
Abstract
Mesenchymal stromal cells (MSCs) are applied as novel therapeutics for their regenerative and immune-suppressive capacities. Clinical applications, however, require extensive expansion of MSCs. Fetal bone marrow-derived MSCs (FBMSCs) proliferate faster than adult bone marrow-derived MSC (ABMSCs). To optimize expansion and function of MSC in general, we explored the differences between ABMSC and FBMSC. Gene expression profiling implicated differential expression of genes encoding proteins in the Wnt signaling pathway, including excreted inhibitors of Wnt signaling, particularly by ABMSC. Both MSC types had a similar basal level of canonical Wnt signaling. Abrogation of autocrine Wnt production by inhibitor of Wnt production-2 (IWP2) reduced canonical Wnt signaling and cell proliferation of FBMSCs, but hardly affected ABMSC. Addition of exogenous Wnt3a, however, induced expression of the target genes lymphocyte enhancer-binding factor (LEF) and T-cell factor (TCF) faster and at lower Wnt3a levels in ABMSC compared to FBMSC. Medium replacement experiments indicated that ABMSC produce an inhibitor of Wnt signaling that is effective on ABMSC itself but not on FBMSC, whereas FBMSC excrete (Wnt) factors that stimulate proliferation of ABMSC. In contrast, FBMSC were not able to support hematopoiesis, whereas ABMSC displayed hematopoietic support sensitive to IWP2, the inhibitor of Wnt factor excretion. In conclusion, ABMSC and FBMSC differ in their Wnt signature. While FBMSC produced factors, including Wnt signals, that enhanced MSC proliferation, ABMSC produced Wnt factors in a setting that enhanced hematopoietic support. Thus, further unraveling the molecular basis of this phenomenon may lead to improvement of clinical expansion protocols of ABMSCs.
Introduction
M
MSC represent a relatively rare cell population in the adult bone marrow, and the frequency of MSC declines with increasing age from 1 per 104 nucleated bone marrow cells in a newborn to about 1 per 106 nucleated marrow cells in the elderly [6]. The prevalence of MSC in fetal tissues is significantly higher and estimated at 1 per 400 in the second trimester fetal bone marrow (4, reviewed in O'Donoghue and Fisk [7]). Both culture-expanded adult and fetal MSC have some fundamental properties that are used to define them as bona fide MSC [8]. They can be derived by plastic adherence, express CD73, CD90, and CD105, and lack expression of hematopoietic markers. In addition, MSC have trilineage differentiation potential [4] and are able to suppress T-cell proliferation [9,10]. Although these MSC properties are shared by adult and fetal MSC, there are pivotal aspects that distinguish these two cell sources.
ABMSC and fetal bone marrow-derived MSC (FBMSC) are isolated at different developmental stages, which are reflected in different properties of the MSC. It is suggested that FBMSC are important for bone formation, to generate the bone marrow niche, and to prepare it for definitive hematopoiesis [11,12]. ABMSC are involved in maintenance and support of hematopoietic stem and progenitor cells (HSPC) in the fully matured bone marrow.
The differential expression of genes associated with Wnt signaling prompted us to focus on this pathway because Wnt signaling is involved in regulation of tissue development and tumorigenesis [13,14], bone formation [15], MSC differentiation [16 –18] and proliferation [16,19,20], and hematopoietic support [21 –23]. Wnt factors comprise a large family of secreted, lipid-modified glycoproteins that are expressed in a variety of tissues [13]. Wnt proteins bind a receptor complex consisting of a Frizzled receptor and the LDL receptor-related proteins LRP5 or LRP6. Activation of the canonical Wnt pathway stabilizes beta-catenin that accumulates in the nucleus where it activates target gene expression, among others genes encoding the transcription factors, lymphocyte enhancer-binding factor (LEF) and T-cell factor (TCF) [24]. Noncanonical pathways induce the release of Ca2+, or the phosphorylation of Jun N-terminal kinase (JNK) [24 –26]. Wnts are promiscuous and bind different classes of receptors. The biological effect of signaling depends on the receptor and its context, and not on the specific Wnt [27,28]. New players of the Wnt pathway, such as the R-spondin receptors LGR4 and −5, and Norrin added another layer of complexity on Wnt signaling [26,29,30].
Wnts are long known to control maintenance of HSPC, although seemingly contradictory results were published with respect to hematopoietic stem cell (HSC) renewal, and progenitor proliferation and differentiation. Wnt levels are crucial and tightly regulated by the HSPC environment [31,32]. Cultured ABMSC express a variety of Wnt proteins and receptors [33], but how Wnt production and signaling are controlled in FBMSC, when hematopoiesis still has to develop, is unknown.
In the current study, we demonstrate that several Wnt-related genes were differentially expressed between ABMSCs and FBMSCs, among which regulators of Wnt signaling resulted in different sensitivity and reactivity of the cells. Inhibition of endogenous Wnt production affected the proliferation of FBMSC, but not their hematopoietic-supportive capabilities. The opposite was observed for ABMSCs: they retained proliferating capacity, but lost the ability to support HSPC when endogenous Wnt production was attenuated. Unraveling the molecular basis of this phenomenon is of high clinical importance and may improve the production of adult MSCs for therapeutic use.
Materials and Methods
Cell culture
All media were supplemented with 10% fetal calf serum (FCS; Bodinco), 50 U/mL penicillin, and 50 μg/mL streptomycin (PS; Gibco), unless indicated otherwise.
Fetal bones were obtained from legally terminated second trimester pregnancies (15–20 weeks) after informed consent and with approval of the medical ethical review board of the Academic Medical Center (AMC) (MEC: 03/038). To obtain FBMSC, fetal bones were flushed with Iscove's modified Dulbecco's medium/FCS/PS (Lonza). Erythrocytes were lysed using NH4Cl for 10′ on ice. Cells were rinsed in PBS. Cells/well (1.6 × 106) were seeded in six-well dishes in MSC culture medium [M199 (Gibco), FCS-PS, 20 μg/mL endothelial cell growth factor (Roche Diagnostics), and 8 IU/mL heparin (Leo Pharma)].
Nonadherent cells were removed after 48 h, and the remaining cells were considered FBMSC.
ABMSC were isolated from bone marrow aspirates from the sternum of patients (age 40–70 years) undergoing cardiac surgery after informed consent and approval of the medical ethical review board of the AMC (MEC:04/042 No. 04.17.370).
