMSCs showed a significant inhibitory effect on PBMC proliferation induced by CD3 and CD28 stimulation in a dose-dependent manner, whereas APCP-pretreated hMSCs exhibited less suppressive effects (Fig. 5F).
CD73 may contribute to immunosuppression through adenosine production
A colorimetric malachite green assay was used to measure the ability of CD73 on mMSCs to generate inorganic phosphate from exogenous 5′-AMP in the absence or presence of APCP, whereas the capacity of CD73-expressing mMSCs to produce adenosine from 5′-AMP substrate was detected by HPLC. As expected, APCP significantly inhibited phosphate (Fig. 6A) and adenosine (Fig. 6B) production by the cultured mMSCs following the addition of 50 μM 5′-AMP. These observations implied that CD73 on mMSCs may facilitate immunosuppression through adenosine production.
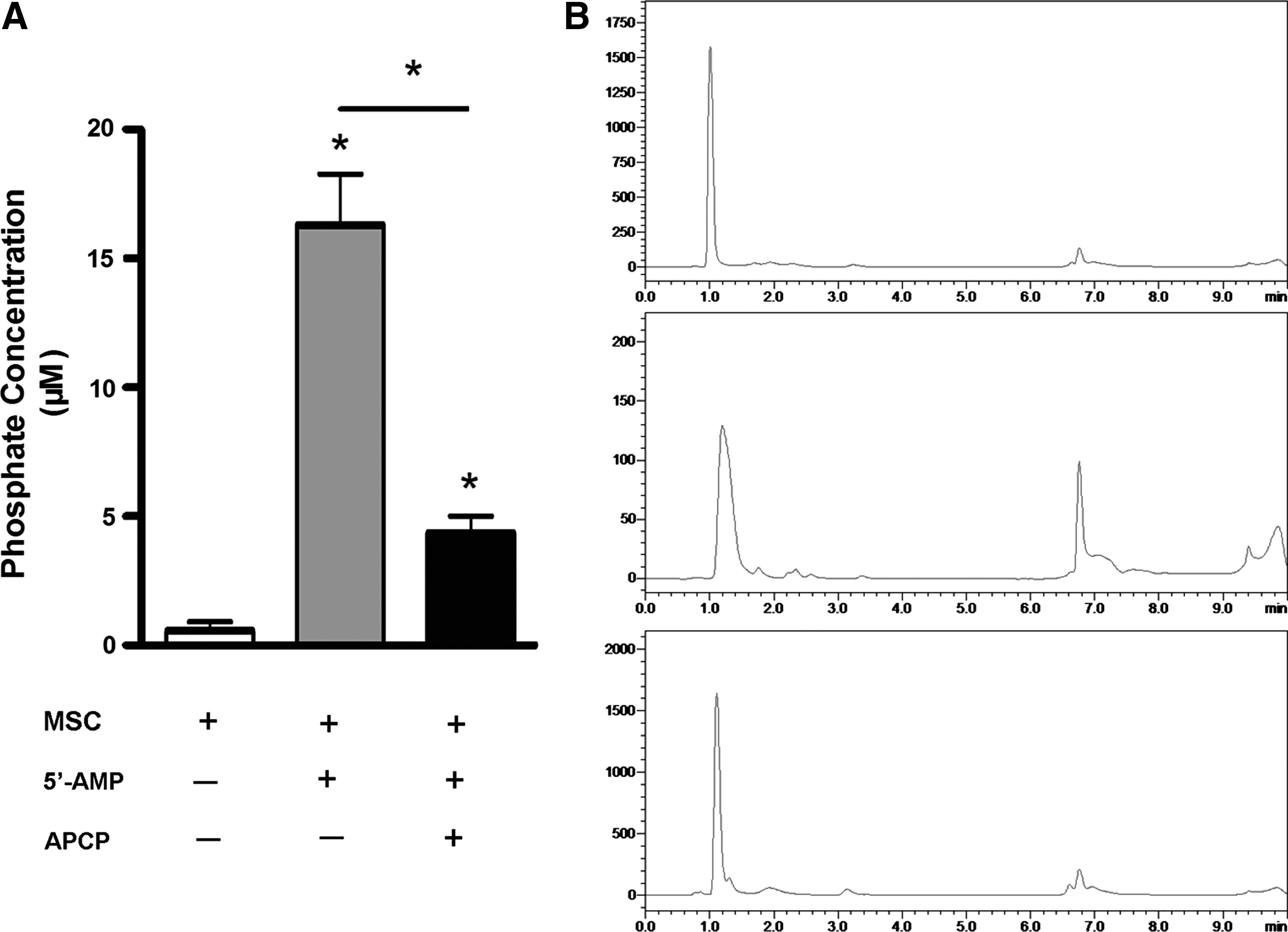
CD73 on MSCs hydrolyzes exogenous 5′-AMP to adenosine.
