
Abstract
It has been suggested that hematopoietic stem/progenitor cells (HSPCs) could become specified from a population of migrating primordial germ cells (PGCs), precursors of gametes, during embryogenesis. Some recent experimental data demonstrated that the cell population that is usually considered to be PGCs, moving toward the gonadal ridges of an embryo, contains a subset of cells coexpressing several germ cell and hematopoietic markers and possessing hematopoietic activity. Experimental data showed that bone morphogenetic protein 4 (BMP4) generates PGCs from mouse bone marrow-derived pluripotent stem cells. Interestingly, functional reproductive hormone receptors have been identified in HSPCs, thus indicating their potential role in reproductive function. Several reports have demonstrated fertility restoration and germ cell generation after bone marrow transplantation in both animal models and humans. A potential link between HSPCs and germinal lineage might be represented by very small embryonic-like stem cells (VSELs), which have been found in adult human bone marrow, peripheral blood, and umbilical cord blood, express a specific pattern of pluripotency, germinal lineage, and hematopoiesis, and are proposed to persist in adult tissues and organs from the embryonic period of life. Stem cell populations, similar to VSELs, expressing several genes related to pluripotency and germinal lineage, especially to PGCs, have been discovered in adult human reproductive organs, ovaries and testicles, and were related to primitive germ cell-like cell development in vitro, thus supporting the idea of VSELs as a potential link between germinal lineage and hematopoiesis.
Introduction
S
A potential link between HSPCs and germinal lineage might be represented by very small embryonic-like stem cells (VSELs), which have been found in adult human bone marrow [5,6], peripheral blood [7], and umbilical cord blood [8]. It has been suggested that VSELs, similar to PGCs, originate from epiblast during embryonic development, are in a quiescent state, and persist in human adult tissues and organs [9]. VSELs seem to be epigenetically locked to prevent teratoma formation in human adult tissues and organs [10]. These small stem cells have garnered considerable attention and are very interesting for different kinds of medicine, from regenerative medicine and oncology to reproductive medicine.
The aim of this review is to summarize the experimental evidence on the potential developmental link of germinal and hematopoietic lineages in mammals, including humans.
PGCs and HSPCs
Human preimplantation development may be supposed to begin with PGCs, which, surprisingly, arise outside the genital ridge region and can first be identified in the human embryo at about 3 weeks in the yolk sac epithelium near the base of the developing allantois [11]. They are recognized by their distinctive morphology showing blebbing and pseudopodial extrusions on the surface, alkaline phosphatase (AP) activity, and surface expression of stage-specific embryonic antigen 1 (SSEA-1) [1,11]. PGCs are generated in the region of epiblast, an embryonal layer that arises from the blastocyst inner cell mass and otherwise gives rise to the somatic cell lineages (endoderm, mesoderm, and ectoderm) that make up the proper embryo during gastrulation in mammals [11]. The specification of PGCs in mammals is mainly attributed to two signaling pathways: (1) the canonical WNT/β-catenin pathway and especially (2) the bone morphogenetic protein (BMP) signaling pathway, which activates expression of the transcription factors Blimp-1/Prdm1 and Prdm14, as demonstrated in mice [12,13].
The expression of Blimp-1 is the earliest known sign of PGC specification, and its mutation results in the generation of PGC-like cells, which resemble the surrounding somatic cells more than the real PGCs [14]. It has been found that the central role of Blimp-1 is the induction of transcription factor Tcfap2c, which regulates the expression of somatic genes (including mesodermal markers). The mutants for this transcription factor exhibit an early loss of PGCs [15 –17]. Similarly, the mutation of Prdm14 results in the loss of PGCs because of failure in the global erasure of histone 3 methylation patterns [18,19]. Both Blimp-1 and Prdm14 are also involved in another epigenetic event that causes global DNA demethylation of PGCs [20]. Actually, Blimp-1 and Prdm14 activate PGC specification by promoting germline development and pluripotency. They positively regulate the expression of pluripotency-related genes, such as Sox2, Nanos3, Nanog, Stella, and Figla, and inhibit the expression of the Hox family genes, which are related to somatic differentiation [12]. In this way, they prevent the PGCs from taking on a somatic fate.