Upon reaching 80% confluency after initial plating, ABMSC and FBMSC were replated and cultured in T80 tissue culture flasks at an initial density of 2,500 cells/cm2. For all experiments, 80% confluent passage 3–6 MSC were used. CD34+ cord blood cells were isolated as described before [34]. L-cells, L-Wnt3a, and L-Wnt-5a were a kind gift from Dr. D. ten Berge (Department of Cell Biology, Erasmus Stem Cell Institute) and were cultured in Dulbecco's modified Eagle's medium-FCS-PS and 0.4–0.6 mg/mL G418 (PAA Laboratories, for L-Wnt3a and Wnt-5a only). Cells were counted using a Casy Cell Counter (Casy® Technology, Innovartis). Pictures were taken with Leica 351 microscope (Leica Microsystems, Wetzair GmbH) with 10 × magnification objective.
RNA extraction and microarray analysis
RNA was extracted from third passage adult or fetal MSC (three adult donors, three fetal donors) using a RNeasy mini kit (Qiagen) according to the manufacturer's instructions. RNA integrity was analyzed on an Agilent 2100 bioanalyzer (Agilent Technologies, Inc.) using the Agilent RNA 6000 Non-Assay protocol. Three hundred nanograms' RNA was hybridized to an Affymetrix Human Exon 1.0 ST array (Affymetrix, Inc.). Hybridized assays were scanned with a GeneChip scanner 3000 7G (Affymetrix). Affymetrix CEL files were imported into Partek® (Partek Genomic Suite software, version 6.4 Copyright © 2008; Partek, Inc.), where only core probe sets were extracted and normalized using the robust multi-array average algorithm with GC background correction. Core transcript summaries were calculated using the mean intensities of the corresponding probe sets, representing the quantitative expression levels of all genes. The correspondence of the three replicate adult and fetal samples was confirmed using principle component analysis and Pearson correlation analysis. We performed an analysis of variance (ANOVA) on the log2 probe intensities, representing the gene expression intensities, of 15 samples and did a post hoc analysis to compare adult and fetal MSC samples. Data were corrected for multiple testing. Genes were considered significantly differentially expressed when P ≤ 0.05. Data are deposited in Gene Expression Omnibus under the accession No. GSE68374.
cDNA synthesis and real time quantitative PCR
A maximum of 1 μg total RNA was used for cDNA synthesis using random hexamers (25 μM; Gibco-BRL Life Technologies). Real time quantitative PCR (RQ-PCRs) were performed on a StepOne Plus (Applied Biosystems) using SYBR green dye for detection (SYBR green mastermix, Applied Biosystems [9]). Primer sequences are listed in Supplementary Table S1 (Supplementary Materials and Methods are available online at
Factor stimulation
Subconfluent MSC cultures were stimulated with 50% L-cell supernatants, or with recombinant human Wnt3a (R&D Systems) as indicated. The final concentrations were 50% L-cell conditioned medium (CM), or 200 ng/mL Wnt3a in M199 medium, and 0.2–25 μM inhibitor of Wnt production-2 (IWP2; Miltenyi Biotech) [35] for 16 h in fresh medium unless mentioned otherwise. Thirty thousand MSC were plated 2 days before Wnt stimulation in 12-well plates. For medium change experiments, both ABMSC and FBMSC were seeded at 2,500/cm2 2 days before the harvest of CM. MSC were seeded at 2,500 cells/cm2 for short-term proliferation experiments, and cells were counted by flow cytometry using Flow-Count Fluorospheres (Beckman Coulter).
Production of lentiviral particles
7TGC (Addgene plasmid No. 24304) was a gift from Roel Nusse [36]. DNA was harvested by maxiprep (Nucleobond; Clontech). T cells (293) were transfected with psPax2, pMD2.G, and 7TGC by the calcium phosphate method [9]. Viral particles were harvested after 1 and 2 days. MSC were infected with concentrated virus at a multiplicity of infection of 100. Transduction efficiency was measured by flow cytometry.
Cell lysis and western blot
MSC were lysed in RIPA buffer [containing protease and phosphatase inhibitors (Roche)] and DNase I (Invitrogen) for 10′ at 4°C. Lysates were cleared by centrifugation at 14,000 rpm at 4°C for 10′. Protein samples were separated by electrophoresis using a Criterion Pre-cast Gel 4%–20% (Bio-Rad Laboratories) and transferred onto polyvinylidene fluoride membranes (Invitrogen) using iBlot gel transfer stacks. Membranes were incubated with antibodies for beta-catenin (sc-7199; Santa Cruz Biotechnology), RhoGDI (clone 16; BD Transduction Laboratories), or tubulin (clone DM1A; Sigma) for 1 h in TBST (Tris-buffered saline, Tween 20) containing 5% nonfat dry milk, followed by 45 min incubation with horseradish peroxidase-conjugated goat-anti-mouse (Pierce) or goat-anti-rabbit (Dako) secondary antibodies. Immunoreactive bands were revealed using an enhanced chemiluminescence kit (ECL; Pierce).
Hematopoietic and mesenchymal cocultures
Cultured adult and fetal MSCs were seeded in 24-wells plates at the density of 50,000 cells/well in MSC culture medium (Gibco). Stromal layers were irradiated at 12 Gy when confluent. Five thousand cord blood-derived CD34+ cells were cocultured with stromal layers using MyeloCult (H5100; Stem Cell Technologies, Inc.) supplemented with 10−6 M hydrocortisone (Sigma) in the presence of 25 μM IWP2 or dimethyl sulfoxide control at 37°C and 5% CO2. Half of the culture medium was refreshed weekly.
Hematopoietic colony formation
For colony forming unit (CFU) assays with cord blood-derived CD34+, HSPC were plated in methylcellulose medium (Methocult H4434; Stem Cell Technologies) in 24-well plates and incubated at 37°C and 5% CO2. After 12–14 days, the colonies were counted.
Statistical analysis
Statistical significance was determined by the Mann–Whitney-U test, using SPSS 15.0 (SPSS, Inc.) and by the (un)paired Student's t-test.
Results
Gene expression profiling of ABMSC and FBMSC
To determine differences in gene expression programming between ABMSC and FBMSC, we cultured MSC from adult and fetal bone marrow. Cultured cells derived from all donors showed a spindle-shaped morphology, differentiated toward osteoblasts and adipocytes upon induction of differentiation, and expressed CD73, CD90, and CD105 but not hematopoietic markers CD34 and CD45 as previously shown in Maijenburg et al. [37,38].
RNA was isolated from third passage ABMSC and FBMSC. Gene expression was analyzed using Affymetrix Human Exon 1.0 ST arrays and ANOVA on the normalized log2 probe intensities of six samples (three ABMSC and three FBMSC). The expression of 687 genes was at least twofold increased in either ABMSC or FBMSC (303 genes up in ABMSC, 384 upregulated in FBMSC; P < 0.05). Gene ontology-term analysis indicated an enrichment for genes involved in developmental and cellular processes, adhesion, and locomotion (Fig. 1A). The cluster cellular processes predominantly contained gene ontology terms associated with cell cycle processes and cell division. This is in agreement with the previously reported increased proliferation rate of fetal MSC compared to adult MSC [39]. In the group developmental processes, gene ontology terms were linked to morphogenesis of a variety of tissues, including skeletal morphogenesis.