MSCs upregulate CD73 expression on CD4+ T cells in vitro
Next, to further explore the involvement of CD73 in the interaction of MSCs and T cells, we assessed changes in the expression of CD73 on T cells in the presence of MSCs. Flow cytometric analysis revealed that the expression level of CD73 on CD4+ T cells decreased upon IRBP 1–20 stimulation, whereas coculture with mMSCs prevented CD 73 downregulation and actually enhanced CD73 expression on T cells (Fig. 7A). Among the tested cytokines, only the addition of TGF-β1 to the culture system of IRBP 1–20-activated T cells demonstrated a clear influence on CD73 expression by reversing the reduction. In contrast, a neutralizing anti-TGF-β antibody antagonized the induction of CD73 in the coculture system of mMSCs and T cells. Other cytokines produced by mMSCs, including IL-6, IL-10, and HGF, did not show a preventive effect on the downregulation of CD73 expression induced by IRBP 1–20 (Fig. 7B). Therefore, it could be hypothesized that mMSC-secreted TGF-β1 promotes upregulation of CD73 on T cells, which might further enhance T-cell inhibition.
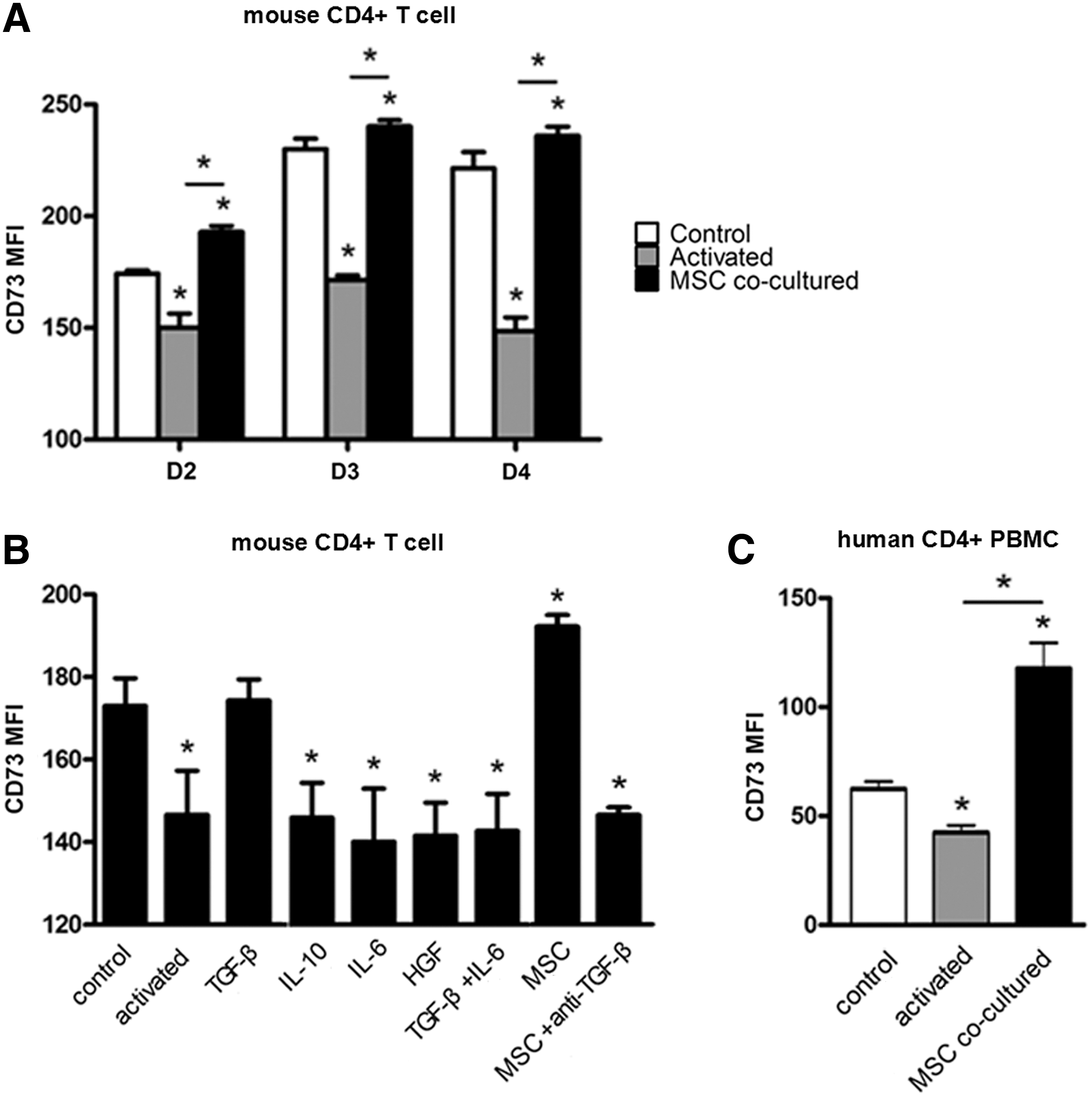
MSCs increase CD73 expression on mouse CD4+ T cells and human PBMCs.
The hMSCs were also tested for their ability to induce upregulation of CD73 on CD4+ T cells. As shown in Fig. 7C, CD3/CD28-induced T-cell activation led to downregulation of CD73 expression on T cells. Nevertheless, the expression of CD73 was dramatically increased by coculture with hMSCs (Fig. 7C).
Discussion
Our previous study demonstrated that rat bone marrow MSCs ameliorate both monophasic and recurrent EAU in rats by downregulation of Th1/Th17-mediated responses, restoring the balance between Th17 and Treg cells (regulatory T cells) and suppressing the function of APCs [17 –19]. In this study, to clarify the role of CD73 in the therapeutic effect of mMSCs, we used a chronic EAU model in mice induced by adoptive transfer of antigen-specific T cells, which is characterized