
Abstract
Generation of gametes derived in vitro from pluripotent stem cells holds promising prospects for future reproductive applications. Indeed, it provides information on molecular and cellular mechanisms underlying germ cell (GC) development and could offer a new potential treatment for infertility. Great progress has been made in derivation of gametes from embryonic stem cells, despite ethical issues. Induced pluripotent stem cells (iPSCs) technology allows the reprogramming of a differentiated somatic cell, possibly emanating from the patient, into a pluripotent state. With the emergence of iPSCs, several studies created primordial GC stage to mature gamete-like cells in vitro in mice and humans. Recent findings in GC derivation suggest that in mice, functional gametes can be generated in vitro. This strengthens the idea that it might be possible in the future to generate functional human sperm and oocytes from pluripotent stem cells in culture.
Introduction
I
Significant progress has been realized in the treatment of human infertility. However, artificial reproduction techniques (ARTs) cannot offer an answer for all causes of infertility. Knowledge concerning early stages of gamete development is rather poor. Indeed, therapeutic methods are missing, particularly for gametogenesis abnormalities. A human model of in vitro gametogenesis from stem cells would have a significant impact on evolution of fundamental knowledge and on the development of adapted therapies. Thus, obtaining male and female gametes in vitro could modify current ARTs and provide laboratories with a plentiful source of mature and functional oocytes and sperm [2].
Embryonic stem cells (ESCs) are derived from the inner cell mass (ICM) of a blastocyst stage embryo [3]. These cells exhibit pluripotency and self-renewal properties, and can give rise during development to the three embryonic germ layers allowing potential use in therapeutics, transplantation, drug testing, and early embryogenesis study [3]. Stem cells include ESCs as well as fetal and adult stem cells.
Up to the morula stage [3 days postcoitum (dpc), in humans], embryonic cells are totipotent, meaning each cell is theoretically capable of generating a whole organism. At the blastocyst stage (5 dpc in humans), ICM cells segregate into epiblast and hypoblast. The epiblast gives rise to the three primary germ layers (ectoderm, endoderm, and mesoderm). The hypoblast forms the yolk sac. Cell lines of pluripotent and self-renewal ESCs are derived from the ICM, while induced pluripotent stem cells (iPSCs) arise from the reprogramming of differentiated adult or fetal somatic cells.
The iPSCs were obtained in mice [mouse induced pluripotent stem cells (miPSCs)] and in humans [human induced pluripotent stem cells (hiPSCs)] by inducing overexpression of defined genes such as POU class 5 homeobox 1 (POU5F1 or Oct4), SRY-box 2 (Sox2), v-myc avian myelocytomatosis viral oncogene homolog (c-Myc), Kruppel-like factor 4 (gut) (Klf4), Nanog homeobox (Nanog), and Lin28 [4,5]. These genes are involved in pluripotence and cell proliferation.
In vivo gametogenesis is a complex process, including a succession of defined stages. First, the epiblast cells adopt a primordial germ cell (PGC) fate during the extraembryonic phase after receiving an induction signal from the surrounding extraembryonic tissues. Then, the PGCs migrate to the genital ridge finally to become gonocytes that will differentiate into gametes through meiosis and cellular differentiation.
Cells are first pluripotent, then evolve during migration under the influence of defined signals emanating from neighboring tissues. Thus, each stage matches a window of responsiveness reflected by the presence of specific inducers and by the appearance/disappearance of a set of specific or ubiquitous molecular markers (also named molecular determinants) in a context of cell niches (Fig. 1 and Supplementary Table S1; Supplementary Data are available online at
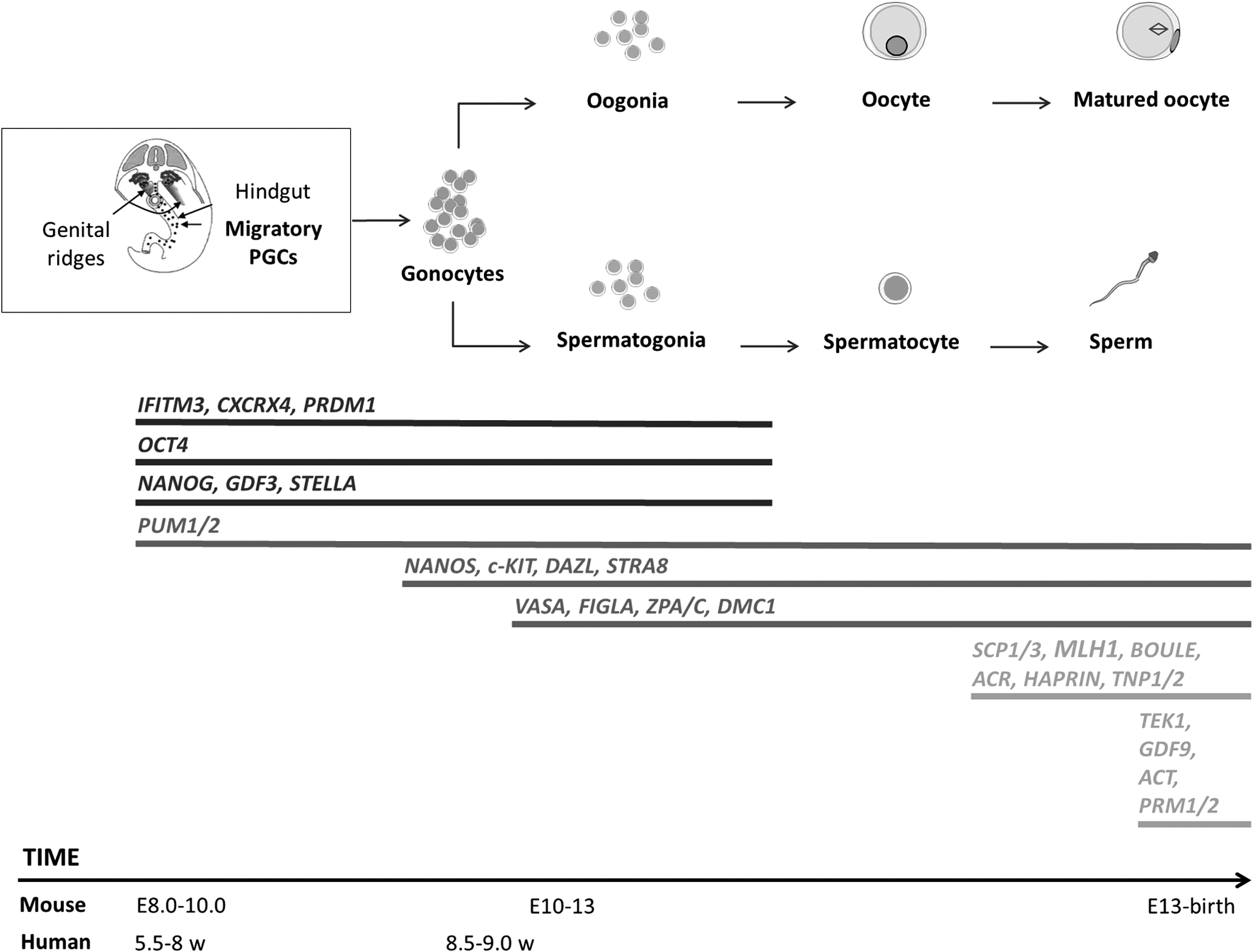
Molecular determinants' expression during germ line specification. Chronology of male and female germ line development events. Expression profile of premeiotic (black lines), pre-, and postmeiotic (dark gray lines) or exclusively meiotic (light gray lines) key genes (adapted from De Felici [8]). PGC, primordial germ cell; w, weeks.
Furthermore, germ line cells belong to an exceptional lineage because they become haploid through the process of meiosis, evolve into mature gametes, and perpetuate life. All these events are under the dependence of genetic (sexual chromosomes) and epigenetic (genomic imprinting) factors updating at each generation [7].
This review describes scientific advances toward germ cell (GC) differentiation from pluripotent hESCs and iPSCs, in the light of results from experiments performed in mice.
In Vitro Production of PGCs and Gonocytes
The formation of PGCs is a critical step in mammalian embryogenesis to ensure normal fertility. Indeed, their successful later differentiation leads to the obtaining of fertilizable oocytes and sperm. PGCs are indistinguishable from neighboring embryonic cells in the yolk sac wall, then acquire gradually germinal molecular markers also named molecular determinants, that is to say, genes expressing at a precise timing during germinal development.