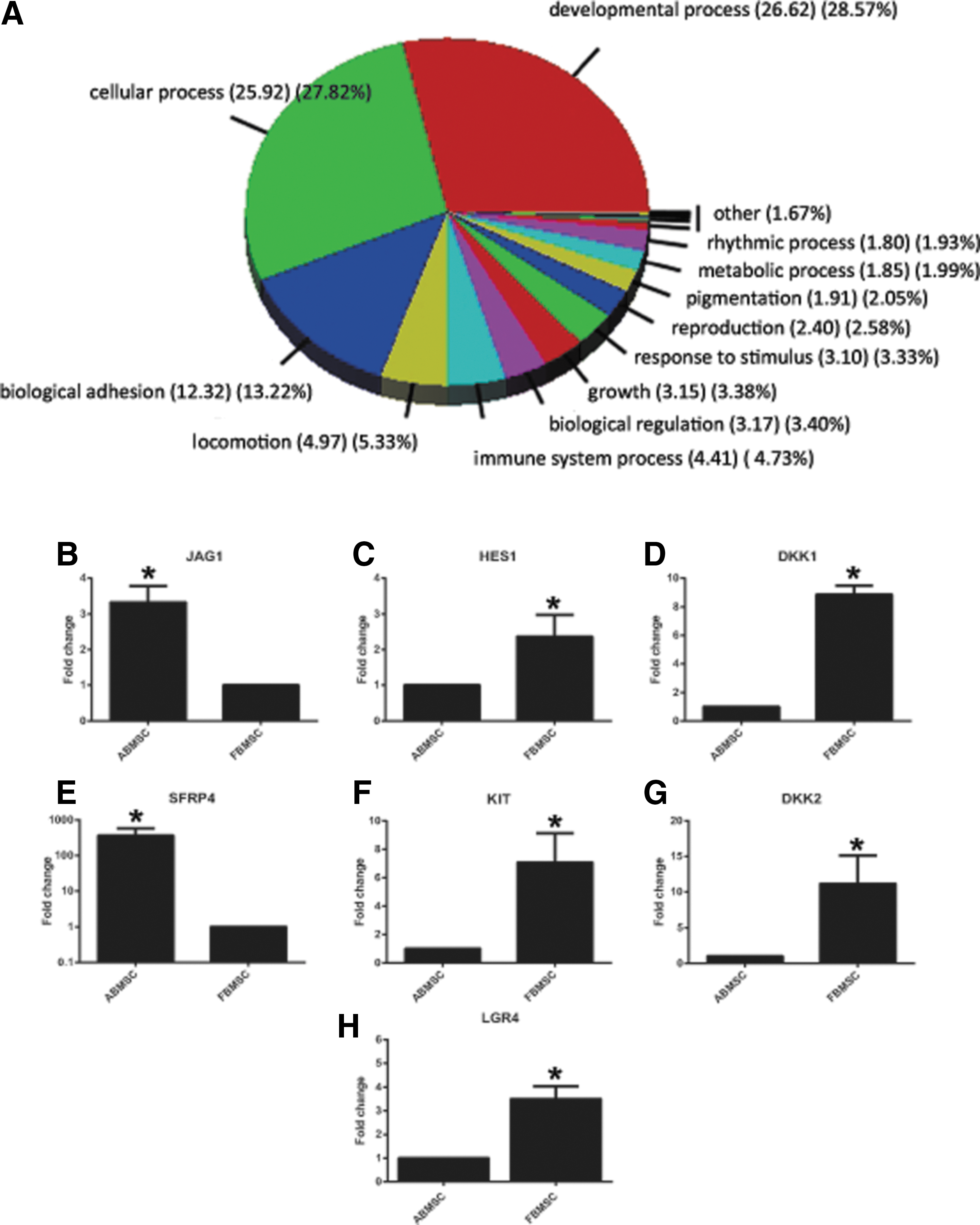
Functional enrichment analysis and confirmation of micro array data by RQ-PCR.
The 10 most differentially upregulated genes in ABMSC or FBMSC are listed in Table 1. Secreted frizzled-related protein-4 (SFRP4), an inhibitor of the Wnt pathway, was 50-fold upregulated in ABMSC compared to FBMSC. Because the Wnt pathway is involved in MSC differentiation [16 –18] and proliferation [16,19,25], we focused on genes that are involved in the Wnt pathway. These comprised 11 Wnt proteins, 13 Wnt receptors, and 5 Wnt inhibitors (Supplementary Table S2), which were detected above the cutoff value of the array from which eight genes were significantly higher expressed in ABMSC, whereas nine genes were increased in FBMSC (Table 1; Wnt target genes). DKK1 and DKK2 expression was enhanced in FBMSCs while SFRP4 expression was enhanced in ABMSC. They encode inhibitors that interact with the Wnt signaling pathway at different levels. DKK1 and DKK2 bind and block the LRP coreceptors, Whereas SFRPs sequesters Wnt proteins away from their cognate receptors [40]. The array data were successfully confirmed by RQ-PCR for 68 genes (data not shown) from which seven chosen and representative targets are depicted in Fig. 1B–H)
95% CI, 95% confidence interval; ABMSC, adult bone marrow-derived mesenchymal stromal cells; FBMSC, fetal bone marrow-derived mesenchymal stromal cells.
Cell culture-dependent expression of Wnt5a, DKK1, DKK2, and SFRP4
It has been demonstrated that expression of several Wnt proteins, Wnt target genes, and Wnt inhibitors in ABMSC is dependent on cell culture conditions [41]. To study whether the differential expression of DKK1, DDK2, Wnt5a, and SFRP4 was intrinsic to the developmental stage or the result of variation in cell division kinetics, we analyzed the expression of these genes in MSC cultures, which grew from low density to confluency. In ABMSC, confluency was reached at day 8–10, while FBMSC were confluent after 6–10 days. In addition, FBMSC underwent more cell divisions (Fig. 2A). DKK1 mRNA levels were high in low confluency cultures, and expression gradually decreased with increasing confluency of both ABMSC and FBMSC cultures (Fig. 2B). Thus, DKK1 expression was culture dependent, rather than cell type specific. Expression of DKK2 was much higher in FBMSC compared to ABMSC under all conditions tested, despite also being dependent on confluency (Fig. 2C). Gene expression levels measured by Q-PCR were related to the household gene Abelson kinase (ABL), which indicated that the expression level of DKK2 remained at least 10-fold lower than DKK1 (Fig. 2B, C). Similar to DKK1, Wnt5a expression appeared more dependent on confluency than on the cell type (Fig. 2D). SFRP4 expression, however, was consistently higher expressed in ABMSC under all conditions, although it also increased with culture confluency (Fig. 2E). Thus, the observed differences in expression of DKK2 and SFRP4 are intrinsic characteristics of adult and FBMSC.