Despite that, the mechanisms of human PGC specification from germline-competent pluripotent stem cells remain poorly understood. Human PGCs and germinal tumors (eg, seminoma) share the expression of CD38, a cell surface marker, which collectively defines the likely progression of the early human germline. A current study has demonstrated that SOX17 is the key regulator of human PGC fate, whereas BLIMP-1 represses endoderm-specific and other somatic genes during specification of human PGCs [21]. Recently, human induced pluripotent stem (iPS) cells were successfully in vitro differentiated into incipient mesoderm-like cells, which then robustly generated the human PGC-like cells that were subsequently purified using the surface markers, EpCAM and INTEGRIN α6, and shown to be quite comparable to nonhuman primate PGCs in terms of gene expression [22]. It has also been demonstrated that BLIMP-1 activated and stabilized the germline transcriptional circuit and repressed a default neuronal differentiation program in these cells.
The PGC population expands by mitosis and migrates by ameboid movement to the connective tissue of the hindgut and then on to the gut mesentery. Approximately 30 days after fertilization, the majority of the PGCs pass into the region of the developing kidneys and then to the adjacent primordial gonads where they, based on chemotaxy, join the cells of the sex and medullary cords [11]. Throughout the PGC migration and early colonization, it is not possible to discriminate between the female and male gonads; thus, any reference is to the indifferent gonads. In the human embryo, the PGC colonization is completed during the sixth week of gestation; after that time, the female gonad—the ovary—begins its development in the absence of chromosome Y, namely the gene SRY, where the PGCs start to develop into female gametes. If chromosome Y is present, testicles start to develop, and the PGCs transform into male gametes.
PGCs are neither unique nor uniform but a heterogeneous population of cells with different properties. The mouse migratory PGCs are heterogeneous for the expression of integrin α6 and c-Kit receptor [23]. PGCs with a negative or low integrin α6 expression and with side population cell phenotype showed a higher potential to convert to pluripotent embryonic germ (EG) cells. Some recent experimental data from the mouse model also indicated that the cell population (usually regarded as PGCs) moving toward the gonadal ridges contains a subset of cells both coexpressing several germ cell and hematopoietic markers and possessing hematopoietic activity [24]. Various OCT3/4-positive subpopulations of cells, which had settled in the AGM region, showed a distinct expression of PGC markers, such as BLIMP-1/PRDM1, AP, TG-1, and STELLA/DPPA3, as can be seen in Table 1, and coexpressed several proteins (CD34, CD41, and FLK-1) and genes (Brachyury, Hox-B4, Scl/Tal-1, and Gata-2) of hematopoietic precursors. Moreover, these cells possessed robust hematopoietic colony-forming activity in vitro [24].
+, expressed; −, nonexpressed; +/−, expressed/nonexpressed; ND, nondefined; ESCs, embryonic stem cells; PGCs, primordial germ cells; VSELs, very small embryonic-like stem cells.
Hematopoietic cells from PGCs
The above findings are in accordance with some earlier studies. Mouse PGCs from the embryonal yolk sac, positive for AP and SSEA-1, have been grown on an embryonic fibroblast feeder cell layer in the presence of leukemia inhibitory factor, stem cell factor, and interleukin-3 (IL-3). The individual colonies, taken from day 5 cultures, have differentiated into erythroid lineage cells in secondary methylcellulose culture. Interestingly, the cells from these colonies have formed cobblestone areas on irradiated adult bone marrow stromal layers, thus indicating that the most primitive hematopoietic stem cells in vitro, the cobblestone area-forming cells, were present in the cell cultures. In this way, it has been demonstrated that PGCs can be induced to differentiate into hematopoietic cells when cultured in methylcellulose and in the presence of IL-3 [1]. Furthermore, it has been suggested that PGCs represent a totipotent stem cell population capable of producing a variety of different cell types, including cells of the hematopoietic system. The same was found in another study, where the induction of hematopoiesis in an EG cell line, derived from mouse PGCs, was examined [25]. When single undifferentiated EG cells were seeded into the methylcellulose medium, both primitive and definitive erythropoiesis were identified in EG-derived embryoid bodies, similar to the ones in embryonic stem cells (ESCs), and production of myeloid cell lineages was induced by IL-3 [25]. These results further indicated that EG cells possess the potency to differentiate toward hematopoietic cells in vitro.