To do so, cells activate a germinal specific program and inhibit a somatic program [8]. This double action comes along with a reactivation or maintenance of pluripotency [8]. In vivo, when PGCs reach the genital ridges, they differentiate into gonocytes and proliferate actively [8]. In vitro, it has been possible to obtain the expression of PGC and gonocyte markers mainly from human embryonic stem cells (hESCs) and mice embryonic stem cells (mESCs). The ESC-PGC and PGC-gonocyte transitions are not easy to distinguish because markers are often common. Results are described according to the most representative molecular determinants of each stage.
Clark et al. published a pioneering work in obtaining GCs from hESCs [9]. Differentiated cells expressed Deleted in Azoospermia-like (DAZL), a specific gene of germinal lineage, DEAD (Asp-Glu-Ala-Asp) box polypeptide 4 (DDX4 or VASA), whose expression appeared during embryoid bodies (EBs) formation after 14 days of culture, and other later markers [9].
Ever since, in vitro PGC differentiation has generated great interest as PGC specification is the first and crucial step in germ line development. Therefore, the crucial question arises of the relevance of the PGC-specific markers used. Several strategies used to enrich cultures with a PGC population leading to the identification of PGC-specific markers (Supplementary Table S2). These strategies modulate the expression level of specific genes by using gene transfer technologies or try to reproduce in culture PGC physiological environment.
PGC and ESC common molecular signature
hESCs share a panel of markers with human and/or mouse PGCs [8]. However, these genes showed differential expression patterns between ESCs and PGCs [8]. Some genes associated with pluripotency are critical in human primordial germ cell (hPGC) specification, proliferation, and survival. hPGCs are pluripotent as shown by the expression of specific markers, such as OCT4, NANOG, Developmental pluripotency-associated 3 pseudogene 2 (DPPA3P2 or STELLAR), Stage-specific embryonic Antigen 4 (SSEA4), or V-kit Hardy-Zuckerman 4 feline sarcoma viral oncogene homolog (C-Kit) [8,9]. In this way, PGCs can also be a source of pluripotent stem cells, called embryonic GCs [10]. Pluripotent gene expression is found in hESCs, in PGCs, and in gonocytes until the gonia stage.
Stage-specific GC marker genes [PR domain containing 1, with ZNF domain (PRDM1), Deleted in azoospermia (DAZ), DAZL, Pumilio homolog 2 (PUM2), Nanos homolog 1/3 (NANOS1/3), STELLAR, Piwi-like RNA-mediated gene silencing 2 (PIWIL2), Testis expressed 14 (TEX-14) among others] were found to be expressed in both hPGCs and hESCs [8,9,11]. PRDM1 expression is controlled by the bone morphogenetic protein (BMP)/Mothers Against DPP Homolog family member (SMAD) signaling pathway and is essential to ESC orientation toward a PGC identity in mice as in humans [12,13]. The highly conserved DAZ genes family contains DAZ [also named azoospermia factor c (AZFc)], DAZL, and Homolog of the human gene DAZ in Drosophila (BOULE) genes.
Mice knockout for Dazl are infertile because GCs are lacking [14]. BOULE null mice show a complete spermatogenic arrest at the round spermatid stage, before elongation [15]. In addition, 15% of human azoospermic or oligozoospermic patients carry a genetic microdeletion in the AZF regions. Using overexpression experiments on hESCs, Kee et al. showed DAZL is involved in PGC formation [16]. In addition, undifferentiated ESCs expressed other premeiotic genes such as PUM2, NANOS1, and C-KIT [9]. Therefore, given the similarity of gene expression profiles between PGCs and ESCs, it is very difficult to distinguish early GCs from undifferentiated ESCs. Meiotic and postmeiotic markers are more suitable markers in GC isolation from pluripotent stem cells.
PGC-specific molecular signature
Several markers began to be recognized as inescapable to establish the PGC phenotype allowing the distinction between pluripotent stem cells and early-stage GCs.
SSEA1/3/4 are expressed by migrating PGCs. The use of SSEA1 expression as a cell surface marker for identifying human in vitro-derived PGCs led to isolation of a subpopulation of hESC-enriched PGCs [17,18]. Another relevant cell surface marker is Chemokine (C-X-C motif) receptor 4 (CXCR4). The interaction of CXCR4 with its ligand Chemokine (C-X-C motif) receptor 4/ligand 12 (CXCL12) is necessary for PGC migration toward the primitive gonads [19]. Furthermore, CXCR4 is not expressed in ESCs and, until now, it has been one of the rare markers allowing the distinction between ESC and PGC stages [12].
VASA is also a valuable gene for the detection of GCs differentiated from pluripotent stem cells. VASA is regulated by DAZL and remains a significant marker of the PGC stage as seen previously in the work by Clark et al. [9,16]. Indeed, it has been shown that deficiency in VASA gene impaired mice male GC proliferation and differentiation [9,16]. Moreover, several studies showed that PGCs obtained from ESCs expressed the Developmental pluripotency associated 3 (DPPA3 or STELLA) gene [9,16,17]. The importance of Stella expression for germinal specification in mESCs was demonstrated by the generation of transgenic mice [20].
Finally, authors also reported the key role of the transcription factor SOX17 as a critical specifier of human PGC fate. SOX17 expression was significantly upregulated in generated primordial germ cell-like cells (PGCLCs) and, thus, was considered the earliest marker of PGCLCs in humans [21].
Besides, some markers are expressed exclusively by ESCs and iPSCs and not PGCs. For example, TRA-1-60 and TRA-1-81 (cell surface antigens) on human embryonic carcinoma cells and human pluripotent stem cell surfaces are used mainly as markers in identifying and isolating ESCs and iPSCs to discriminate them from differentiated PGCs [8].
Thus, PGC differentiation and isolation has been possible using GC gene and protein expression analysis and/or reporter construct tools [expression of Green fluorescent protein (GFP) under control of PGC-specific gene] [11,12,16 –18,21 –26]. All these works were directed toward generating PGCs from pluripotent stem cells in mice and humans.
Postmigratory and gonocyte-like cells, stages downstream of the PGC stage, were obtained with several culture strategies. Cells often expressed premeiotic markers [OCT4, NANOG, STELLA, Growth differentiation factor 3 (GDF3), PUM2, and NANOS], postmeiotic markers [Synaptonemal complex protein 1/3 (SCP1/3), MutL homolog 1, colon cancer, nonpolyposis type 2 (MLH1), Tektin 1 (TEKT1), and GDF9] or both (DAZL, VASA) [16,17,21 –24,26].
In conclusion, even if it was possible to isolate populations enriched with PGCs, the expression of specific markers of the PGC stage can also be associated with later markers of more advanced cell stages. This observation underlined the absence of a clear in vitro transition between the PGCs and the later meiotic steps. Furthermore, both genetic manipulation and environment modification strategies were successful in generating PGCs from ESCs or iPSCs in both species. However, the environment modification strategy seemed the most suitable since it strived to reproduce the in vivo environment in culture. Furthermore, iPSCs have already been genetically manipulated during the reprogramming process, so further genetic engineering could increase the risk of genetic instability even more.
Differentiation of Male and Female Advanced GCs and Mature Gametes
Gametogenesis is a general process allowing gamete formation in both male and female. In the testis, gonocytes proliferate considerably and gradually reach critical points of transition before initiating meiosis and sperm differentiation. This orientation and the successful progression of spermatogonia into sperm specification depend on local factors [BMP, retinoic acid (RA), stem cell factor (SCF), leukaemia inhibitory factor (LIF)] and, more particularly, on the cellular niche constituted by Sertoli and Leydig cells.
At puberty, hormonal factors dependent on the hypothalamus–pituitary axis [luteinizing hormone (LH) and follicle-stimulating hormone (FSH)] add to the paracrine factors. Leydig cells secrete testosterone, the main hormonal regulator of spermatogenesis, and regulate Sertoli cells that nourish, support, and protect developing GCs. These processes allow a normal progression of spermatogenesis [27]. Thus, the process of spermatogenesis in the seminiferous tubules is controlled by the regulation of both somatic and GCs, including physical interaction and secretion of biological factors [27]. These events follow one another according to a precise chronology that is necessary to reproduce in vitro.
In ovaries, gametogenesis is a discontinuous process during life. Meiosis begins during fetal life, is stopped before birth, restarts during puberty, stops again, then ends with fertilization. Gamete formation in the ovary is a cyclic phenomenon associated with an important apoptosis process. The relationship between oogenesis and folliculogenesis underlines the close and essential link between somatic cells and GCs. Folliculogenesis is under the control of locally produced factors [KIT/KIT ligand system; transforming growth factor ß (TGFß); GDF9; Anti-Mullerian hormone (AMH); androgens] and then becomes sensitive through granulosa and theca cells to pituitary gonadotrophin LH and FSH [28].