Effect of cell culture on expression of Wnt-5a and Wnt inhibitors SFRP4, DKK1, and DKK2
The differential expression of Wnt-related genes comprised both canonical and noncanonical Wnts; therefore, we decided to focus on studying the Wnt pathway from a more general perspective.
Wnt3a-induced canonical Wnt signaling in adult and fetal MSC
To investigate whether there was a difference in canonical Wnt signaling between the two MSC types, we first investigated the stability of beta-catenin and the expression of the target genes LEF and TCF under steady-state conditions. Expression of beta-catenin protein and LEF and TCF mRNA differed among donors, but not between FBMSC and ABMSC in unstimulated conditions (Fig. 3A, B; expression compared to ABL mRNA).
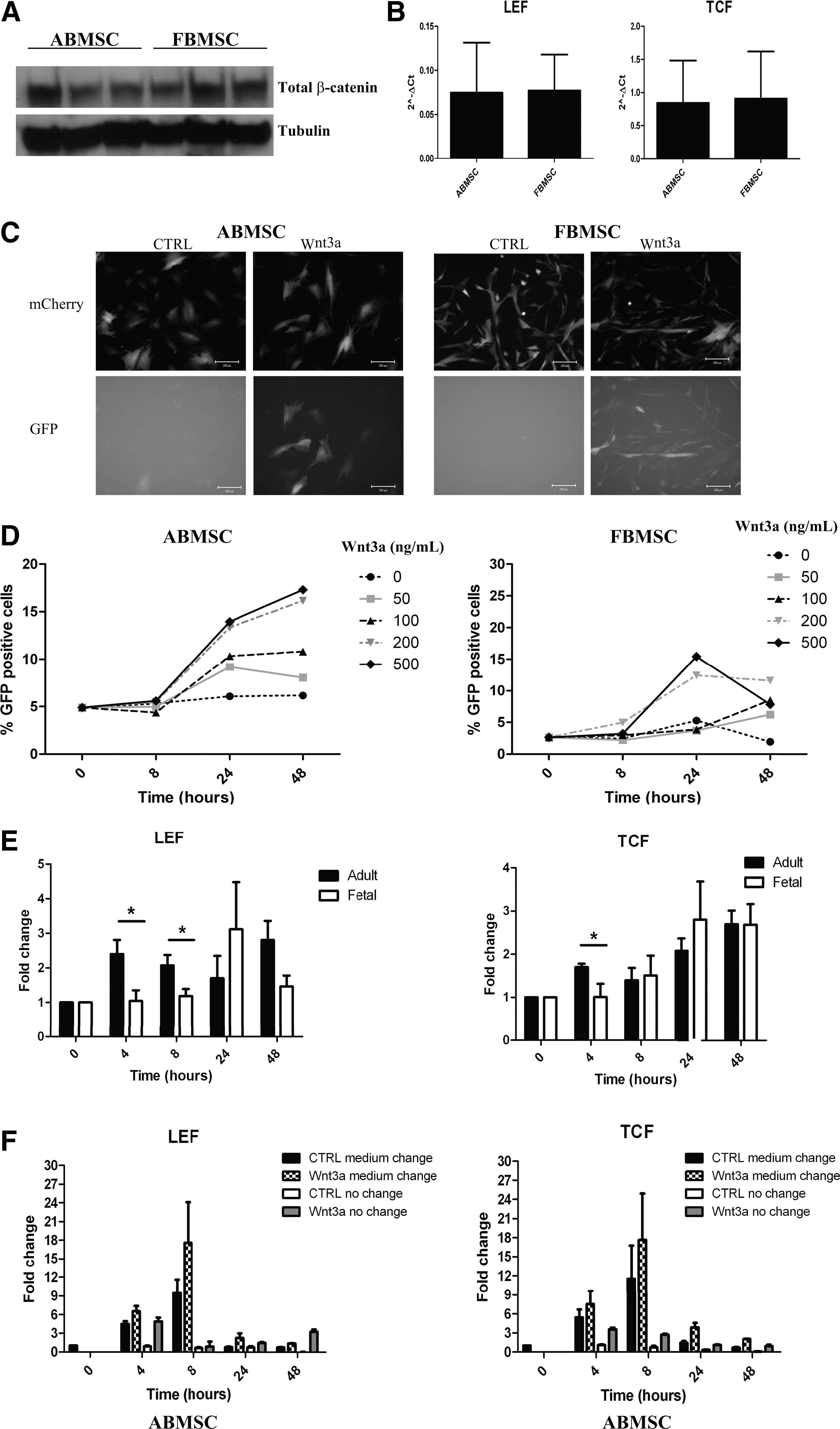
Wnt-3a induced canonical Wnt signaling in ABMSC and FBMSC.
Next, we measured activity of beta-catenin using TCF reporter constructs. MSC were transduced with the TCF reporter 7TGC (Fig. 3C, D) to assess both kinetics and functionality of the Wnt3a response. This reporter harbors a m-cherry marker to control overall plasmid expression and a GFP reporter gene whose expression is controlled by 7 copies of the beta-catenin binding site in the TCF promoter. The cells were stimulated with increasing amounts of Wnt3a up to 500 ng/mL. The percentage of GFP-positive cells increased with time- and concentration-dependent kinetics. ABMSCs were responsive to concentrations of 50 and 100 ng/mL, whereas FBMSC only responded to 200 and 500 ng/mL Wnt3a (Fig. 3D). In addition, the Wnt3a response was quenched 24 h after induction in FBMSC, but not in ABMSC suggesting a difference in negative feedback control (Fig. 3D). Thus, canonical Wnt signaling was induced in both ABMSC and FBMSC, although with different kinetics.
TCF and LEF expression was measured to study induction of endogenous genes by canonical Wnt signaling; Wnt3a induced a rapid increase in TCF and LEF expression in ABMSC, and a more delayed increase in FBMSC (Fig. 3E). Wnt3a was added to the medium in which the cells were being cultured, that is, in the presence of excreted inhibitors such as SFRP4. To examine whether excreted factors affect Wnt signaling, we stimulated ABMSC with Wnt3a in CM or in fresh medium. Induction of LEF and TCF by Wnt3a added to CM was maximal at 8 h (Fig. 3F). Notably, medium replacement induced both TCF and LEF expression independent of the addition of Wnt3a and surpassed the effect of Wnt3a addition to CM (Fig. 3F). These data indicated that secreted Wnt inhibitor(s) present in ABMSC-CM interfere(s) with exogenous Wnt3a and endogenously produced Wnts and prevent(s) maximal induction of TCF and LEF expression.
