
Abstract
Lymphohematopoietic stem cells (L-HSCs) generated from self-somatic cell-derived induced pluripotent stem cells (iPSCs) are a potential source of cells for the treatment of hematological disorders. However, the generation of truly functional L-HSCs from iPSCs has yet to be achieved. Thus, whether iPSCs have the inherent potential to generate a normal differentiated phenotype and functional population of multiple lineages of terminally differentiated lymphocytes needs to be assessed. Here, we used tetraploid embryo complementation to provide a normal environment for the differentiation of hematopoietic cells from iPSCs and embryonic stem cells (ESCs). We then evaluated the characteristics, populations, and functions of lymphocytes derived from iPSCs, ESCs, and naïve isogenic C57BL/6 mice. The results showed that iPSC-derived lymphocytes (iPSLs) expressed normal levels of major histocompatibility complex-I (MHC-I) and exhibited a fully pluripotent capacity to differentiate into CD4+ T, CD8+ T, regulatory T, B, and natural killer cells. Following in vitro stimulation with either concanavalin A or an alloantigen, iPSLs exhibited the same capacities for proliferation and cytokine secretion as ESC-derived or isogenic lymphocytes. Furthermore, iPSC-derived bone marrow cells could differentiate into multiple lymphocyte lineages that reconstituted the lymphocyte population in syngeneic lethally irradiated recipient animals. Our results demonstrated that iPSCs have the inherent potential to differentiate into multiple lineages of functional lymphocytes without bias, and further support the practical application of iPSC-based treatments to hematological disorders.
Introduction
L
Although iPSCs can differentiate into hematopoietic precursors, there are many differences compared with somatic hematopoietic stem cells (HSCs), such as limited proliferation and migration capacities [16], early senescence [17], and poor multilineage engraftment characteristics [18]. Many genetic and epigenetic differences between iPSCs and ESCs have also been identified, which can promote tumorigenicity [19], hamper differentiation [20 –22], or lead to lineage bias by altering the signaling response in developmental pathways [14,23,24]. Additionally, epigenetic memory renders iPSCs prone to differentiate to their original somatic cell lineage [23,24]. Similarly, cord blood-derived iPSCs possess an enhanced hematopoietic differentiation potential compared with fibroblast-derived iPSCs [25,26]. Furthermore, iPSCs show a limited differentiation capacity for certain lineages such as hematopoietic cells, hepatocytes, and pancreatic cells [17,20,27,28]. Such limitations of iPSCs have restricted their capacity for hematopoietic regeneration, which is associated with the inability to downregulate microRNAs [29]. However, this limitation can be overcome by fusing iPSCs with HSCs [30]. Nevertheless, concerns remain that iPSCs possess inherent defects in multiple functional lymphocyte lineages, which may hinder their advantages in the clinical treatment of hematological disorders.
Thus, it is critical to investigate the ability of iPSCs to reconstitute a functional immune system within an optimal hematopoietic differentiation environment. The multilineage lymphocyte differential potential of iPSCs has been demonstrated in diploid blastocyst chimeras [31]. However, considering the coexistence of a non-iPSC population in the recipient, this approach does not confirm the inherent differentiation capacity of iPSCs in chimeras [32,33]. Tetraploid embryo (4N) complementation, as the gold standard for evaluation of iPSC pluripotency [34,35], can successfully avoid non-iPSC interference, because the fetus is formed completely by iPSCs [32,36,37]. Using this model, Tong et al. found that iPSC-derived mice (iPSm) had normal intelligence and memory, but a higher incidence of tumorigenesis compared with ESC-derived mice (ESm) [38]. However, a thorough evaluation of the postnatal development of the iPSm immune system is lacking. Here, we used 4N complementation to reproduce the entire process of hematopoiesis for iPSCs and ESCs. Then, the characteristics, constitution, and functions of lymphocytes from spleen and lymph node derivatives were compared among iPSm, ESm, and naïve isogenic C57BL/6 mice [wild-type (WT) control].
Materials and Methods
Experimental animals
All animals were purchased from Vital River Laboratory Animal Co. Ltd. (Beijing, China). Male C57BL/6 (H-2Kb) mice were used for iPSC and ESC generation in addition to WT control and syngeneic recipients. BALB/c mice were used as allogeneic stimulators. CD-1 mice were used as embryo donors and pseudopregnant female recipients. All animals were maintained and bred in specific pathogen-free facilities. Animal care and handling were conducted following the National Institutes of Health Principles of Laboratory Animal Care. The protocol was approved by the Animal Studies Committee of Xiamen University, China.