PGCs from bone marrow-derived pluripotent stem cells
Upon further consideration, the above findings may be supported by the fact that the opposite process has also been observed. It has been clearly demonstrated that BMP4 can generate PGCs from mouse bone marrow-derived pluripotent stem cells [26]. The SSEA-1-positive cells were directly separated from the suspension of murine mononuclear cells, which were harvested from adult bone marrow of 2- to 4-week-old mice. The flow cytometry and reverse transcription PCR analyses, after differentiation of SSEA-1-positive cells into cells of all three germ layers, demonstrated the pluripotency properties of isolated cells. The SSEA-1-positive cells were induced to differentiate into the germ cell lineage by adding BMP4 to the culture medium. According to the expression of germ cell markers, it was found that adding exogenous BMP4 to the culture medium could differentiate adult bone marrow-derived pluripotent SSEA-1-positive cells into PGCs expressing specific molecular markers, such as Oct4, Figla, Stella, and Mvh (mouse Vasa homolog) [26].
Very Small Embryonic-Like Stem Cells
The VSELs were initially discovered by Ratajczak and colleagues in human adult bone marrow [5,6], peripheral blood [7], and umbilical cord blood [8], and they have also been confirmed by several other groups [27 –36] despite some skepticism [37]. The VSELs are indeed small cells with diameters of <3.6 μm (3–5 μm) and are smaller than HSPCs, which have diameters of <6.5 μm (6–7 μm). They can be efficiently isolated from tissues by fluorescence-activated cell sorting (FACS) [2]. It has been suggested that VSELs, similar to PGCs, originate from epiblast during embryonic development, are in a quiescent state, and persist in human adult tissues and organs from the embryonic period of life [4,9]. Their molecular status has already been well characterized in both humans and mice [38 –40]. Evidence shows that VSELs are related to the germinal lineage and express several markers common to PGCs, such as fetal-type AP, OCT4, OCT4A, SSEA-1, CXCR4, DDX4/VASA, DPPA3/STELLA, FIGLA, NOBOX, and HDAC6. These data indicated that they could be indeed developmentally related to a population of migrating epiblast-derived PGCs.
It has been suggested that VSELs are deposited in the bone marrow during ontogenesis. The bone marrow development occurs by the end of the second/beginning of the third trimester of gestation in mammals and represents a major hematopoietic organ in postnatal life. The CXCR4–SDF-1 axis plays a major role in the colonization of bone marrow by stem cells through the end of the second trimester of gestation. Hematopoietic stem cells are chemically attracted from the fetal liver. Interestingly, evidence also shows that VSELs express the CXCR4 and SDF-1 markers and respond to the SDF-1 gradient [41]; therefore, they may take part in the colonization of bone marrow by stem cells. They expressed hematopoietic stem cell marker SCA-1 but not CD45, thus indicating another origin.
In several studies, it has been demonstrated that VSELs express a specific pattern of pluripotency, including markers SSEA-1, SSEA-4, OCT4, NANOG, KLF4, and REX1. In contrast, these VSELs were confirmed to be negative for markers of mesenchymal stem cells (eg, CD90, CD105, and CD29). They could be mobilized from bone marrow to peripheral blood in response to tissue/organ injury to regenerate the damaged area [7]. These small stem cells might be involved in tumor formation when they are mobilized at the wrong time and place or under inappropriate conditions in the body [42]. Otherwise, VSELs appear to be epigenetically locked to prevent teratoma formation in human adult tissues and organs in the healthy condition [10].
The VSELs are very promising for regenerative medicine. The experimental data, mostly from animal models, show that VSELs are not only able to differentiate into hematopoietic lineage in vitro when cocultured over stromal cells [43] but can also be differentiated into lung epithelium [31] and are involved in hepatic [33] and pancreatic [35,44,45] regeneration. Human and murine VSELs represent multipotent tissue progenitors in vitro and in vivo and can differentiate into cells of all three embryonic germ layers [36]. The VSELs are present in various somatic adult organs and tissues [46,47], and mobilization of VSELs into peripheral blood can occur when the function of organs, such as the heart (myocardial infarction) [30,48 –53], pancreas [54], brain [55 –58], liver [33], lungs [31], bowel [59], and skin (burns) [60], is compromised.