Secreted soluble factors influence MSC proliferation
To investigate whether the difference in proliferative capacity is due to the combination of excreted factors or due to an inherent difference in cell programming, we switched media between ABMSC and FBMSC (Fig. 4A). At days 3, 5, and 7 following seeding of adult or fetal MSC, the medium of both ABMSC and FBMSC was replaced by 50% fresh medium plus either 50% CM from ABMSC (ACM) or 50% CM from FBMSC (FCM). The addition of FCM to ABMSC significantly stimulated proliferation of adult cells at all indicated time points (Fig. 4B), whereas addition of ACM or FCM on FBMSC made no difference (Fig. 4C).

Secreted soluble factors influence MSC proliferation–Medium switch assay.
Inhibition of endogenous Wnt production
Given the positive effect of FCM on ABMSC, we questioned whether endogenous Wnt production and signaling by ABMSC and FBMSC contribute to the observed differences in growth kinetics between fetal and adult MSC. The endogenous Wnt production of ABMSC and FBMSC was inhibited by adding the small molecule IWP2 that prevents the palmitoylation of Wnt proteins by porcupine, which is required for Wnt secretion [35]. To control for the inhibitory effect of IWP2, L-cells overexpressing Wnt3a (L-Wnt3a) were incubated with IWP2 for 24 h. IWP2 dose-dependently decreased the level of beta-catenin (Fig. 5A). Although both ABMSC and FBMSC express WNT genes, addition of IWP2 for 16 hrs decreased beta-catenin protein levels in FBMSC, but not in ABMSC (Fig. 5B). Concordantly, overnight incubation of FBMSC with IWP2 decreased LEF and TCF mRNA expression (Fig. 5C, D), whereas no change was observed in ABMSC (Fig. 5E, F). Thus, autocrine Wnt signaling is important for canonical Wnt pathway activation in FBMSC, but not for ABMSC.
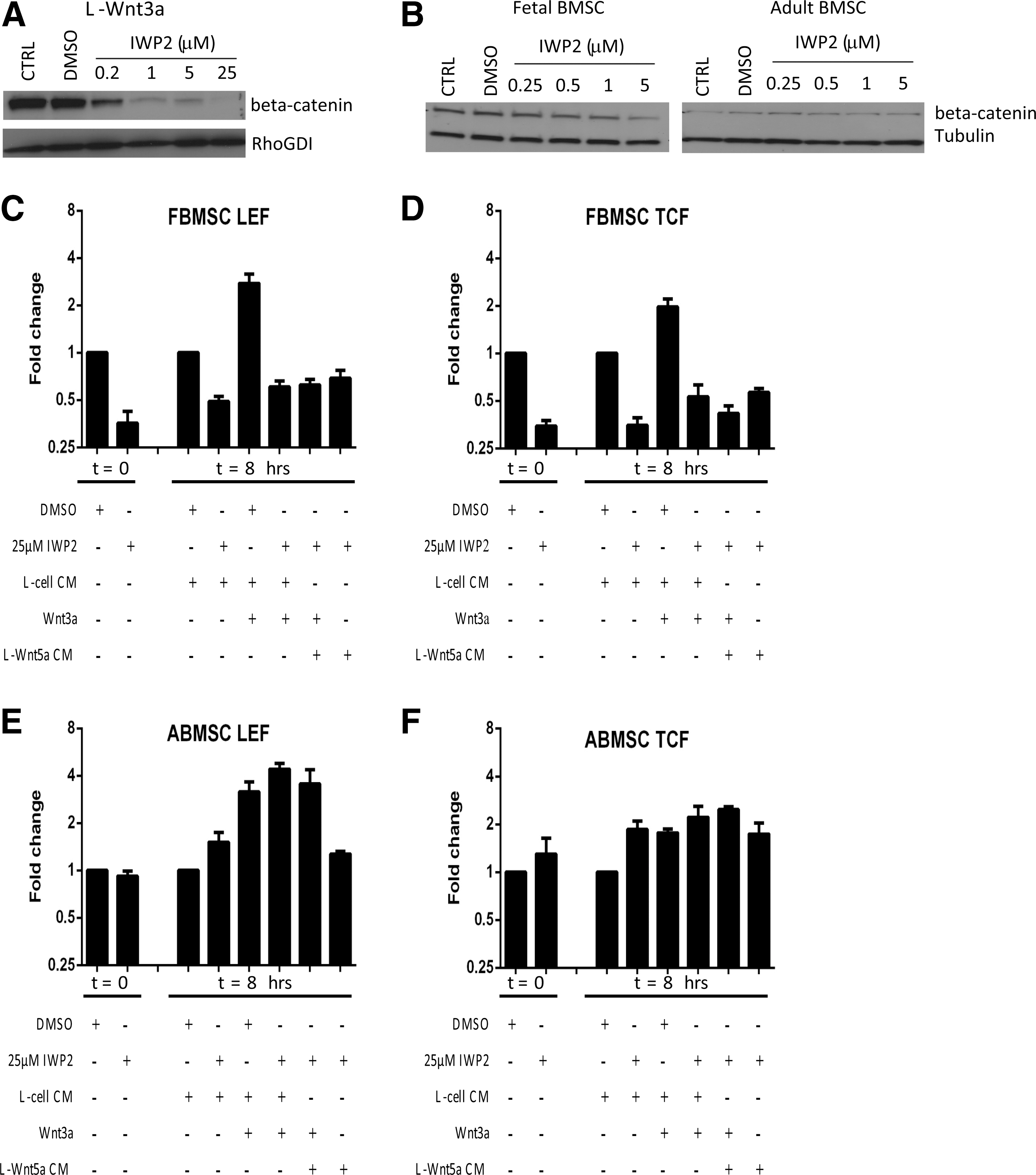
IWP2-mediated inhibition of endogenous Wnt signaling. Cell lysate of L-Wnt3a
IWP2 should not impair the induction of signaling by exogenously added Wnt factors. However, addition of Wnt3a or Wnt5a to FBMSC pretreated with IWP2 for 16 h could not induce LEF or TCF expression (Fig. 5C, D). Thus, endogenously expressed Wnt factors are involved in the effect exogenously added Wnt3a, or FBMSC react to other Wnt factors.
In contrast, ABMSC remained responsive to Wnt3a measured by induction of LEF and TCF expression (Fig. 5E, F). Possibly, IWP2 may have decreased production of noncanonical inhibitory Wnts [16].