Generation of iPSCs from murine embryonic fibroblasts using an episomal plasmid
Murine embryonic fibroblasts (MEFs) were reprogrammed to iPSCs using an episomal plasmid transfection method as described previously [39,40]. Briefly, 6 μg of an episomal plasmid (Fig. 1A) harboring the coding sequences for OCT4, KLF4, SOX2, and NANOG was transfected into 1 × 106 C57BL/6 MEFs using the Neon® transfection system (Invitrogen, Carlsbad, CA), followed by culture in MEF medium for 24 h. ESC medium [41] with 5 μg/mL fibroblast growth factor-2 was used for subsequent culture. After ∼8 days, several single cell-formed colonies appeared with ESC-like phenotypes. These colonies were picked and expanded. The pluripotent properties of each clone were confirmed by staining with an alkaline phosphatase kit (Beyotime, Shanghai, China) and detecting pluripotency markers by immunofluorescence. For immunofluorescence staining, anti-NANOG (1:100 dilution; Abcam, Cambridge, United Kingdom), anti-OCT4 (1:50; Santa Cruz Biotechnology, Dallas, TX), and anti-stage-specific embryonic antigen 1 (SSEA1, 1:200; Millipore, Darmstadt, Germany) antibodies were used as primary antibodies. The cells were then incubated with AlexaFluor® 488- or 647-conjugated secondary antibodies diluted in 2% bovine serum albumin with 4.5% Triton X-100. Images were acquired using anLSM780 Meta confocal microscope (Zeiss, Oberkochen, Germany). Teratoma formation was detected in severe combined immunodeficiency mice following injection of 1 × 107 iPSCs. Four weeks later, teratomas were excised and analyzed by hematoxylin and eosin staining. G-banding karyotype analysis was carried out by Peking Union Medical College (Beijing, China). The qualified iPSC clones were cryopreserved for later use.
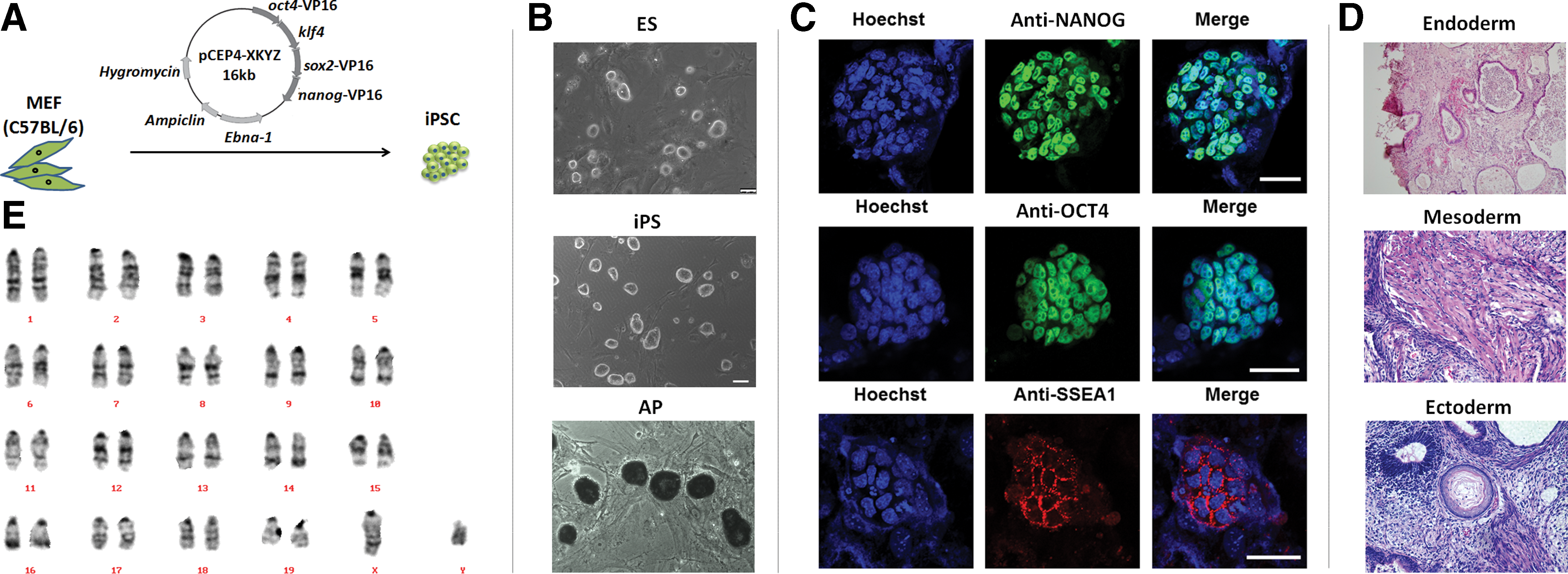
Generation and identification of pluripotent integration-free induced pluripotent stem cells (iPSCs).
Tetraploid embryo complementation
The generation of mice by 4N complementation was carried out as described previously [37]. Briefly, two-cell embryos were collected from the oviducts of CD-1 females, electrofused to produce one-cell tetraploid embryos, and then cultured in CZB medium (Fig. 2A). Each tetraploid blastocyst was then injected with 10–15 iPSCs or ESCs and transferred to CD-1 pseudopregnant female recipients. Fetuses derived from tetraploid blastocyst injection were dissected on the day of birth (embryonic day 19.5).