Spermatogenesis and oogenesis involve the formation of haploid cells from a diploid cell through meiosis. Nevertheless, two major differences remain. First, the number of gametes produced from every gametogenesis (only one for oogenesis in contrast to four for spermatogenesis). Second, oocyte production occurs only once a month during normal oogenesis from puberty to menopause, whereas sperm production begins at puberty and usually continues uninterrupted throughout adult life.
ESC-based research is rather limited, especially in humans due to ethical concerns involving human embryos' fate. Internationally, many jurisdictions have established specific legislation, ethical guidelines, and oversight bodies to govern stem cell and related embryo research [29]. Despite this, several groups were able to report ESC ability to differentiate into germ line cells [9,12,16,22,23,30 –46]. Some have successfully obtained male and female GCs at various stages of gametogenesis [spermatogonia, spermatids, sperm, oocytes with or without zona pellucida (ZP), follicle-like structures] by using different experimental strategies. The PGC or gonocyte intermediate stages were not a necessary intermediate step.
The development of the iPSCs technology has contributed enormously to the in vitro gamete derivation research. Recently, several research groups have shown that gametes could be produced in vitro from iPSCs [10,24,30,36,44,47 –56].
Male germinal differentiation from ESCs
Experiments in mice
Most of experimental strategies for ESC differentiation are based on EB formation. EBs are three-dimensional structures in suspension, in which ESCs acquire the ability to differentiate into a wide variety of cell types, including GCs. Toyooka et al. stated that mESC differentiation into male GCs was dependent on EB formation [22]. In this study, EB formation was induced by BMP4 addition. The BMP4 factor, produced by the extraembryonic ectoderm, plays a crucial role in GC formation in the proximal region of the epiblast. After coculture of mESCs with cells producing BMP4, and after 6–8 weeks of transplantation under a host testis capsule, they induced testicular tubule reconstruction and mature sperm in the light of the seminiferous tubules. Indeed, elongated spermatids were observed as shown by an anti-HSC70T (constitutively expressed in mice spermatids) staining.
RA is another inductor used for GC differentiation. Following this approach, Geijsen et al. obtained haploid male GCs by selecting a SSEA1+/Oct4+ population from mESC-derived EBs in the presence of RA [31]. Genomic imprinting erasure of the insulin-like growth factor 2 receptor (Igf2r) and H19 genes was observed as well as the expression of LHR, AMH, sex determining region Y (Sry), Acrosin (ACR), and Tripartite motif containing 36 (Trim36 or haprin) transcripts. After intracytoplasmic sperm injection (ICSI) of the obtained round spermatids into oocytes, blastocysts had developed. Diploidy was confirmed by fluorescent in situ hybridization [31].
Other authors showed that RA associated with testosterone allowed the expression of specific genes of the male germinal lineage [Stra8, Dazl, Activator of CREM in testis (Act) and Prm1] [33]. However, cell morphology was not described, and cell functionality was not evaluated.
Kerkis et al. reported in vitro production of early and late male gametes from XY mESCs using RA induction within nonadherent EBs. Interestingly, the authors were able to generate gamete-like cells without genetic manipulation or cell population selection. Produced GCs expressed germinal-specific genes [Mvh, Stella, Dazl, Piwil2, Tex14, Bmp8b, and ring finger protein 17 (Rnf17)], but also meiotic and postmeiotic male germinal-specific genes (Scp1, Scp3, Stra8, Haprin, and Acr) [34]. Early spermatids were observed with a meiosis I chromosome organization and a normal morphology. In this study, both types of presumptive gametes were obtained [34].
Other authors developed a culture system allowing PGC specification in mice, through the epiblast-like state. In mice, PGCs can be found in the ICM of preimplanted embryos and in the epiblast of postimplanted embryos. The epiblast-like cells (EpiLCs) transition obtained in PGC induction exhibits similar features to the transitional postimplantation epiblast, including the capacity to give rise to PGCs. Hayashi et al. reported for the first time viable healthy offspring from PGCLCs derived from mESCs and miPSCs [47]. To achieve this, a two-step differentiation protocol (the same one used for iPSCs) was undertaken, including the EpiLC transition step. In this way, PGCLCs produced were fully potent cells, since they differentiated into sperm. Fertilization of oocytes with the obtained sperm gave rise to healthy adult mice [47].
Finally, Nayernia et al. chose genetic manipulation strategies to facilitate in vitro GC isolation [32]. mESCs were transfected with the fusion genes Stimulated by retinoic acid gene 8 (Stra8)-EGFP or Protamine 1 (Prm1)-DsRed in the presence of RA allowing the development of haploid male gametes (early spermatids) [32]. To show that the resulting gametes were functional after fertilization by ICSI, 65 embryos at the two-cell stage were transferred into the fallopian tubes of a pseudopregnant mouse. Twelve births were obtained; however, all newborns died between day five and the fifth month after birth. A defect of genomic imprinting establishment is the most likely hypothesis to explain this neonatal mortality [32].
Experiments in humans
Based on the presence of meiotic markers (SCP1/3, MLH1, TEKT1), Clark et al. showed that hESCs, once gonocyte stage was obtained as described above, were capable of spontaneous differentiation into male GCs. However, no morphological criteria were evaluated [9]. Others strategies were based on the addition of various elements in the culture medium (RA, BMP, testicular extracts, laminin, spermatogonial stem cells [SSCs]) or target expression of PGC marker genes to induce hESC differentiation into male GCs [12,25,35,36].
West et al. reported that simply addition of mouse embryonic fibroblast feeders and basic fibroblast growth factor (bFGF) induced spontaneous GC differentiation. Hence, they extended the differentiation of a POU5F1+(OCT4+)/DDX4+(VASA+) population (reflecting pre/postmigratory PGC stages) and measured determining factors' expression of the spermatogonia stage (PUM2, DAZ1/4, and NANOS1) and meiotic markers (SCP3 and MLH1) [25] (Supplementary Table S2). Another team achieved haploid cells as round spermatid-like cells by cultivating hESCs with standardized mouse SSC culture conditions [36]. Bucay et al. also generated spontaneous emergence of PGCs expressing specific markers of male differentiation (ACR), including somatic-specific gene expression (FSHR, AMH, and SOX9) (Supplementary Table S2) [12].
These results displayed that spontaneous differentiation of ESCs into GCs was possible since PGCs and mature gametes were identified and isolated from cell aggregates. Thus, EB formation is often used as a mean to initiate ESC spontaneous differentiation. However, spontaneous differentiation is generally low and inefficient [33].
The addition of specific inductors for GC differentiation could actually mimic in culture the in vivo developmental processes. Using culture conditions enriched with various factors (BMP4, LIF, RA, or a preparation of neonatal testis), Aflatoonian et al. obtained round spermatids from hESCs after a selection of a SSEA1+/C-KIT+ PGC population [35]. Differentiated human cells expressed gradually early germinal (DAZL, VASA), meiotic (SCP3), and spermatic [Transition nuclear protein 1 (TNP1), PRM1/2] mRNA. In this study, round haploid spermatids with a compacted nucleus and a flagellum emergence were obtained. Simultaneously, the authors detected dihydrotestosterone and estradiol concentrations in the culture medium, two key hormones of spermatogenesis physiologically produced by surrounding somatic cells [35].
Thus, used together, EB induction and the addition of specific factors promoted hESC differentiation by simulating the physiological environment surrounding SSCs.
Concerning genetic manipulation strategies, VASA, DAZ, DAZL, and BOULE genes were mainly used by the authors. Following the overexpression strategy of DAZ, DAZL, and BOULE genes, Kee et al. succeeded in generating round spermatids and haploid male gametes (Supplementary Table S2) [16]. Progression through meiosis was initiated, and SCP3 staining patterns were observed consistent with the leptotene stage of prophase I [16]. In response to the overexpression of intrinsic regulators (VASA and/or DAZL), Medrano et al. also completed meiosis, as judged by haploidy and ACR expression of induced GCs [30].
Thus, strategies to induce male GCs in vitro used either forced expression of specific genes (DAZL, DAZ, BOULE, VASA) and/or the introduction of specific factors (BMP, RA, testosterone, bFGF, LIF, laminin-1, mSSC standard culture medium) into the medium.
Success was reported in the generation of haploid GCs from mice and hESCs [16,22,30 –32,34 –36,47]. In these articles, teams obtained spermatogenic cells matured from spermatid [16,22,31,34 –36] to sperm [47] stages. Cells expressed male germinal, meiotic, or postmeiotic genes [47]. EB formation can be used to promote spontaneous differentiation of mESCs into male GCs [12,22]. Available evidence indicated that the use of RA may be beneficial too [31 –35]. RA promotes meiosis initiation by stimulating Stra8 expression and may require the supporting cells for paracrine interaction leading to meiotic process. As showed by Silva et al., [33] the combination of RA and testosterone led to early spermatid-like cell formation.