by a chronic inflammatory course and retinal damage. We observed that mMSCs effectively controlled the development and severity of EAU and protected the retinal structure and function. However, APCP pretreatment of mMSCs significantly weakened their therapeutic efficacy to suppress the progression of EAU.
CD73 is an important marker of MSCs and also serves as an ectonucleotidase on the surface of a variety of cell types such as T cells, B cells, and myeloid cells [20]. CD73 catalyzes dephosphorylation of 5′-AMP to generate adenosine, thereby exerting immunosuppressive effects. Consistent with the clinical results, we found that pretreatment of MSCs with APCP, a CD73 inhibitor, mostly abrogated the ability of MSCs to suppress both human and mouse T-cell proliferation in vitro. Interestingly, a burst of T-cell proliferation was found when APCP was directly added to the mixed lymphocyte reaction without mMSCs, suggesting a key inhibitory role of CD73 on T cells. These results indicated that CD73 expressed on both mMSCs and T cells is important to maintain an environment that inhibits T-cell activation. Important regulatory roles of CD73 in the immune system have also been reported by several studies, involving various cell types, including lymphocytes [21 –24], monocytes/macrophages [25,26], dendritic cells [27], natural killer cells [28], neutrophils [29,30], and endothelial cells [31,32]. Recently, Romio et al. reported that ablation of CD73 in T cells and inhibition of CD73 with APCP increased the release of proinflammatory cytokines and T-cell proliferation [33]. Smyth et al. also demonstrated that CD73 expression and adenosine generation mediate the immunosuppressive function of exosomes released by Treg cells [34]. In our study, the proliferative ability of T cells was still potent but decreased when mMSCs were added to the mixed lymphocyte reaction with APCP, indicating that mMSCs still exert an inhibitory effect partially by other mechanisms after blocking CD73.