Expansion of fetal and adult MSCs with IWP2 and Wnt3a
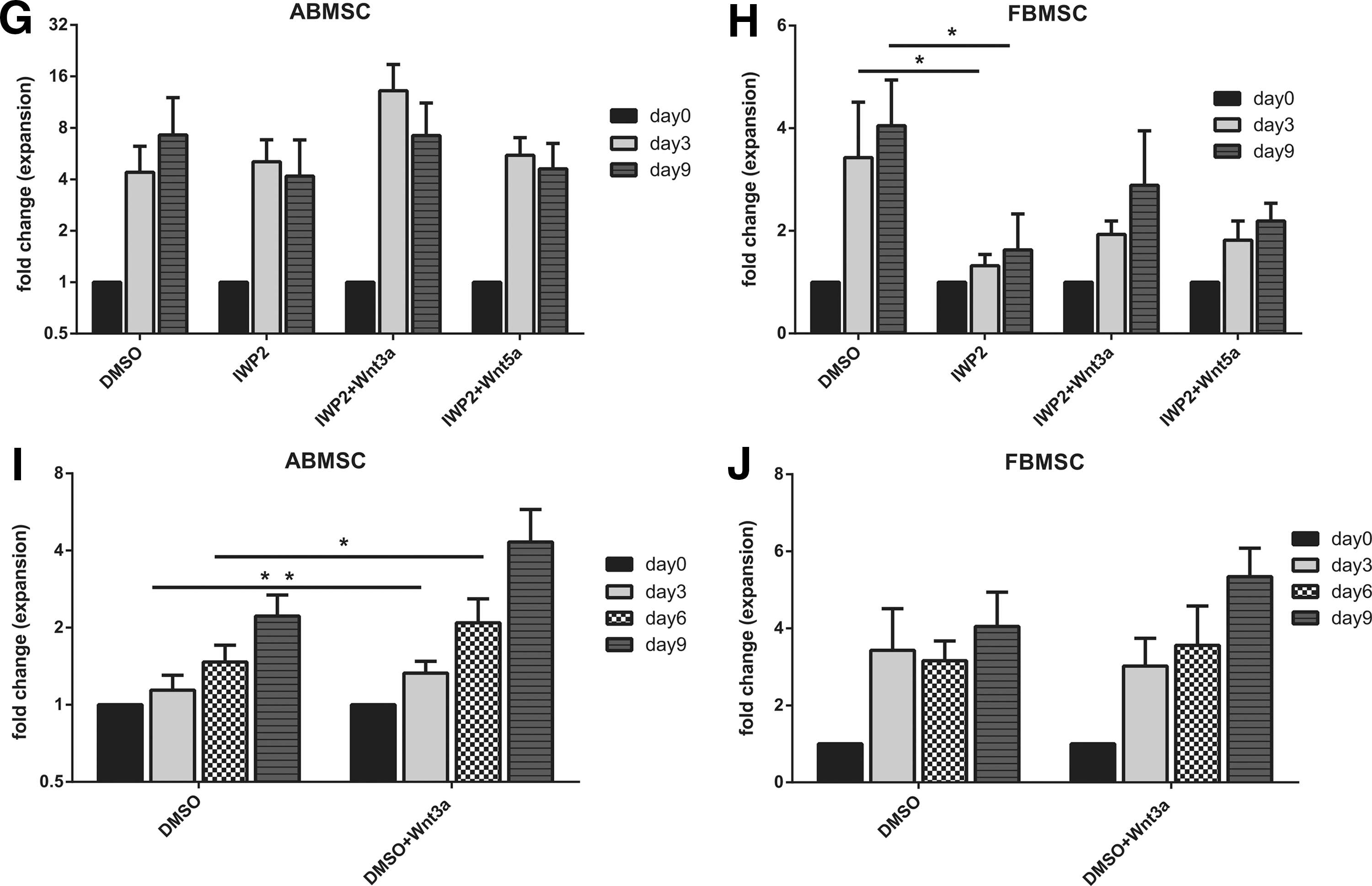
Knowing that Wnt signaling is crucial for cell expansion [16,19], we also studied the long-term proliferative capacity of both adult and fetal MSCs when exposed to IWP2 inhibitor and exogenous Wnt3a.
In accordance to short-term exposure, the addition of IWP2 to MSC cultures significantly inhibited expansion of FBMSCs, but not ABMSC (Fig. 5G, H). Addition of Wnt3a, in contrast, increased the proliferation of ABMSC, while FBMSC proliferation remained similar to the control situation (Fig. 5I, J). Proliferation of IWP-2-exposed fetal MSCs was not rescued by the addition of Wnt3a and Wnt5a (Fig. 5H), indicating that both do not affect expansion of fetal MSCs.
CM of IWP2 preincubated FBMSC inhibits proliferation of ABMSC culture compared to FCM
To assess whether the proliferation-stimulating effect of FCM was Wnt mediated, we exposed ABMSC to FCM of IWP2-exposed FBMSC. At day 3, ABMSC proliferation was significantly decreased in the presence of IWP2-preincubated FCM compared to control FCM (Supplementary Fig. S1A). Addition of either Wnt3a or Wnt5a did not restore the stimulatory effect of fetal CM on ABMSCs. This is surprising because Wnt3a alone enhanced ABMSC proliferation in all experiments thus far. Expression of both TCF and LEF was barely changed by FCM, independent of IWP2 treatment (Supplementary Fig. S1B).
Rspo-1–LGR4/5 pathway in MSC expansion
R-spondin-1 is a small secreted growth factor that binds its cognate LGR4 and −5 receptor, which stabilizes Wnt receptors on the membrane and potentiates Wnt signaling [25,26]. The different response of ABMSC and FBMSC to Wnt3a may be due to distinct activation of LGR4 or −5. We observed a 3.5-fold increased LGR4 expression in FBMSC compared to ABMSCs. (Fig. 1H). In addition, expression of R-spondin-1 (RSPO1) was 1.6-fold increased in adult MSCs when compared to fetal MSCs (microarray data not shown).
However, addition of R-spondin-1 in combination with Wnt3a to MSC cultures did not yield any synergistic/additive effect on either adult or fetal cell expansion (Supplementary Fig. S1C, D). Thus, differential LGR4 expression seems not to underlie differences in proliferation between adult and fetal MSCs.
Endogenous Wnt signaling in HSPC support
In the bone marrow niche, both MSC and the Wnt family molecules were implicated in the regulation of hematopoiesis [42]. Therefore, we studied the hematopoietic support capacity of both ABMSC and FBMSC in coculture with cord blood-derived CD34+ HSPC in the absence and presence of IWP2. ABMSC showed a significantly better hematopoietic support as determined by cell counts and total colony forming capacity [CFU-granulocyte-monocyte (GM), burst forming unit-erythroid (BFU-e), CFU-granulocyte, erythrocyte, monocyte, megakaryocyte (GEMM)] compared to FBMSC (Fig. 6A, B). IWP2 decreased colony formation in the presence of ABMSCs, whereas hematopoietic support provided by FBMSCs remained unchanged (Fig. 6B). Mainly CFU-GM colonies were detected, the numbers of BFU-e and CFU-GEMM colonies were too low to draw conclusions on skewing of the differentiation potential. Thus, secreted Wnt proteins are necessary for hematopoietic support by ABMSC.