In mice, transplantation of EB derived from ESCs succeeded in the reconstruction of seminiferous tubules with the formation of mature sperm in the lumen [22]. In humans, one recreated the physiological microenvironment in vitro by generating surrounding somatic cells [35]. Some studies further evaluated the fertilization potential of the generated male gametes [31,32,47]. In mice, ICSI experiments succeeded in embryo development [31,32] and gave birth to an offspring [32,47].
Nevertheless, a single study, which generated PGCs through an epiblast-like state, succeeded in generating healthy offsprings [47]. Individuals reached adult age and had normal fertility [47]. It is important to underline that the functionality of obtaining cells was never tested in human protocols.
Female germinal differentiation from ESCs
Fewer studies were undertaken for female germinal differentiation from pluripotent stem cells compared to male. As in male, two main strategies were used: overexpression of specific genes and/or the creation of an in vitro microenvironment most suitable for gamete differentiation.
Experiments in mice
In several studies, fluorescently labeled cells (transgene containing a specific gene promoter upstream of a fluorescent protein sequence) were used to isolate the initial steps of GC formation in vitro. In mice, Hübner et al. were the first to report the successful derivation of oocyte-like cells (OLCs) from mESCs in vitro. They used a monolayer culture system and introduced in mESCs a GFP construct under the control of the GC-specific Oct4 promoter [23]. Oct4 expression remained low during the last stages of oocyte maturation (Supplementary Table S2) [38].
Thus, after PGC induction, authors were able to observe meiotic GCs recognizable by a GFP−/c-kit−/Vasa+ population (Supplementary Table S2). Further differentiation led to OLC differentiation expressing Gdf9 and Scp3, surrounded with a visible ZP expressing the membrane proteins ZP2 and ZP3 (Supplementary Table S2). Follicle-like structures were also observed with a typical morphology and three-dimensional organization, expressed steroidogenesis enzymes, and secreted estradiol [23]. Stimulation by pregnant serum gonadotropin and human chorionic gonadotropin led to oocytes' expulsion and polar body-like formation, suggesting the complete achievement of the first meiotic division. Parthenogenetic embryos were then observed and developed up to the blastocyst stage [23].
Using the same protocol, Novak et al. described the formation of primary and primordial follicles from mESCs, but without clear meiotic chromosomal organization [41]. The synaptonemal protein SCP3 was detected, but its localization was atypical and its molecular partners were not detectable. So, the differentiation remained incomplete. An increase of estradiol concentration was however observed and signed the functionality of follicular-like somatic cells [41].
Using an eGFP reporter construct under the control of Gdf9 gene promoter, Salvador et al. reported the derivation of an OLC phenotype. Differentiation was based on the monolayer culture system or EB formation. Authors observed ZP with Zp3 expression, typical metaphase spindle with condensed chromosomes, and one polar body. Despite the fact that most of the Gdf9/eGFP-positive cells degenerated, authors obtained parthenogenetic zygotes at two- and four-cell stages [39].
Other studies focused on in vitro gamete induction with determined environmental factors only. Nicholas et al. combined the use of fluorescently labeled cells and a specific culture medium [37]. Indeed, they differentiated mESCs carrying Oct4-GFP reporter in culture with defined factors (BMP4, Kit-ligand, SCF, RA, R115866, bFGF). In this way, oocyte-like structures were obtained by secondarily selecting a SSEA1− population [37]. The authors showed that these cells initiated meiosis, but chromosomal localization of SCP3 remained partial. mESC-derived oocyte finally aborted in vitro [37]. These cells, transplanted under the renal capsule of a receiver mouse, were able to form primordial and primary-like follicles [37]. These data underlined that the niche context is required for follicle differentiation and maturation.
Lacham-Kaplan et al. and Qing et al. combined EB formation with coculture [40,42]. Lacham et al. obtained follicle-enclosed oocytes from mESCs cocultivated with testes of newborn male mice preparation medium [40]. Zp3 and Folliculogenesis-specific basic helix-loop-helix (Figα, an oocyte-specific transcription factor) expression was detected without visible ZP [40]. The benefic use of testicular cell condition medium in follicle-like generation from mESCs suggested that the male gonad microenvironment provided the necessary cues for oocyte development. However, authors failed to observe a ZP indicating that oocyte cells were at an early stage.
Qing et al. underlined the necessity of reproducing in vitro cellular interactions between germ and somatic cells by cocultivating mESC-derived PGCs with granulosa cells [42]. Obtaining oocytes expressed the oocyte-specific genes Figα, Gdf9, Zp1/3, and the meiosis marker Scp3 (well localized in the cytoplasm) with a typical morphology. However, the authors failed to fully distinguish the ZP [42].
Under the same experimental conditions described above, Kerkis et al. generated not only male but also female presumptive gametes from XY mESCs under RA induction [34]. This study represented significant advance as expression of meiotic (Scp1, Scp3, and Stra8) and female postmeiotic (Gdf9, Zp2, and Zp3) genes was detectable and also chromosomal reduction was observed in these ESC-derived gamete-like cells [34]. Hence, these observations indicated that GCs derived from mESCs progressed also into female gamete-like cell differentiation.
More recently, Wan et al. showed the potential of RA and oleanolic acid/or (OA) to induce mESC differentiation toward female germinal fate [43]. Indeed, RA and/or OA induced a significant upregulation of early GC markers (Gdf-9, Stra8, Mvh), Scp3, and oocyte-specific markers (Zp1, Zp2, and Zp3) [43]. These results suggested the OA could be also used alone as an inducer.
Hayashi et al. managed to develop PGCs as a precursor step for fully functional oocyte derivation [44]. Indeed, mESCs carrying germ line-specific fluorescent marker were induced successively into EpiLCs and PGCLCs. This two-step procedure appeared clearly efficient in generating PGC-like cells from mESCs. PGCLCs and embryonic gonadal somatic cells put together result in an in vitro reconstructed ovary. Then, the transplantation of this “artificial” ovary under ovarian bursa or kidney capsules of recipient mice allows follicle development, including oocyte-like growth with multiple layers of granulosa and theca cells [44]. However, the authors noticed abnormalities in the cumulus complex formation.
In vitro maturation (IVM) and in vitro fertilization (IVF) led to the development of two-cell embryos and further into blastocysts [44]. Two-cell embryos were transferred into mice and ended in a fully fertile offspring. However, some oocytes were unable to extrude the second polar body [44]. In a promising way, Hayashi et al. manifestly proved the full potency of ESC-derived OLCs.
Experiments in human
In humans, differentiation of female GC from hESCs was demonstrated. Performing spontaneous differentiation in vitro, ovarian follicle-like structures were obtained from hESCs [45]. The structures, obtained after 21 days of spontaneous culture, expressed VASA, STELLA, and GDF9, but quickly degenerated [45].
Chen et al. [45] and Aflatoonian et al. [35] concentrated their work in recreating local extracellular signals from the gonad microenvironment. This strategy was based on neonatal testicular factors utilization and led to in vitro OLC differentiation [35]. By using this type of culture on SSEA1+-selected hESCs, Aflatoonian et al. obtained OLCs as well as primordial follicle-like structures [35]. The expression of SCP3 and GDF9 was measured, but ZP essential proteins were not detected [35]. Using granulosa cells combined with RA cotreatment, female GC differentiation from hESCs was improved [46]. A subpopulation of cells Oct4-EGFP+ expressed VASA, GDF9, and SCP3, but the marker expression for mature oocyte ZP3 appeared to be low [46]. However, cells were able to develop into ovarian follicle-like structures, but putative oocytes did not progress through normal meiosis [46].
As in male, two main strategies were used: overexpression of specific genes and/or the creation of an in vitro microenvironment most suitable for gamete differentiation. In humans, promising progress was made in the generation of OLCs from ESCs by using spontaneous differentiation or coculture strategies. Results showed the potential of hESCs to differentiate into female GCs. Nevertheless, human oocyte-like differentiation in vitro was limited to the earliest stages. In contrast to humans, ESC-derived oocytes [42,43] and follicle-like structures [23,34,37,39 –41,44] were reported in mice. Some authors failed to observe nuclear meiotic progression [37,40,41,46].