Lee et al. demonstrated that, when cocultured with human bone marrow-derived MSCs, human Th17 cells express high levels of CD73 [13]. Natural killer cells were also reported to acquire CD73 expression upon exposure to human umbilical cord-derived MSCs [28]. In this study, we confirmed that CD73 expression on both human and mouse CD4+ T cells was upregulated by coculture with MSCs. It has been reported that CD73 expression on activated CD4+ T cells can be induced by Treg-cell-secreted TGF-β in a manner independent of Foxp3 expression [35], whereas both IL-6 and TGF-β have been proved to be essential for CD73 expression during the differentiation of Th17 cells [24]. IL-6 activates signal transducer and activator of transcription 3 (Stat3), a promoter of ectonucleotidase expression, whereas TGF-β downregulates growth factor-independent protein 1 (Gfi-1), a repressor of ectonucleotidase expression. However, in our study, only TGF-β1 alone enhanced CD73 expression on CD4+ T cells, and neither IL-6, IL-6 plus TGF-β1, nor other mMSC-secreted cytokines could evoke upregulation of CD73. Furthermore, an anti-TGF-β1 antibody antagonized the induction of CD73 in the coculture system of mMSCs and T cells. These results suggest that mMSCs upregulate CD73 expression on T cells through TGF-β1, which might further enhance autoimmune inhibition.
CD39 is a cell surface ectonucleotidase that was first found on Treg cells as an immunosuppressive marker by Deaglio et al. [21], and has been recently described in several other cell types, including T cells, B cells, monocytes, and neutrophils [36]. It is widely believed that CD39 catalyzes dephosphorylation of ATP/ADP into 5′-AMP, initiating the first step of converting a proinflammatory environment into an anti-inflammatory state. Recent studies have reported that CD39 is also highly expressed on cultured MSCs (>50%), and pretreatment of MSCs with the CD39 inhibitor POM-1 downregulates the immunosuppressive functions of MSCs [12 –14]. Notably, we found that MSCs derived from mouse adipose tissue as well as hMSCs from adipose tissue, umbilical cord, and bone marrow expressed low levels of CD39 (2.1%–3.2%). This difference might be attributed to the different tissue sources, species, and culture methods. Although CD39 was not highly detected on our cultured MSCs, we found CD39 on activated T cells (human PBMCs, 13.45%; mouse T cells, 69.67%), and blocking CD39 in a mixed lymphocyte reaction by adding POM-1 led to a remarkable increase in the T-cell proliferation index. Our results indicate that, without CD39 activity, CD73 exerts little effect on T-cell proliferation, and CD39 expression by T cells ensures the integrity of the CD39/CD73/adenosine pathway in mMSC and T-cell interactions.
Adenosine accumulation in extracellular spaces elicits immunosuppressive effects by binding to several type 1 purinergic (adenosine) receptors, among which the A2A adenosine receptor is predominant on lymphocytes. Stimulation of the A2A adenosine receptor on activated T cells results in the inhibition of proinflammatory cytokine production and effector reactions [37,38]. Of note, Naganuma et al. showed that the A2A adenosine receptor plays a critical role in alleviation of T-cell-induced experimental colitis by inhibiting the expression of proinflammatory cytokines [39]. Here, we found remarkable T-cell proliferation by blocking the binding of adenosine to the A2A adenosine receptor with SCH58261.
Conclusions
In conclusion, we have first demonstrated that MSCs inhibit the autoimmune response partially through the cooperation of CD73 expression on MSCs and CD39/CD73 coexpression on T cells. Moreover, MSCs promote the expression of CD73 on activated T cells, which in turn might enhance autoimmune suppression. These findings further support the notion that MSCs have the potential to treat autoimmune diseases by creating an immunosuppressive microenvironment around T cells through multiple mechanisms. The clarification of the immunomodulatory mechanism of MSCs will further provide basis for developing a new strategy to improve immunosuppressive ability of MSCs.
Footnotes
Acknowledgments
This research was supported by grants from the National Natural Science Foundation of China (81371005, 81428012), Tianjin Municipal Science and Technology Commission (15JCZDJC35600), National Basic Research Program of China (2013, CB966904), and Research to Prevent Blindness, Inc., New York, New York.
Author Disclosure Statement
No competing financial interests exist.