Functional Reproductive Hormone Receptors in HSPCs and VSELs
In some studies, HSPCs responded with clonogenic growth in vitro to certain reproductive hormones, such as prolactin, androgens, and estrogens [61 –63]. The terminal differentiation and function of some hematopoietic cells are regulated by reproductive hormones and are gender related [61]. Mouse HSPCs exhibited gender differences in cell-cycle regulation in the presence of estrogen. In female mice, they divided significantly more frequently than in male mice. This difference depends on the presence of ovaries but not testes. Administration of estradiol, a hormone produced mainly in the ovaries, increased HSPC division in both male and female mice. Interestingly, estrogen levels increased during the pregnancy and had an increased effect on HSPC division, its frequency, cellularity, and erythropoiesis in the spleen in female mice [61]. It has been found that HSPCs express high levels of estrogen receptor α (ER-α). The conditional deletion of ERα from them reduced hematopoietic stem cell division in female but not in male mice [61]. It has been demonstrated that estradiol acts directly on bone marrow myeloid progenitors to differentially regulate GM-CSF or Flt3 ligand-mediated dendritic cell differentiation [62]. Androgens are currently used to treat aplastic anemia patients [63]. The stimulatory effects of testosterone on erythropoiesis are well known, but the mechanisms underlying the erythropoietic action of testosterone are still poorly understood. Erythropoietin has long been considered as a potential mediator, but a clinical study showed that transdermal testosterone treatment of older men for 36 months significantly increased hemoglobin but not erythropoietin levels [64]. Therefore, the hematopoietic effect of testosterone did not appear to be mediated by stimulation of erythropoietin production.
Recent research reports the first incidence that human HSPCs express functional gonadotropin—follicle-stimulating hormone (FSH) and luteinizing hormone (LH)—receptors and that both types of receptors proliferate in vivo and in vitro in response to stimulation by pituitary hormones FSH and LH [65]. Furthermore, based on observations that at least some CD45-negative VSELs from bone marrow may become specified into CD45-positive HSPCs, the expression of pituitary and gonadal FSH and LH receptors was tested in these cells. In addition, testing is being carried out whether otherwise quiescent VSELs may expand in vivo in response to administration of reproductive hormones FSH and LH. It has been found that human VSELs express receptors for reproductive hormones, FSH and LH, and that they respond in vivo to stimulation with these hormones, as evidenced by BrdU accumulation. The authors have concluded that at least some VSELs share several markers characteristic of migrating PGCs and that they can be specified into HSPCs.
VSELs and Generation of Germ Cells in Adult Human Reproductive Organs
Regarding the suggestion that VSELs persist in adult human tissues and organs from the embryonic period of life, it is expected that they are also present in human adult reproductive organs. Indeed, small stem cells resembling VSELs have already been found in adult human ovaries and testicles.
VSEL-like cells in adult human ovaries
The research of stem cells in adult human ovarian surface epithelium (OSE) has been encouraged by the finding of Bukovsky et al. who have found that large cells expressing some oocyte-specific markers were developing in vitro in the OSE cell cultures of aged postmenopausal women from unknown progenitor cells [66]. Further experimental data showed that in adult human OSE, there is a population of small stem cells with diameters of 2–4 μm (Fig. 1), which resemble VSELs from adult human bone marrow. These small stem cells in the OSE were for the first time published by our group [67,68]. We have demonstrated that these small stem cells can be isolated from the remaining population of OSE cells by two different methods: magnetic-activated cell sorting and FACS, based on the expression of stage-specific embryonic antigen 4 (SSEA-4), a marker of pluripotent stem cells [69].