However, oocyte-specific expression of Zp was successfully reported [23,34,39,43,44]. In mice, a parthenogenetic development of embryos was described [23,39] as well as a functional oocyte maturation [44]. Indeed, one study reported successful mESC-derived gamete cell fertilization resulting in a healthy offspring [44]. This underlines the difficulty in coordinating in vitro oocyte differentiation and meiosis. To date, fully mature oocytes derived from ESCs have not been obtained in humans.
In conclusion, fewer studies were undertaken for female germinal differentiation from pluripotent stem cells compared to male. Although it remains particularly difficult to isolate oocytes from ESCs due to the lack of suitable surface markers to distinguish GCs and oocytes, female gamete differentiation drafts from ESCs were possible [37]. Several protocols were undertaken and key factors regulating GC formation were identified. The efficiency of specific induction factors (RA, BMP, OA, R115866, neonatal testicular factors) [34,35,37,43] and coculture with gonad-specific cells or condition medium [40,42,46] were assessed for GC differentiation from ESCs.
Late GC stages were generated from both sexes, but the functionality was not always achieved. However, in mouse, Hayashi et al. generated successfully both functional oocyte and sperm in vitro and healthy offspring [44,47]. Nonetheless, this protocol implied a mouse transplantation step to recreate the in vivo microenvironment, making it difficult to consider any application in humans.
Male germinal differentiation from the iPSCs
Experiments in mice (Figs. 2 and 3)
Many approaches combined in vitro PGC differentiation to further in vivo maturation [11,47 –51] (Fig. 2). Niu et al. transplanted iPSCs under the kidney capsule of recipient mice. This led not only to large round cell differentiation expressing GCs and meiotic markers (Oct4, Dazl, Vasa, Stra8, and Scp3) but also to other cells expressing the sperm marker ACR [48]. This indicated that miPSCs had the capacity to differentiate into GCs. Once transplanted into seminiferous tubules, in vivo maturation obtained cells expressed Mvh and Stra8 and succeeded in repairing of the fibrosis testis [48].
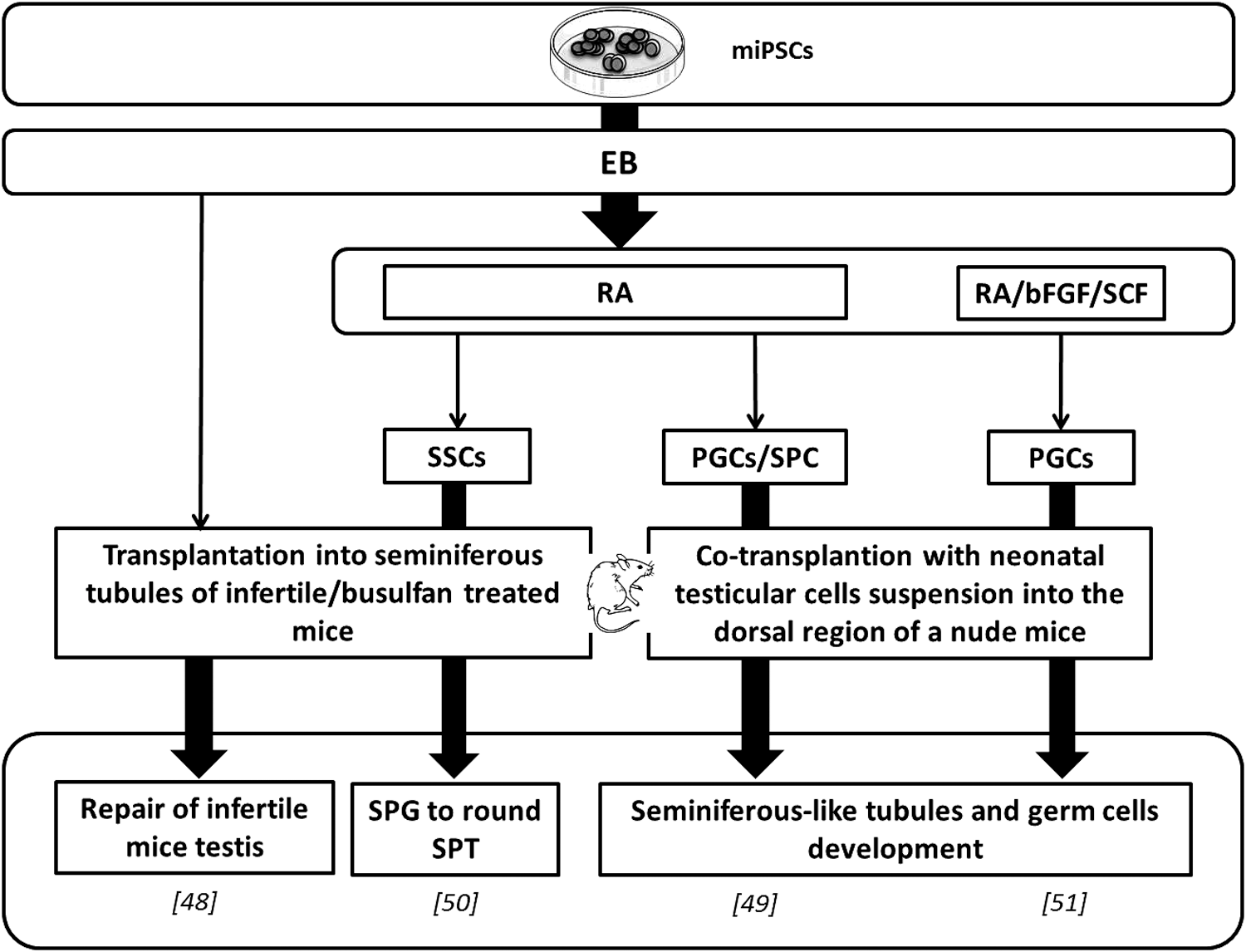
Main germ line differentiation protocols from miPSCs. Protocols reproducing in vivo conditions in culture to generate primitive germ cells and then transplanted obtaining cells in recipient mice for further in vivo maturation. bFGF, basic fibroblast growth factor; EB, embryoid body; miPSC, mouse-induced pluripotent stem cell; PGC, primordial germ cell; RA, retinoic acid; SCF, stem cell factor; SPC, spermatocyte; SPG, spermatogonia; SPT, spermatid; SSC, spermatogonial stem cell.
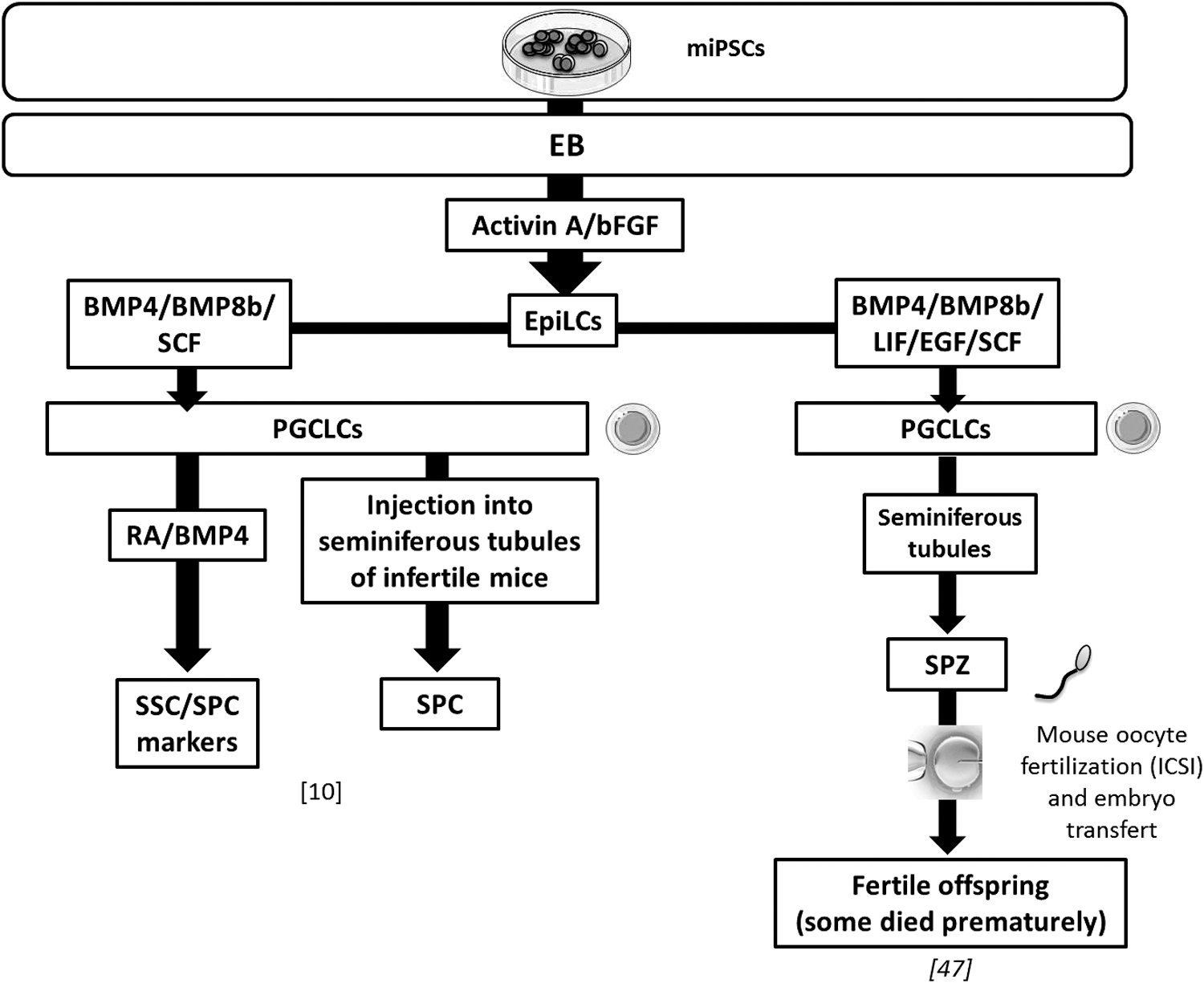
In vitro germ line differentiation protocols based on the EpiLC intermediate stage from miPSCs. One of the studies generated a healthy and fertile offspring. BMP4/8b, bone morphogenetic protein 4/8b; EGF, epidermal growth factor; EpiLC, epiblast-like cell; ICSI, intracytoplasmic sperm injection; LIF, leukemia inhibitory factor; PGCLC, primordial germ cell-like cell; SPZ, spermatozoa.