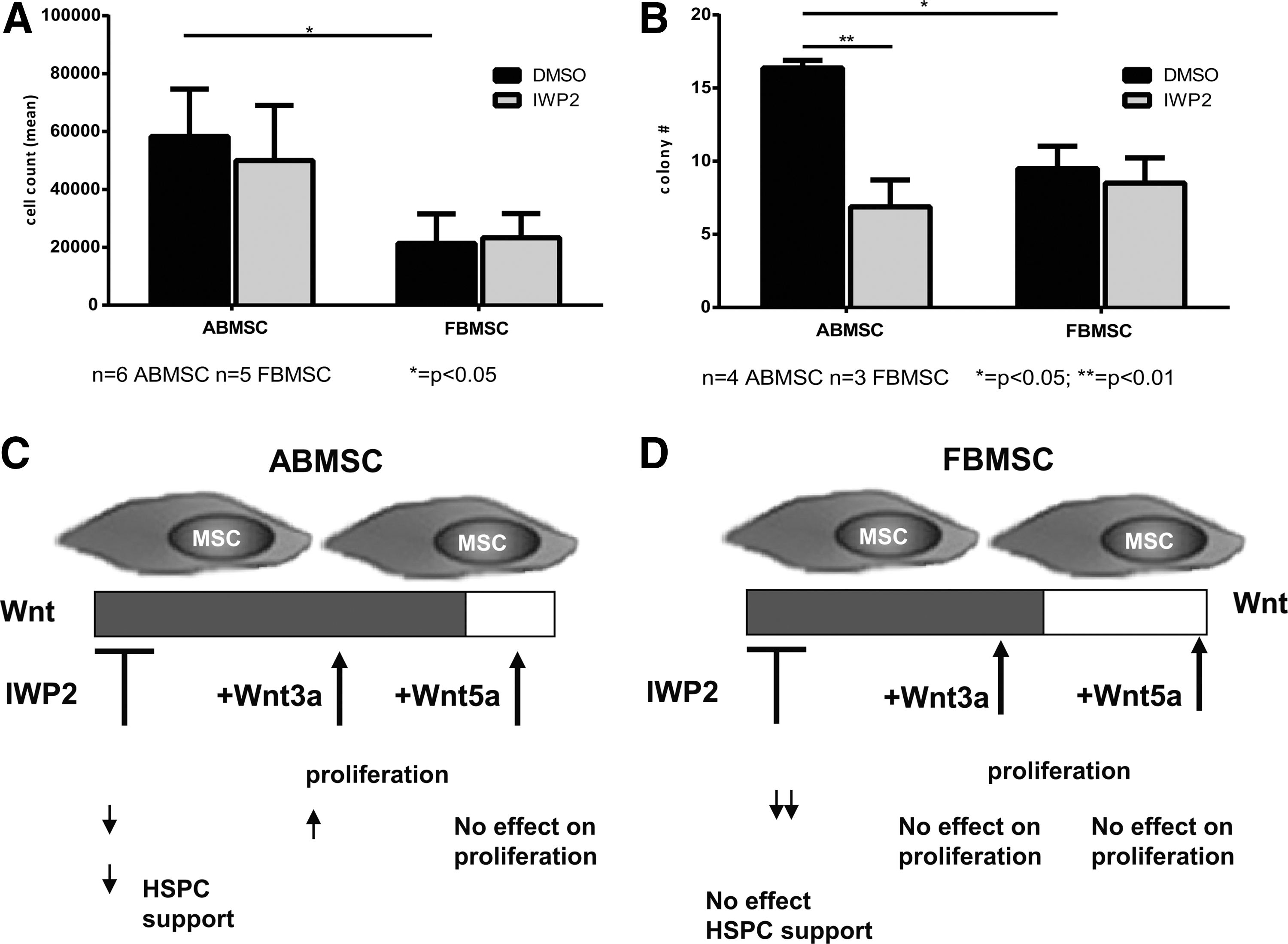
Endogenous Wnt signaling in HSPC support.
Discussion
Clinical use of MSC requires a robust cellular expansion capacity without loss of MSC function. Compared to ABMSC, FBMSC proliferate faster [39,43]. We explored the functional differences between ABMSC (obtained from adult donors in the age range between 40–70 years) and FBMSC (15–20 weeks gestation) with emphasis on genes encoding proteins in the Wnt pathway. Autocrine Wnt signaling was specifically important for the enhanced proliferation of FBMSC, but not for the proliferation of ABMSC. The ABMSC, instead, rapidly respond to exogenous Wnt3a and to medium replacement, which is in concordance with the high expression of the inhibitor SFRP4 in ABMSC. Despite an autocrine Wnt signaling loop, FBMSC do not provide Wnt-dependent HSPC support in cocultures. Hematopoietic support by ABMSC, in contrast, was Wnt dependent. Thus, we uncovered fundamental differences in the role of Wnt signaling in MSC proliferation and hematopoietic support, and in its function in FBMSC and ABMSC.
ABMSC and FBMSC identity
Gene expression profiling of FBMSC and ABMSC uncovered 687 genes to be differentially expressed between ABMSC and FBMSC. Functional gene enrichment analysis revealed that genes associated with limb and skeletal muscle development; cell cycle regulation and DNA-repair were enriched in FBMSC compared to ABMSC. Several genes that were differentially expressed in our microarray, such as intercellular adhesion molecule-1 (ICAM1, increased in FBMSC), vascular cell adhesion molecule-1 (VCAM1), and HLA-DR (both upregulated in ABMSC), were also differentially expressed in a similar pattern between fetal liver-derived MSC and ABMSC in the study of Gotherstrom et al. [44]. Thus, fetal MSC share some intrinsic characteristics that discriminate them from ABMSC and are independent of the fetal tissue of origin.
Although the developmental ontogeny of MSC is unknown, there are several indications that adult and embryonic or fetal MSC arise from phenotypically distinct cells. In the murine system, the first wave of MSC is derived from Sox1+ neural crest cells that are transiently replaced by cells with an undefined ontogeny [45]. Also in the human system, different subpopulations of MSC seem to exist during development and aging. In the fetal bone marrow at 15–20 weeks of gestation, CFU fibroblast (a measure for clonogenic MSC in bone marrow) can be derived from CD146+ bone marrow cells independent of CD271 expression [46]. In adult bone marrow, however, clonogenic cells only reside in CD271+ cells independent of CD146 expression [46,47]. FBMSC may be a more immature state of ABMSC, but it is equally well possible that it is a distinct cell type.