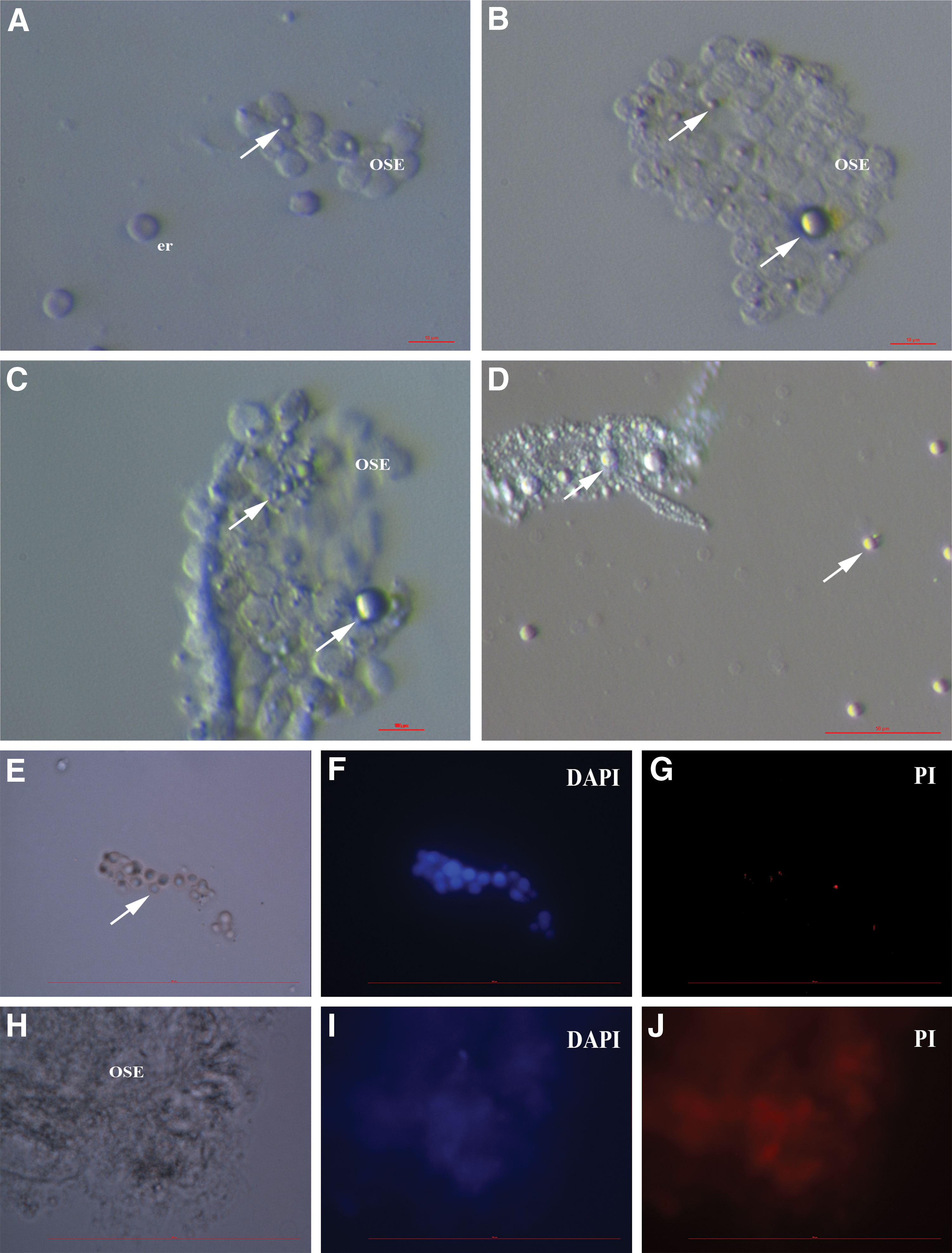
Human adult ovarian surface epithelium (OSE) scrapings retrieved by laparoscopic brushing of the ovaries in a reproductive-age patient with ovarian insufficiency.
These small stem cells from adult human ovaries were analyzed on a gene expression profile using microarrays and validated by quantitative real-time PCR. The data demonstrated that small stem cells from OSE expressed several genes related to pluripotency (eg, SALL4, CDH1, LIN28B, PUU5F1, NANOG, SOX11, LEFTY1, ZIC3, and HESRG), PGCs (eg, PRDM1/BLIMP-1, PRDM14, and STELLA/DPPA3), and germinal lineage (eg, VASA/DDX4 and PLD6); genes related to PGCs and germinal lineage were upregulated in comparison to human embryonic stem cells (hESCs) and adult dermal fibroblasts [70].
The spalt-like transcription factor 4 (SALL4) gene was among the most prominently expressed genes related to pluripotency in small ovarian stem cells. The recent findings show that the expression of Sall4 is essential for mouse PGC specification through the suppression of somatic cell program genes [71,72]. Interestingly, this gene regulates transcription in ESCs, hematopoiesis, and acute myeloid leukemia. Overexpression of this gene induces acute myeloid leukemia in mice [73]. It has been demonstrated that hematopoiesis is highly sensitive to appropriately balanced SALL4 expression.