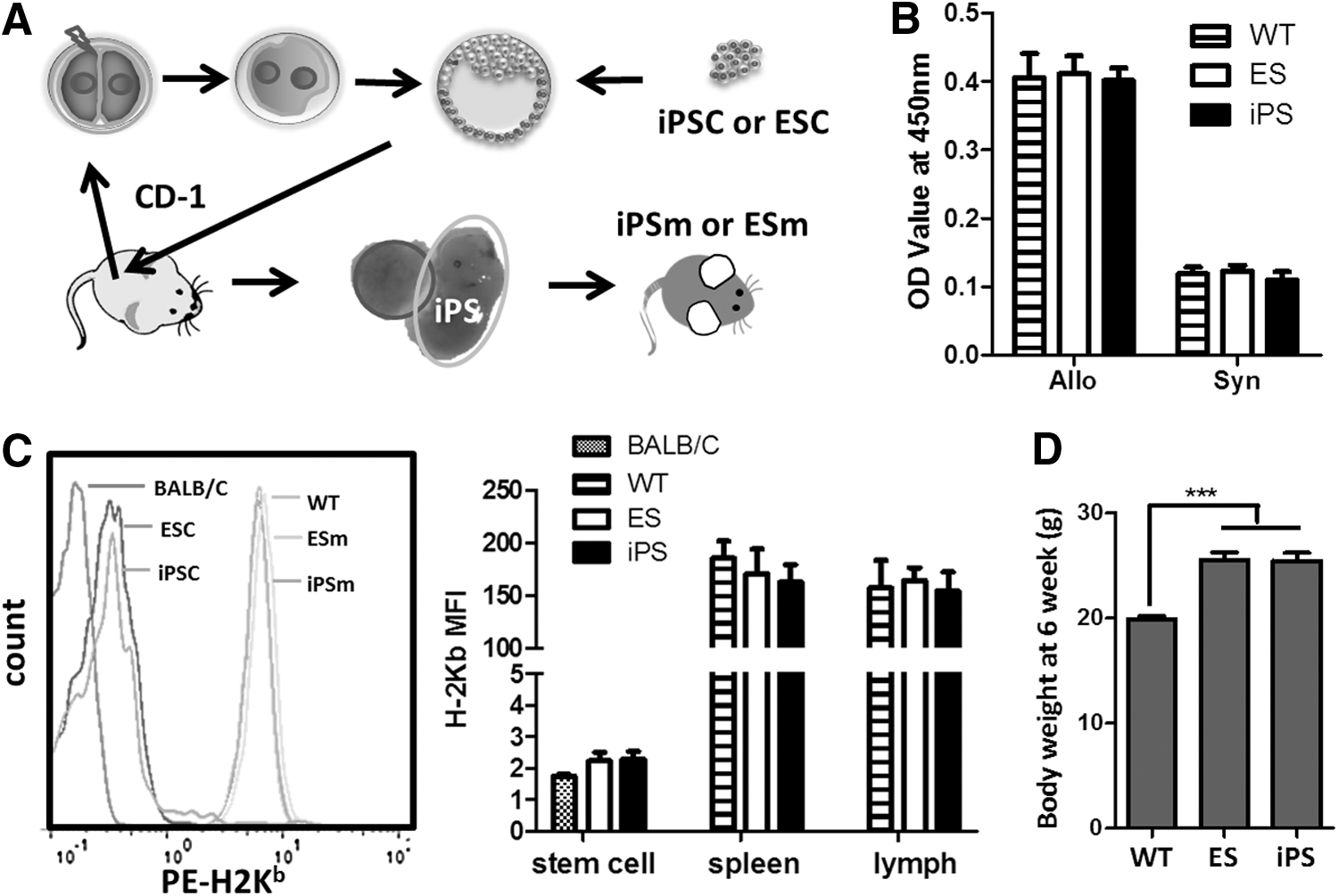
Generation and characterization of iPSm by 4N complementation.
Flow cytometry
Lymphocytes were obtained from the spleen and inguinal lymph nodes. After red blood cell lysis, 1 × 106 cells were resuspended in 100 μL FACS buffer and stained with antibodies for 30 min at 4°C. The antibodies used were PE-conjugated anti-H-2Kb (AF6-88.5), FITC-conjugated anti-mouse CD4 (RM4-5), PECY5-conjugated CD4 (GK1.5), PE-conjugated anti-mouse CD8a (53-6.7), PE-conjugated anti-mouse CD19 (6D5), PE-conjugated anti-mouse NK-1.1 (PK136), and relevant isotype controls (all from BioLegend, San Diego, CA). Regulatory T cells (Tregs) were labeled using the Mouse regulatory T-cell staining kit (eBioscience, San Diego, CA) following the manufacturer's protocol. Stained cells were detected on a flow cytometer (Partec Co., Munster, Germany). Data were analyzed using FlowJo software (Tree Star, Inc., Ashland, OR).