Few studies combined in vitro early GC-like stages acquisition with further in vivo steps. In vivo steps involved either transplantation into a sterile mice testis [50] or an ectopic cotransplantation with immature testicular cell suspension [49,51]. In this way, Zhu et al. reached SSC stages from miPSC EBs [expressing MVH, cadherin 1 (CDH1), and glial cell line-derived neurotrophic factor family receptor alpha 1 (GFRα1)], then, following transplantation, they managed to get spermatogonia to round spermatid stages (expressing MVH and SCP3). Thus, it underlined the real potential of miPSCs to differentiate into late male germinal GCs [50].
Yang et al. obtained not only in vitro RA induction cells expressing GC markers (Stella, Vasa, Dazl, and Stra8) but also spermatogonia (c-kit), meiotic, and postmeiotic (Scp3 and haprin, respectively) marker genes [49]. PGCLCs expressing Oct4, male germ line markers c-kit and Mvh were obtained in vitro by Cai et al., thanks to RA, bFGF, and SCF induction [51]. In both studies, the PGCLCs were then cotransplanted in addition to neonatal testicular cells into the dorsal region of nude mice allowing an ectopic reconstruction of seminiferous tubule structures containing spermatogenic cells (Fig. 2) [49,51]. This underlines the beneficial use of in vivo transplantation. However, further differentiation was not observed in reconstituted seminiferous tubules.
Two teams performed PGCLC differentiation from iPSCs through EpiLC induction [11,47] (Fig. 3). In one study, PGCLCs acquired spermatogenetic specification in vitro as in vivo by differentiating into spermatocytes expressing meiotic markers such as SCP3 [11]. In a functional study, haploid male GCs were generated and authors obtained fertile adult mice from PGCLCs derived from miPSCs, although some of the newborn died prematurely [47] (Fig. 3).
Few studies tested the iPSC ability to differentiate without any in vivo stage. Spontaneous differentiation of miPSCs into EBs and RA and/or testosterone induction for further differentiation strategies were efficient in GC differentiation from miPSCs [52]. This succeeded in haploid cell development [Outer dense fiber of sperm tails 2 (Odf2), Act, Prm1 transcripts] with a significant increase of Stra8 expression [52].
Experiments in humans (Fig. 4)
Since 2011, human iPSCs have been successfully used to differentiate into postmeiotic male GCs, including haploid cells [24,30,36,53]. Using conjointly BMP addition and overexpression of the specific regulator genes with or without BMP addition, authors obtained meiotic cells (with extensive synaptonemal complexes) and postmeiotic haploid cells with a similar pattern of ACR staining as observed in human spermatids [24,30].
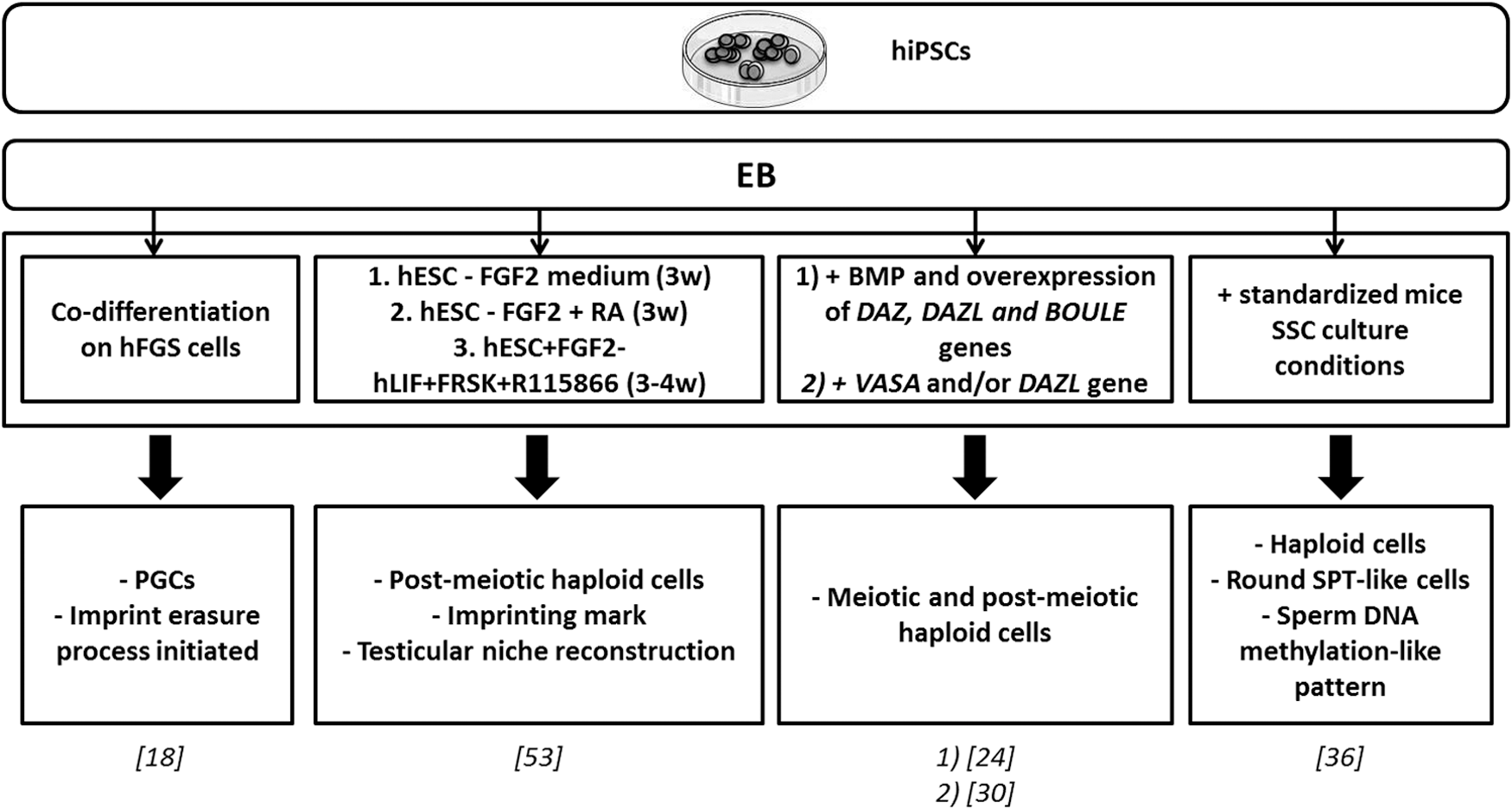
Main protocols for in vitro male germ line differentiation from hiPSCs. Germ cell differentiation from human iPSCs using forced expression of specific genes (DAZL, DAZ, BOULE, VASA) or introduced specific factors in the culture medium. BOULE, homolog of the human gene DAZ in Drosophila; DAZ, deleted in azoospermia; DAZL, deleted in azoospermia like; FGF2, fibroblast growth factor 2; FRSK, forskolin; hESC, human embryonic stem cell; hFGS, human fetal gonadal stromal; hiPSC, human-induced pluripotent stem cell; hLIF, human leukemia inhibitory factor; R115866, CYP26 inhibitor; VASA (DDX4), DEAD (Asp-Glu-Ala-Asp) box polypeptide 4; w, weeks.