Parte et al. have confirmed a similar population of small stem cells in adult human ovaries; these cells were referred to as the “VSELs” [74]. Such cells have also been found in adult ovaries of other mammalian species, such as monkeys [74], sheep [74], rabbits [74], mice [75], and pigs [76]. Moreover, a similar population of stem cells has also been found in mouse endometrial tissue [77].
Comparison of small stem cells from adult human ovaries, bone marrow-derived VSELs, PGCs, and ESCs
The small stem cells from adult human ovaries express similar dimensions, namely diameters, as VSELs from adult human bone marrow (2–4 vs. 3–5 μm) and the nucleus, which spreads over almost the whole-cell volume. They share several other characteristics, including the expression of pluripotency related to VSELs, PGCs, and ESCs, as can be seen in Table 1 [38 –41,66 –88]. The common characteristic of all these types of cells is the expression of a specific pattern of pluripotency. It needs to be recognized that small ovarian stem cells, VSELs, and PGCs strongly express markers related to germinal lineage, such as DPPA3/STELLA, DDX4/VASA, PRDM1/BLIMP-1, and PRDM14, while these markers are not expressed or are only weakly expressed in ESCs, as can be seen in Table 1. Despite the fact that VSELs express several genes related to pluripotency and germinal lineage, such as PGCs and small ovarian stem cells, they also express some markers related to hematopoietic cells, such as prominin-1 (CD133) and hematopoietic progenitor cell antigen (CD34) but not protein tyrosine phosphatase, receptor type C (CD45).
Development of oocyte-like cells in vitro
The experimental data showed that small VSEL-like stem cells, isolated from adult human ovaries, namely OSE, can grow and develop into primitive oocyte-like cells in vitro either spontaneously or in the presence of follicular fluid from the in vitro fertilization program, which contains several substances important for growth and development of oocytes [67,68,88]. This observation has been followed by other groups, which have confirmed the in vitro development of oocyte-like cells in human [74] and other mammalian species, such as monkeys [74], sheep [74], rabbits [74], mice [75], and pigs [76]. Therefore, it may be hypothesized that these small stem cells, similar to VSELs, in OSE may also serve as progenitors of oocytes in vivo in appropriate conditions.
An earlier period saw some theoretical skepticism about these small ovarian stem cells, in vitro development of oocyte-like cells from them, and potential postnatal neo-oogenesis, but no experimental data have been found to counter the possibility [89]. In 2012, White et al. reported in the journal Nature Medicine about the oocyte formation by mitotically active germinal stem cells purified from the ovaries of reproductive-age women, based on DDX4/VASA expression [90]. The morphology, dimensions, and gene expression of oocyte-like cells developed in vitro in this study were quite comparable to those in the studies mentioned above [67,68,88], while the sorted germinal stem cells were slightly larger (5–8 μm). Despite that, the possibility that these populations of ovarian stem cells are related is not excluded.
We suggest that White and coworkers have sorted the immediate descendants of VSELs, based on the size and characteristics of cells. They have also mentioned by themselves that they have found ovarian stem cells equivalent to oogonial and spermatogonial stem cells [91], but unfortunately, they did not refer to VSELs to maybe better elucidate the subject.
The work of White and coworkers has been criticized because some other studies were unable to confirm it in the mouse model and humans [92,93]; therefore, more time is needed to avoid potential methodological obstacles and inconsistency by also respecting the knowledge on VSELs and small stem cells in adult human ovaries.
VSEL-like cells in adult human testicles and their chemoresistance
Interestingly, a population of small stem cells, similar to VSELs, has been found in the testicular tissue of azoospermic patients without sperm [70,94], in histological sections of human testicular tissue [95], and in a culture of hESCs [70]. Moreover, recently it has been demonstrated that chemoablated cells from mouse testicular seminiferous tubules, enriched for VSEL-like cells, undergo spontaneous spermatogenesis in vitro [96]. The cells in seminiferous tubules (mostly VSEL-type cells) that survived the busulfan treatment of the testicles were cultured using the Sertoli cell-conditioned medium containing fetal bovine serum, FSH, and no additional growth factors. It was found that these VSEL-type cells underwent proliferation and clonal expansion in the culture and spontaneously differentiated into sperm-like cells, while the surrounding Sertoli cells attached and provided the needed somatic support. Transcripts specific for various stages of spermatogenesis, such as Gfra (spermatogonia), prohibitin (spermatocytes), Scp3 (meiosis), and protamines (postmeiosis), were confirmed using quantitative reverse transcription PCR. Based on these data, it has been suggested that VSEL-type cells are chemoresistant [96].