Lymphocyte proliferation assay
For the lymphocyte transformation test, splenocytes were first stimulated with 5 μg/mL concanavalin A (ConA). For the mixed lymphocyte reaction (MLR) assay, 5 × 105 responder cells were stimulated with 5 × 104 splenocytes pretreated with mitomycin C (40 mg/mL; Amresco, Solon, OH). Cells were cultured in 200 μL RPMI 1640 medium supplemented with 10% (vol/vol) fetal bovine serum, and 1% penicillin and streptomycin, and incubated in 96-well plates at 37°C in a humidified atmosphere with 5% CO2 for 48 h. Lymphocyte proliferation was detected using either the Cell Trace carboxyfluorescein succinimidylester (CFSE) Cell Proliferation Kit (Molecular Probes, Invitrogen, Eugene, OR) or BrdU Cell Proliferation Assay Kit (Roche Diagnostics, Indianapolis, IN) according to the manufacturers' instructions. For CFSE staining, cells were stained with 5 μM CFSE reagent for 7 min before stimulation. Cells were also stained with PECy5-conjugated anti-mouse CD4 and PE-conjugated anti-mouse CD8a antibodies followed by flow cytometry (FCM) analysis. For the BrdU assay, a microplate reader(Model 680; Bio-Rad Laboratories, Hercules, CA) was used to obtain optical density values at 450 nm (reference wavelength:570 nm). All assays were performed in triplicate.
Cytokine secretion assay
Culture supernatants were collected after 48 h of stimulation with or without ConA. The presence of interleukin (IL)-2, IL-4, IL-6, IL-10, IL-17, tumor necrosis factor (TNF), and interferon-γ (IFN-γ) was detected using a Cytometric Bead Array (CBA; BD Biosciences, San Jose, CA) and FACS Aria (BD Biosciences) according to the manufacturer's instructions. Data were analyzed by FCAP Array™ Software v1.0.1 (Soft Flow Hungary Ltd., BD Biosciences). Each sample was analyzed in triplicate.
Bone marrow transplantation
Single cell suspensions of bone marrow cells (BMCs) were prepared from the femurs and tibias of 6-week-old iPSm, ESm, or WT control mice. Following lethal X-ray irradiation (8.5 Gy), each 8-week-old C57BL/6 mouse recipient received a single intravenous injection of 1 × 107 BMCs. Mice in the control group were administered phosphate-buffered saline. Four weeks later, peripheral blood cells from recipient mice were analyzed using an automated hematology analyzer (Sysmex KX-21N, Japan) and FCM.
Statistical analysis
Data obtained from ESm, iPSm, and WT C57BL/6 mice were analyzed by one-way analysis of variance with the Bonferroni correction. To show variability between individual cell lines, representative data from each cell line are provided in the Supplement. Results are expressed as the mean ± standard deviation. A value of P < 0.05 was considered to indicate a statistically significant difference. Samples were obtained from 10 mice in each group (three to five mice derived from each cell line were chosen). All statistical analyses were conducted using GraphPadPrism software (GraphPad Software, Inc., San Diego, CA).
Results
Generation of iPSC- and ESm by 4N complementation
Three genome integration-free iPSC lines (iPS-1, -2, and -3) and two ESC lines (ES-1 and-2) generated from C57BL/6 donors were chosen. All iPSC lines were assessed for pluripotent properties by comparison with ESCs based on morphology (Fig. 1B), alkaline phosphatase staining (Fig. 1B), pluripotency marker staining (Fig. 1C), and teratoma formation (Fig. 1D). All iPSC lines showed the same pluripotent characteristics as ESC lines (Table 1). G-banding karyotype analysis showed that reprogramming did not cause chromosomal abnormities in iPSCs (Fig. 1E). Stem cells were injected into more than 100 electrofused embryos for each cell line, and about 40%–52% of embryos were implanted. Few embryos survived to full-term pups. Only 2.8%–8.5% iPSC-embryos and 5.9% (ES-2) or 12% (ES-1) ESC-embryos survived to adulthood (Table 1). No tumors or other lethal diseases were identified in iPSC- or ESC-derived adult mice. The mean adult (6 weeks old) body weights of ESm and iPSm were both higher than that of normal syngeneic mice (Fig. 2D and Supplementary Fig. S1A; Supplementary Data are available online at
AP+ clusters%, alkaline phosphatase positive clusters/total clusters, n > 60; NANOG, OCT4 or SSEA1 positive cell%, positive stained cells/total number of cells within the cluster; survival ratio, survival number/injected embryo number.
Expression of major histocompatibility complex-I proteins in ESCs, iPSCs, and their lymphocyte derivatives
Major histocompatibility complex-I (MHC-I) is important for self-recognition, cell differentiation, and transplantation rejection [42]. We measured the expression of MHC-I proteins on ESCs, iPSCs, and their derived lymphocytes by analyzing H-2Kb expression using FCM. The mean fluorescence intensity of each ESC and iPSC line was as low as that of BALB/c splenocytes (Supplementary Fig. S1B). Statistical data showed a decrease in the MHC-I expression level on iPSC lines after reprogramming, which was similar to ESCs and BALB/c negative controls (Fig. 2C). Following differentiation, both iPSm and ESm lymphocytes from the spleen and lymph nodes had the same levels of MHC-I expression as those from syngeneic WT mice (Fig. 2C). As the most important allo-immunogenicity marker, the MHC-I expression level on iPSC-derived lymphocytes (iPSLs) was further confirmed by MLR. No differences in the stimulation ability were found between cell lines (Supplementary Fig. S1C). All iPSLs could stimulate the alloreactive responses of BALB/C (Allo) lymphocytes as effectively as ESm- and WT mouse-derived lymphocytes. However, no such response was elicited from C57BL/6 (Syn) lymphocytes by iPSLs, ESm-, or C57BL/6 (WT) mouse-derived lymphocytes (Fig. 2B).