Conversely, Eguizabal et al. and Easley et al. demonstrated that postmeiotic haploid cells could also be obtained from hiPSCs without the overexpression process of germ line-specific factors [36,53]. Eguizabal et al. developped a strong two-step differentiation protocol and obtained GCs differentiated into postmeiotic sperm-like cells showing ACR staining [53]. Furthermore, the expression level of six imprinted genes after 10 weeks of differentiation was evaluated that turned out to be comparable with sperm [53]. Efforts in this work to reproduce the physiological gonad microenvironment must be emphasised. Indeed, this 10 weeks protocol used hiPSCs of different origin and from both genetic sex for GC differentiation.
Eguizabal et al. demonstrated that without the overexpression of specific germ line-related regulator genes, postmeiotic haploid cells were successfully obtained from hiPSCs. Easley et al. also reported Piwi-like RNA-mediated gene silencing 1 (HIWI or PIWIL1) and HILI-positive spermatocyte-like cells, and round spermatid-like cells expressing ACR, Trophoblast protein-1 (TP1), PRM1, and showing a DNA methylation pattern similar to those of human sperm on at least two parent-of-origin genomic imprints [36].
Thus, since Park et al. [18], numerous studies have been performed on sperm and oocyte derivation from skin cells in mice and in humans. In this way, the attempts of iPSCs into male GCs appeared to be successful in vitro under EB formation and specific induction or by specific gene overexpression from iPSCs. In vitro haploidization was reported in several studies [24,36,52,53] at a stage close to spermatid [24,30] or even to sperm [53]. Combining in vitro and in vivo differentiations, late male GCs have been generated from iPSCs. In vivo transplantation led to the reconstruction of the testicular tissue [48 –51] that in some cases contained spermatogenetic cells [48 –51] or even spermatocytes [49] and spermatids [50]. As previously described, only one team tested the functionality of the haploid cells obtained from miPSCs, succeeding in giving birth to a fertile offspring [47].
Female GC differentiation from the iPSCs
Experiments in mice (Fig. 5)
Several groups reported in vitro formation of hiPSC-derived OLCs by using different techniques without in vivo steps. Niu et al. showed that RA and porcine follicular fluid were beneficial both for the derivation of male GCs as described above and of OLCs from miPSCs. OLCs expressed the Mvh, Dazl, Stra8, Scp3, and Zp3 after 7 days of differentiation [48]. Differentiation induction using coaggregation culture with cells producing BMP4, SCF, and epidermal growth factor led to an increased expression of premeiotic and meiotic markers (Supplementary Table S2). The formation of round-shaped cells Oct4-GFP−/Mvh-RFP+ resembling immature oocytes has also been reported (Supplementary Table S2) [26].
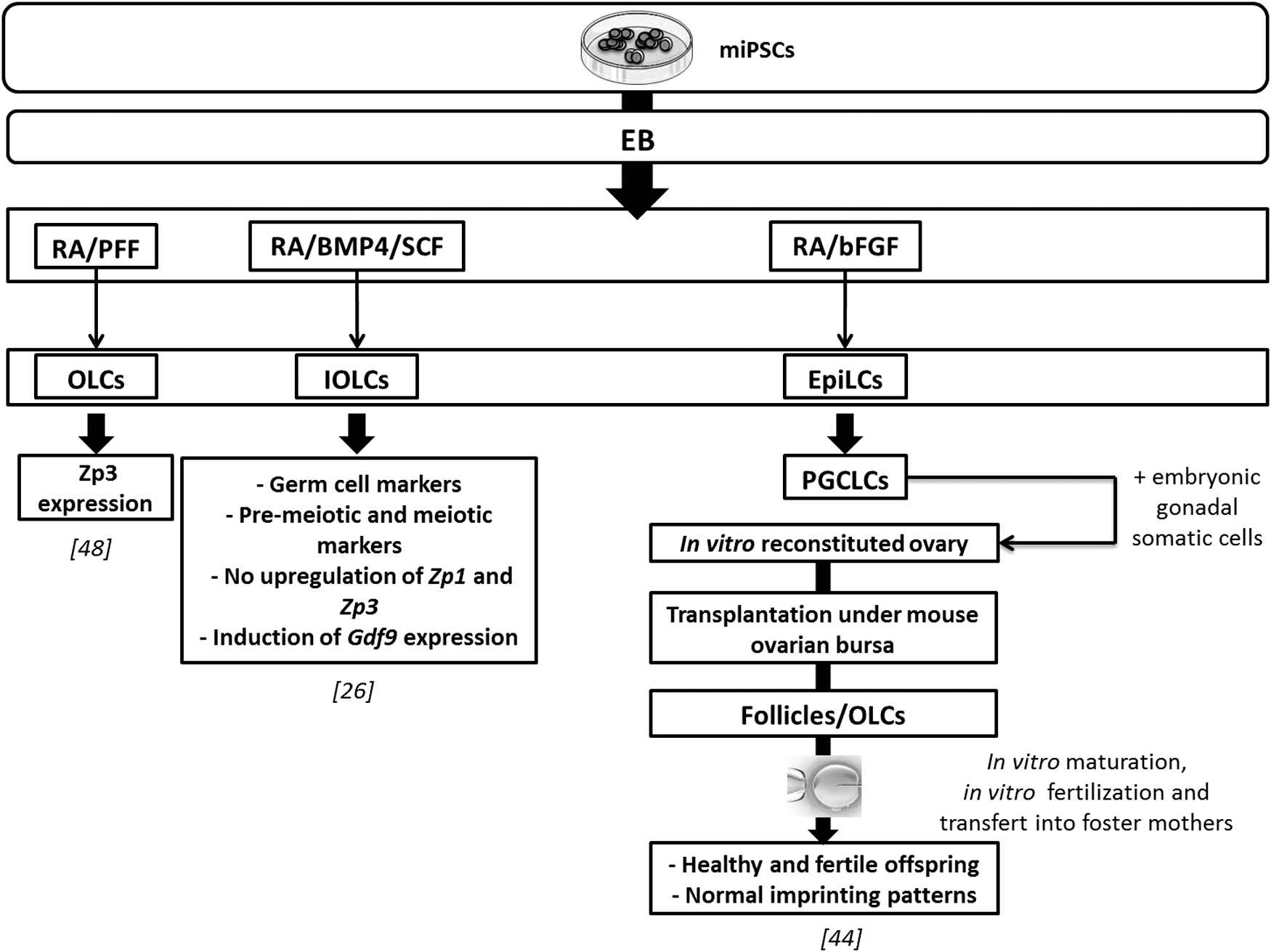
In vitro female germ line differentiation protocols from miPSCs. Female germ line differentiation from miPSCs using defined factors in culture medium. Oocyte-like cells were obtained. Their fertilization led to healthy and fertile offspring birth [44]. Gdf9, growth differentiation factor 9; IOLC, immature oocyte-like cell; OLC, oocyte-like cell; PFF, porcine follicular fluid; Zp1, 2, and 3: zona pellucida glycoprotein 1, 2, and 3.
Female miPSCs (bearing Pou5f1-EGFP transgene) were also shown to be efficient in inducing fully functional oocytes as described above from female mESCs [44]. PGCLCs first generated reconstituted ovaries and then formed OLCs and even contributed successfully to oogenesis when transplanted under ovarian bursa or kidney capsules of recipient mice. Following IVM and IVF from the generated PGCLCs, two-cell embryos were transferred into foster mothers. Three offsprings were obtained and grew into fertile adults [44].
Anchan et al. assumed that initiating iPSC generation from cells originating from the gonadal tissue may provide an advantage due to the iPSC epigenetic memory [56]. In this way, they functionally evaluated the epigenetic memory of novel iPSC lines derived from mouse and human ovarian granulosa cells. Thus, based on spontaneous differentiation, they obtained cells expressing ovarian markers (AMHR, FSHR, P450, family 19, subfamily A, polypeptide 1, estrogen receptor, and inhibin alpha) as well as markers of early gametogenesis (Mvh, Dazl, Gdf9, Boule, and Zp1) [56]. Furthermore, obtained ovarian and primitive GCs were present in higher incidence than in EBs of the other cell lines (from fibroblast or G4 ESCs) and were able to synthesize more estradiol. In summary, these data support the hypothesis for epigenetic-mediated mechanisms of homotypic differentiation of iPSCs [56].