Effects of differential Wnt signature
Despite differences in their Wnt signature, Wnt3a activated basal canonical Wnt signaling in both ABMSC and FBMSC as indicated by upregulation of previously described beta-catenin targets TCF and LEF [19,47,48]. The kinetics of the response, however, was different. LEF and TCF expression increased earlier in ABMSC, and these cells were able to respond to lower concentrations of Wnt3a. In addition, in ABMSCs, canonical pathway seemed to be active longer than in fetal MSCs, which after 24 h rapidly quench canonical pathway's response. Attempts to analyze the induction of canonical signaling by means of increased beta-catenin levels have been inconclusive in our hands. This may be explained by the relatively large amount of beta-catenin present in the cytoskeleton and cell–cell junctions of MSC. Therefore, the minor proportion of Wnt3a-induced beta-catenin accumulation is difficult to detect, a problem also encountered by others [49].
Differential Wnt sensitivity and its effect on cell proliferation
Addition of Wnt3a and inhibition of endogenous Wnt production by IWP2 indicate that the balance in Wnt signaling as well as the autocrine signaling route differs between ABMSC and FBMSC. ABMSC but not FBMSC produce soluble Wnt inhibitors that prevent(s) induction of TCF and LEF expression by endogenous Wnt proteins, as indicated by the upregulation of LEF and TCF mRNA upon medium refreshment. The prime candidate is SFRP4, a soluble Wnt receptor of which the corresponding mRNA is 50-fold upregulated in ABMSC compared to FBMSC. Addition of Wnt3a could overcome the inhibitor levels. FCM contained sufficient Wnts/factors to overcome inhibitor production by ABMSC, because FCM induced proliferation of ABMSC. Surprisingly, addition of Wnt3a, Wnt5a, or combination of Wnt3a and Wnt5a (data not shown) to FCM of IWP2-treated FBMSC did not restore its positive effect on ABMSC proliferation. It suggests that the soluble factor(s) responsible for enhanced expansion of ABMSC in FCM presence may not be Wnt3a or Wnt5a, but another (Wnt related) factor, and that Wnt3a may be quenched by inhibitors produced by FBMSC. This also fits with our observation that the inhibition of the canonical signaling in FBMSC by IWP2 could not be overruled by the addition of exogenous Wnt3a.
In ABMSC, IWP2 treatment did not affect the canonical signaling and exogenous Wnt3a was still able to increase TCF and LEF expression in IWP2 pretreated ABMSC. It is important to note that BMSC hardly produce canonical Wnts [33], and except for the noncanonical Wnt5a, none of the Wnt proteins was differentially expressed between ABMSC and FBMSC.
Both short- and long-term experiments indicated that abrogation of endogenous Wnt signaling affects proliferation of fetal MSCs much more than ABMSC and cannot be rescued by exogenous Wnts. In contrary, ABMSCs remained responsive to exogenous stimulus and their proliferation was positively influenced by Wnt3a. It is therefore possible that addition of Wnt3a shifts the balance of Wnt signaling in those cells and is able to overrule the effect of potent inhibitors, which ABMSCs secrete.
Our data indicate that the net result of Wnt signaling in FBMSC results in canonical activation, while in ABMSC, the balance may result in repression of the canonical pathway.
Although an effect of IWP2 on the previously described noncanonical Wnt targets such as the small GTPase Rac1 [50] JNK [17] and the Src family kinases [51] could not be demonstrated because the amount of Rac-GTP, p-JNK, and p-SRC protein in MSC is relatively low, we cannot exclude that IWP2 treatment can affect Wnt signaling through these targets.
Niche perspective and MSC-HSPC interaction
It is of interest that FBMSC medium or secreted products but not ABMSC promote MSC expansion through an IWP2-dependent autocrine loop, while ABMSC but not FBMSC support expansion of HSPC through an IWP2-dependent paracrine stimulation. This poses the question whether the autocrine and paracrine effects are executed by distinct Wnt factors, or whether the presentation of Wnt factors differ. It has recently been shown that Wnt factors can be excreted in exosomes [52]. This would be an effective mechanism to discriminate the autocrine and paracrine signaling. It enables ABMSC to inhibit Wnt autocrine signaling by excreted inhibitors such as SFRP4 that have to maintain steady state cell numbers, without affecting the paracrine effect that supports expansion of HSPC.
Overexpression or knockout in osteoblastic and stromal cells of the Wnt inhibitors DKK1 or SFRP1, respectively, resulted in an increased proportion of proliferating HSC that upon serial transplantation could no longer reconstitute the hematopoietic system in irradiated wild-type hosts [53,54]. Coculturing HSC on MSC that are transduced with a dominant active form of beta-catenin enhanced their repopulation capacity [31], whereas beta-catenin-deficient stroma has impaired supportive capacity [32]. These studies demonstrate that a precisely balanced level of Wnt signaling in stromal cells is important for hematopoiesis. While the effect of genetic modifications that enhance or decrease Wnt signaling has yielded contradictory results, studies that balance Wnt signaling confirmed this need for the optimal concentration [55].
In conclusion, our data demonstrate that ABMSC (donor age 40–70 years) and FBMSC (15–20 weeks gestation) have a different Wnt signature, which may be a reflection of their developmental ontogeny (Fig. 6C, D). Both MSC sources can be characterized by a certain level of Wnt signaling. They react differently to exogenous Wnts and in early developmental stages (FBMSC) rely on Wnt proteins for their expansion benefit. In adult life (ABMSC), proliferation of MSCs becomes much less Wnt dependent. In the same time, in contrary to early life, Wnt-related interaction is highly supportive for MSC-HSPC axis. In addition, in our hands, fetal-derived CM showed beneficial stimulatory effect on ABMSCs' proliferation. Similarly, Ng et al. [56] demonstrated that FBMSC-derived extracellular matrix can also positively influence both proliferation and differentiation of ABMSCs. It is noteworthy to mention that large-scale expansion of ABMSC has always been challenging. Therefore, determining secreted factor(s) residing in FCM and being responsible for enhanced expansion of ABMSCs' may ultimately lead to improvement of clinical protocols.
Footnotes
Acknowledgments
M.W.M. is financially supported by DPTE grant no. 06728. M.M.P. is financially supported by PPOC-11-021 grant. C.V. is financially supported by LSBR grant F1101. The authors thank S. van Reijmersdal for technical support and Dr. C. Fuerer and Prof. R. Nusse for the 7TGC construct. They also thank the staff of the Bloemenhove Kliniek (Heemstede, the Netherlands) for providing fetal tissues. They would also like to thank B. Nota for additional statistical analysis.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.