Composition of lymphocyte populations in the lymphoid organs of iPSm and ESm
We next investigated whether iPSLs formed T, B, or natural killer (NK) cell populations within the normal cell population and their phenotypes in peripheral immune organs. Lymphocytes were obtained from the spleens and lymph nodes of 6-week-old adult iPSm, ESm, and C57BL/6 mice, and then assessed by FCM. For iPSm splenic lymphocyte populations, the proportions of B, NK, CD4+ T, CD8+ T cells, and Tregs were comparable to those in C57BL/6 lymphocyte populations (Fig. 3A–C and Supplementary Fig. S2A–C). However, splenic lymphocytes in iPSm had a higher proportion of CD4+ T cells compared with those in ESm (Fig. 3C). Although CD4+ T cell numbers derived from each iPSC line showed an increasing trend, only CD4+ T cells from iPS-3 showed a significant increase compared with those from ES-1 and ES-2 lines (Supplementary Fig. S2C). The variation among the iPSC lines was negligible. Similar increasing trends were identified in comparisons of the proportions of CD4+/CD8+ (Fig. 3C and Supplementary Fig. S2C) and Foxp3+/CD4+ T (Fig. 3D) cells. Although only the proportion of CD4+/CD8+ T cells from iPS-3 was higher than that from ES-1, it was not higher than that of other cell lines. Only the proportion of splenic Foxp3+/CD4+ T cells from iPS-2 was higher than that from ES-1 and ES-2 lines (Supplementary Fig. S2C). In lymph nodes, each type of lymphocyte maintained the same proportions in each iPSC line-derived mouse (Supplementary Fig. S2D), and the aggregate proportions in iPSm did not significantly differ from that in WT mice and ESm (Fig. 3C, D). No clear CD4+/CD8+ cell cluster (less than 0.5% total lymphocytes) was identified in any of these groups.
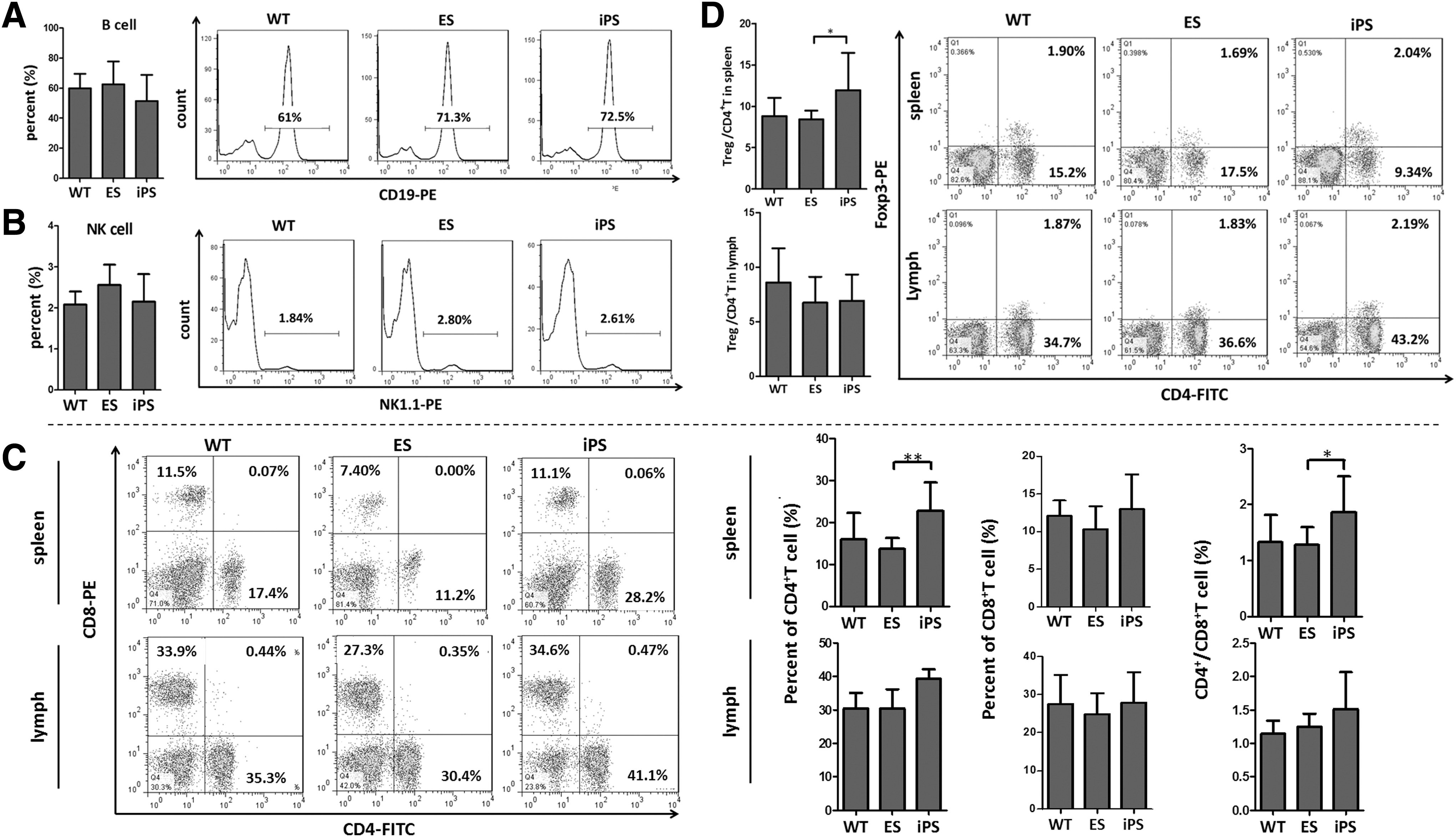