Experiments in humans
In contrast to male germ line induction, not much has been achieved with female, especially from hiPSCs. However, some studies obtained promising results.
Indeed, a team used hiPSCs derived from women with primary ovarian insufficiency (POI) for further differentiation into GCs [55]. POI-iPSCs were transfected with the VASA-GFP construct for selection. Differentiation was induced through wingless-type MMTV integration site family, member 3a, and BMP4 addition. After 12 days of differentiation, the percentage of GFP-positive cells increased as did expression of GC markers (PRDM1, STELLA, DAZL, and VASA). GCs differentiated from POI-iPSCs showed a gene expression profile similar to the premeiotic stage [55]. Thus, iPSC technology could provide in vitro disease modeling helping to accelerate the development of novel therapies notably in fertility-related diseases [55].
In mice, OLCs were generated from miPSCs in four studies [26,44,48,56]. The specific protein ZP1 is a molecular marker of oocyte development competence, but was not systematically identified in the studies. Nevertheless, it has been demonstrated that miPSCs could be driven to develop into mature and functional oocytes [44]. Live births of an offspring using mice-induced GCs were reported [44].
Using a combined in vitro and in vivo induction approach, Hayashi et al. published a successful protocol for PGC derivation from mESCs and miPSCs [44,47]. Nevertheless, in vitro induction also reported great promises in research and clinical applications for human infertility treatment. Progresses were made in the male differentiation field, as demonstrated by the in vitro generation of postmeiotic haploid cells [36,52,53]. Oocyte-like structures were also reported in vitro [26,44,48,56] with even completion of meiosis [44]. To obtain in vitro functional gametes from hiPSCs, protocols need to be optimized in a hope of producing healthy offspring generated from artificial gametes.
Discussion
Much effort has been made in the generation of functional gametes from pluripotent stem cells, including ESCs and iPSCs. Results showed that it is easier to obtain male gametes with morphological and typical meiotic characteristics than mature oocytes. However, recent exciting achievements reported both derived murine sperm and oocytes functionality by ICSI and IVF giving birth to healthy offspring [44,47]. This demonstrated the relevance of animal models in this domain. To our knowledge, no article has described the generation of totally mature and functional gametes in humans. However, results suggest that it may be possible in future to investigate human gamete development in vitro to establish novel reproductive technologies.
The identification of key signaling molecules favoring PGC emergence allowed the development of in vitro protocols for PGC induction from mice and human pluripotent stem cells. Main efficient protocols are presented in Supplementary Table S2. Different experimental strategies were used for PGC differentiation: spontaneous differentiation [17,23,25], forced gene expression [16,24], or the use of defined factors or specific cell types [12,18,21,22,26].
Taken together, these strategies succeeded to various differentiation stages. Based on PGC-specific marker knowledge, authors were able to isolate a PGC population using a simple fluorescence-activated cell sorting assay. PGC-specific genes such as Oct4, c-kit, VASA, NANOS3, and CXCR4 were labeled. The number of PGCs generated in vivo was rather limited [57]. The alternative is to reproduce in vitro PGC differentiation steps starting from pluripotent stem cells [57]. During embryogenesis, two different transitory stages of PGCs can be identified: naive pluripotent stem cells sharing properties with the ICM of the blastocyst (ie, ESCs) and primed pluripotent stem cells similar to postimplantation epiblast cells of a more advanced pregastrulating stage embryo [57].
Pluripotency found in EpiLCs differs fundamentally from that of ESCs. The naive and primed pluripotent states can be easily discriminated as they exhibit a different miRNA expression profile. Therefore, efforts have been made to optimize culture conditions for induction or maintenance of either type of pluripotent stem cells in vitro. Naive ESCs can be maintained by LIF and adopt a primed EpiSC state following bFGF/Activin A induction. Finally, EpiSCs generated can lead to PGCLCs under an appropriate PGC differentiation medium [57].
Debates on the ethics of hESC research have considerably limited advances in the field, despite opening up a promising and pioneering area for biomedical applications and research (cellular therapies and transplantation) [58]. ESC research is the subject of fierce argument as it involves the use and destruction of human embryos [59]. Indeed, early embryos had to be destroyed to serve stem cell research; therefore, it raises serious ethical concerns not only among scientists but also politicians and the general population [58].
All this controversy relies on whether life begins at conception and whether we accord or not a moral status to the embryos [58,59]. In such a way, researchers of hESC topics are also considered complicit in the destruction of embryos that makes hESC research unethical [59]. As hESC research likely creates a demand for embryo destruction, iPSC research has been welcomed as an ethical alternative [59].
However, results in humans evolve quickly. Recently, hESCs and iPSCs were derived from azoospermic men carrying deletions in the AZF regions and were able to differentiate into PGCLCs in vitro and after transplant into mice seminiferous tubules [60]. This underlines the potential and progress of in vitro gametogenesis research in infertility treatment. Obviously, in vitro models contribute not only to open our knowledge of human gamete development and related disorders but also hold great promise for the development of alternative infertility treatments.
Nevertheless, other animal models in which gametogenesis physiology is closer to human (porcine, bovine, primate) could give faster advances. Indeed, PGCLCs have been successfully obtained from fetal porcine skin-derived iPSCs and were able to undergo imprint erasure and further differentiated into OLCs [61]. In vitro gametogenesis research from nonhuman primates' ESCs and iPSCs could also bring additional information on human GC derivation. There is no doubt there are similarities between the embryos and fetuses of nonhuman primates and humans [62]. Thus, Teramura et al. [62] differentiated nonhuman primate ESCs into PGCs, the fetal precursor stage of the germ lineage.
Gametogenesis success is not only connected to direct actions on concerned cells but also to the interaction of GCs with nearby somatic cells. Current studies focus particularly on gaining mature gametes. Targeted experimentation on somatic cells associated with gametes and its regulating factors remains underdeveloped. However, the establishment of the gonadotropic axis, its impact on somatic cells, and indirectly on gametes, is a decisive step in the accomplishment of gametogenesis. It is, therefore, critical to assess if gamete induction processes in vitro faithfully recapitulate normal GC development in vivo.
In one study in mice, an offspring obtained from ICSI experiments showed a limited life span, pointing out the existence of uncontrolled factors for a complete differentiation of the male lineage. If chromosomal sex of starting stem cell lineage has no influence on the male or female orientation, epigenetic factors largely unknown at present, but essential to the establishment of the imprint or to some differentiation stages, could be responsible for this deficiency.
Numerous genetic and epigenetic alterations concerning ESC and iPSC generation have been reported in the literature [63]. The hiPSCs and hESCs acquire several types of aberrations during in vitro culture. Indeed, it has been demonstrated that the acquisition of malignancy or tumorigenicity can occur with high similarities in adaptations, differences in inactivation status of the X chromosome according to lines, cultures and passages, mitochondrial mutations, genomic integrity and chromosomal abnormalities, culture adaptation phenomena, and frequent acquisition of aneuploidy during long-term culture [63]. Therefore, the preservation of genomic integrity during GC in vitro culture is an absolute necessity before considering any clinical application.
Conclusion
Recent advances in GC in vitro induction from pluripotent stem cells provide an unprecedented understanding of the mechanisms guiding PGC development and differentiation. Molecular marker and inductor knowledge required for GC induction allowed the development of in vitro gamete-like cells. Derivation of PGCs was for a long time considered a required first step toward realization of gametogenesis from pluripotent stem cells.
In the view of published works, the efficiency of the transition step to a primed epiblast-like state was demonstrated. It is now necessary to improve PGC formation in vitro. In addition, fertile spermatozoa and oocytes were generated from mESCs and miPSCs using a combined in vitro and in vivo induction system. Some obstacles remain, as in vivo maturation assays cannot be easily applied in humans. The main objective is to differentiate PGCs into mature and functional oocyte and sperm in vitro without any transplantation. It is, therefore, necessary to develop culture systems similar to the gonadal niche environment in humans. This also implies further understanding of the intrinsic regulators and interaction existing between GCs and their microenvironment.
However, in vitro reprogramming and differentiation processes are likely to raise deeper concerns, such as the appearance of abnormalities in epigenetic and imprinting status establishment in iPSCs. Regardless of this, GC research opens new therapeutic perspectives for reproductive medicine applications. Thus, infertility problems have hope of resolution in GC research aimed at generating patient-specific differentiated gametes from iPSCs.
Footnotes
Acknowledgment
We thank all the coauthors for their help with this work.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.