Composition of iPSC-derived lymphocyte populations. Lymphocytes from the spleen and lymph nodes were harvested and analyzed by FCM. Populations of B cells
IPSLs exhibit normal proliferative and secretory functions
We next assessed the proliferative capacities of lymphocytes derived from ESm, iPSm, and C57BL/6 mice. Splenocytes were separated and stimulated with ConA for 48 h, followed by CFSE and BrdU staining. Results showed similar proliferative capacities among lymphocytes derived from each iPSC line (Supplementary Fig. S3A, B). Both iPSC- and ESC-derived lymphocytes had a proliferative capacity similar to that ofC57BL/6 lymphocytes (Fig. 4A, B). Cytokine levels in culture supernatants were then assessed by CBA. Similar levels of IL-2, IL-4, IL-6, IL-10, IL-17, TNF, and IFN-γ were found in iPSC, ESC, and C57BL/6 lymphocyte culture supernatants (Fig. 4D) and between each iPSC and ESC line (Supplementary Fig. S3C). An MLR was then performed to detect the ability of iPSLs to mount an adaptive immune response. Similar to the lymphocytes from ESm and WT mice, iPSLs were found to proliferate under allogeneic stimulation, but not under syngeneic stimulation (Fig. 4C and Supplementary Fig. S3D).
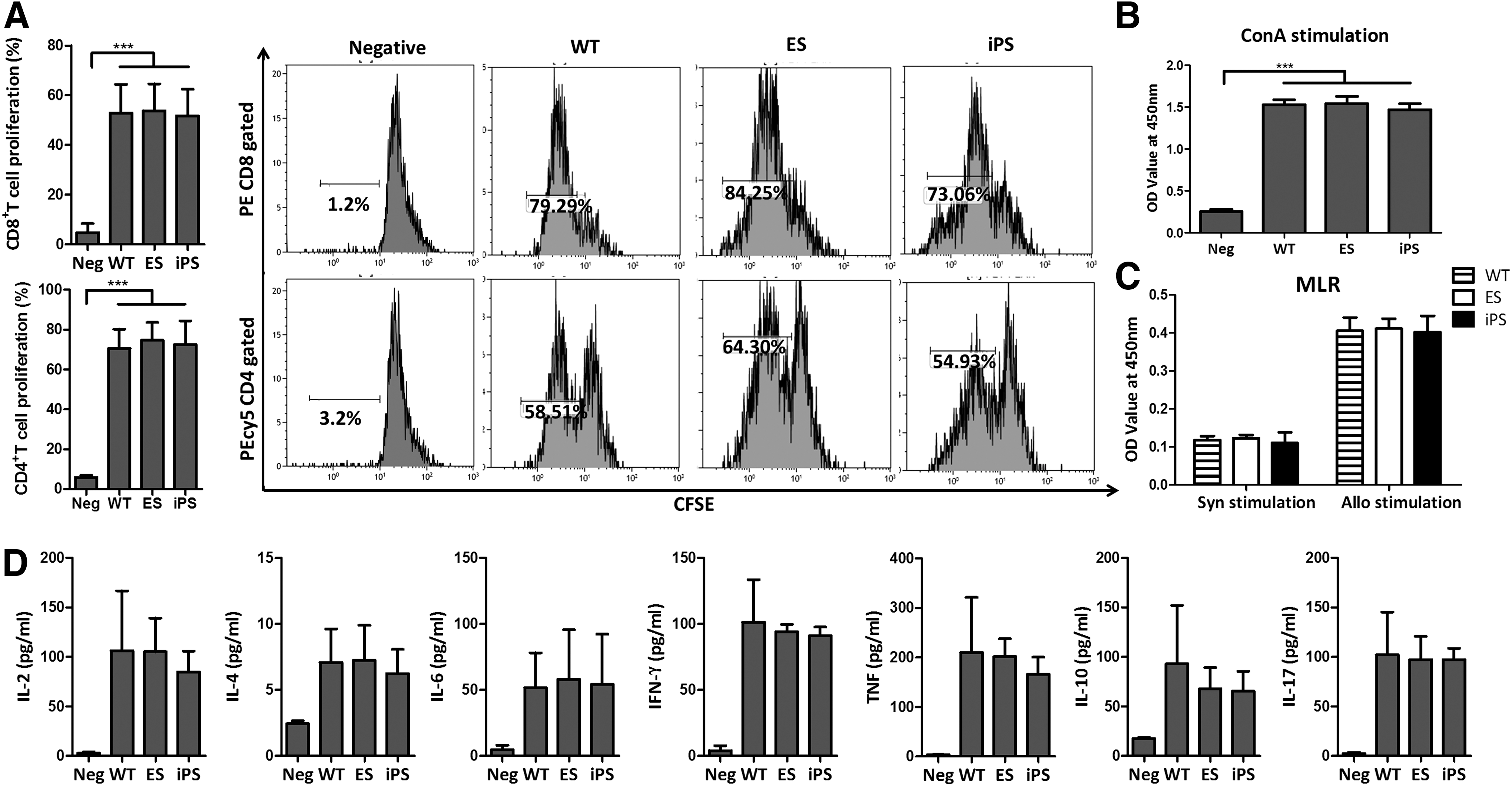
Lymphocyte proliferative and secretory functions. Lymphocytes were isolated from C57BL/6 (WT), ESm, and iPSm spleens.
IPSC-derived BMCs effectively reconstitute the lymphocyte population following myeloablation in syngeneic mice
We next examined the hematolymphopoietic reconstitution potential of iPSC-derived HSCs. BMCs derived from iPSm (iPS-BMCs), ESm (ES-BMCs), or WT mice were injected into syngeneic mice that had been exposed to a lethal dose (8.5 Gy) of X-ray radiation. iPS-BMCs prolonged recipient survival to beyond 1 month, which were as effective as ES-BMCs and WT-BMCs (Fig. 5A). No major phenotypic or behavioral abnormalities were observed in any of these mice after more than 1 month following transplantation. Furthermore, BMCs of all origins negated lymphocyte storage exhaustion in the spleen (Fig. 5B) and peripheral blood (Fig. 5C). FCM analysis demonstrated that iPS-BMCs could effectively regenerate and maintain lymphocyte populations in syngeneic recipients (Fig. 5D). At 1 month after BMC transplantation, similar proportions of CD4+ T, CD8+ T, B, and NK cells were detected in recipient peripheral blood of each group (Fig. 5E).

Reconstitution of lymphocytes after bone marrow transplantation. Bone marrow was harvested from C57BL/6 (WT), ESm, and iPSm. Then, 1 × 107 cells were injected into lethally irradiated C57BL/6 recipients (n = 10 per group).
Discussion
The differentiation capacity of iPSCs is critical for the safety and function of their derivatives [22]. MHC expression on ESCs and iPSCs is usually enhanced by strict epigenetic regulation following embryonic body formation [43]. Low expression levels of MHC-I block immune recognition and promote tumor formation [44]. Such low levels of MHC-I expression also reflect the undifferentiated nature of ESCs and their derivatives [42]. However, human iPSC-derived CD34+ hematopoietic progenitor cells have been found to poorly express MHC-I, which leads to T cell anergy [45]. Our results showed that the level of MHC-I expression in iPSCs was as low as that in ESCs. After differentiation, iPSLs expressed the same level of MHC-I as syngeneic WT lymphocytes that can stimulate allogeneic but not syngeneic lymphocyte responses. These findings suggest that iPSLs are fully differentiated in vivo without any decreases in MHC-I expression related to tumor formation.
Previous studies have shown that iPSCs can differentiate into multiple lymphocyte lineages including pre-B cells [7], functional T cells [46], and NK cells [9]. However, variations have been reported in the efficiency of hematopoietic differentiation in iPSCs and ESCs. Human iPSC cultures often represent a progenitor stage and fail to complete differentiation to HSCs in vivo [11]. Furthermore, fibroblast-derived iPSCs exhibit a lower hematopoietic differentiation efficiency than cord blood-derived iPSCs, which is due to lineage memory issues [25,26]. Here, we found that MEF- derived iPSCs had the same differentiation potential as ESCs for multiple lymphocyte lineages without any bias compared with WT controls. The only identified statistically significant difference in lymphocyte populations was higher proportions of CD4+ T cells and the corresponding higher Treg and CD4+/CD8+ cell proportions in splenocytes from iPSm, which were comparable with those from ESm. By analyzing the variations between each iPSC line, CD4+ and CD4+/CD8+ T cell proportions showed a statistically significant increase only in mice derived from iPS-3 compared with those derived from ES-1 and ES-2, while only mice derived from iPS-2 showed a statistically significant increase in Tregs. This difference was limited to the central immune organs, and no statistically significant changes were found in peripheral lymph nodes. This finding indicates that the increase in these lymphocytes was not caused by inflammation. It is possible that reprogramming and long-term culture may promote heterogeneity and stimulate cell proliferation [47,48]. However, no difference was found by routine identification of pluripotency (Table 1). Additionally, the range of variation in the iPSm lymphocyte composition is still within the confidence interval of WT mice. It appears that the difference between respective cell lines reflects the individual epigenetic difference between each syngeneic mouse. Furthermore, this iPSC heterogeneity did not influence cell functions, expansion, or cytokine secretion characteristics. In contrast to a previous report, which claimed that iPSC-derived hemangioblastic cells exhibit limited expansion and early senescence [17], we found that iPSCs are capable of generating lymphocyte populations with normal proliferative capacities following stimulation with mitogens and alloantigens. Considering the difficulties involved in generating multiple lineages of functional lymphocytes, there are only a limited number of reports on the secretory functions of iPSLs. Vizcardo et al. found that CD8+ T cells derived from iPSCs secrete IFN-γ after stimulation [46]. Here, we demonstrated that iPSLs secrete all Th1, Th2, and Th17 cytokines after stimulation. Additionally, consistent with a previous study on BMC transplantation in chimeric mice [31], we demonstrated that the inherent ability of iPS-BMCs to reconstitute lymphocyte populations in syngeneic recipients subjected to myeloablation. Taken together, these findings indicate that iPSCs have the ability to undergo unbiased differentiation into multiple lineages of lymphocytes with normal proliferation, secretion, and regenerative functions.
A possible explanation for the lack of biased differentiation and the capacity for heterogeneous functions identified in our study, which were compared with those in other studies, might be the iPSC generation protocol employing the nonintegrated reprogramming system and the in vivo differentiation environment by 4N complementation. Following the use of an episomal plasmid transfection reprogramming system, we did not detect any tumorigenicity as reported by another study [38]. In our previous study based on the same iPSC lines, we assessed insertion of mutations, Yamanaka factor overexpression, copy number variations, and transcriptional profile variations, and found no genetic or epigenetic variations leading to tumorigenicity [40]. The 4N complementation method might be another reason for the difference. This method enables re-creation of complete hematogenesis microenvironments, including those present during gastrulation and in the yolk sac, fetal liver, and bone marrow. The migration enrichment step and provision of a suitable microenvironment guaranteed accurate and efficient hemopoietic differentiation. However, none of these microenvironments are fully understood, and they are too complex to re-create in vitro or in an adult in vivo [14]. Furthermore, at present, genetic modifications are necessary to generate fully functional HSCs from mouse or human iPSCs in vitro [49]. However, such modifications may promote abnormal hematopoiesis [50,51]. Therefore, 4N complementation facilitates the inherent characteristics of iPSCs by excluding the interference in a raw differentiation protocol. Indeed, according to phenotypic analysis of a large cohort of ESm, the 4N complementation procedure might cause body weight increase by common experimental procedures including in vitro culture of embryos, specific pre- and postnatal nursing conditions, and maternal uterine heterosis but not obesity [52,53]. Here, we also found that the body weight of ESm and iPSm were higher than that of isogeneic mice. Although the reason is still unclear, we confirmed that 4N complementation and the body weight increase did not influence the composition of the lymphocyte population based on no significant statistical differences between ESm and isogeneic mouse lymphocyte populations. Furthermore, during 4N complementation, both iPSCs and ESCs might be subjected to strict selection, because only 2.8%–8.5% of iPSC-derived embryos and 5.9% (ES-2) or 12% (ES-1) of ESC-derived embryos survived to adulthood. Limited by the number of cell lines, it is difficult conclude whether any developmental variations existed between ESCs and iPSCs. A large deviation existed between iPSCs and ESCs and also between each cell line, which might have been due to various human errors such as embryo quality, the gravidas' status, blastosphere injection, and other procedures. Although it is still unclear why more than 90% of embryos were eliminated, 4N complementation narrowed the system deviation between iPSm and ESm compared with WT mice and made them comparable by strict selection of viable embryos. Only cells that exhibit pluripotency and form a structurally correct embryo in a suitable environment can generate a viable mouse. All these benefits might ensure normal differentiation of iPSCs, and clearly demonstrated the inherent differentiation ability of iPSCs under optimal conditions.
In this study, we demonstrated that iPSCs possess appropriate lymphocyte differentiation potency. Their lymphocytes exhibit normal phenotypes and possess normal proliferative, secretory, and regenerative capacities. These findings further support the possibility of the ultimate use of iPSCs for regeneration and transplantation by further optimization of induction and differentiation protocols to narrow the gap between the results obtainable in vivo and what can be achieved in a wholly in vitro context.
Footnotes
Acknowledgments
This work was supported by grants from the Major State Scientific Research Program of China (no. 2012CBA01303) and the National Natural Science Foundation of China (nos. 31271038, 81302546, and 81441047).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.