Similarly has been observed in adult mouse ovaries. The VSELs have survived chemotherapy and retained their ability to initiate the oocyte-specific differentiation [97]. Moreover, the hormone FSH has exerted a direct stimulatory action on the OSE and induced VSEL proliferation and differentiation into premeiotic germ cell clusters in vitro during culturing of intact chemoablated ovaries [97].
The observation of a similar population of VSEL-type cells in adult ovaries and testicles may suggest the hypothetical persistence of these small stem cells from the embryonic period of life and time of indifferent gonads.
Development of human follicles/oocytes in vivo: involvement of VSELs?
Because in vitro development of oocyte-like cells from ovarian stem cells has already been confirmed, the intriguing question is whether small ovarian stem cells could serve as in vivo oocyte/follicle progenitors in adult human ovaries. In the study of Byskov et al. [98], one of the leading groups researching ovarian physiology, it has been found that all oogonia in human ovaries exclusively stained for SSEA-4, NANOG, and OCT4, while only a small fraction of oogonia stained for MAGE-A4. At birth, only a few oogonia were positively stained for these markers, and they disappeared before the age of 2 years. After that time, the only diplotene oocytes that remained were stained for c-KIT. Interestingly, from the 18th week to 2 years, the medulla contained conglomerates of both healthy and degenerating oogonia and small follicles, waste baskets (WBs), and oogonia enclosed in growing follicles (FWB) [98]. In addition, the medulla of older ovaries contained groups of healthy primordial follicles [98]. This was in conflict with current knowledge on ovarian physiology, which presumes to address the follicles in the ovarian cortex only. This important finding indicated that there might be a population of progenitor cells in the ovarian medulla. From the anatomy, it is known that the ovarian medulla is composed of dense connective tissue and is a place where all the lymphatics, nerves, and vasculature of the ovary are found. We suggest that the progenitors of oocytes/follicles in the medulla might be supplied by blood vessels (Fig. 2). From the literature, it is known that VSELs circulate in the blood stream and may be used where they are needed in the body [7]; therefore, VSELs may also serve as progenitors of oocytes/follicles in the ovarian medulla along with VSEL-like cells in the OSE.

Proposed new model of folliculogenesis/oogenesis in adult mammalian/human ovaries, based on experimental data from literature [7,65
–80,88,90,97
–102]. We suggest that follicles/oocytes in the ovarian cortex develop from small VSEL-like stem cells in the OSE. In addition, the primordial follicles with oocytes can be generated from VSELs in medulla, supplied by the peripheral blood stream. Yellow, VSELs; orange, follicles. Color images available online at
This model could sit in accordance with some previous suggestions that bone marrow-derived cells (BMDCs) accompany origination of PGCs from the OSE stem cells, as observed in normal adult female rats, and from the medullary precursors in adult neonatally estrogenized rats lacking OSE [99]. An alternative origin of PGCs from the medullary region of adult ovaries may explain why ovaries with destroyed or absent OSE are still capable of forming new primordial follicles. In addition, the VSEL-like stem cells in adult ovaries are proposed to possess the FSHR3 receptor and are responsive to FSH, the main reproductive gonadotropin that affects folliculogenesis/oogenesis in mammals [100,101]. Indeed, an effective mobilization of VSELs and HSPCs into the blood stream has been shown in response to ovarian stimulation using FSH to retrieve the oocytes in women in the in vitro fertilization program [102], and the developmental origin of these cells has been suggested.
Gametes After Bone Marrow Transplantation
Experimental data from several studies have demonstrated that in adult bone marrow of both humans and mice reside some progenitor cells, which can develop into female and male germ cell-like cells in vitro and in vivo. Although exciting, these progenitor cells are not well characterized and are still a matter of confusion.
Oocyte-like cells from bone marrow
A good clinical strategy to preserve fertility in young women with blood malignancies (eg, leukemia) is to hormonally stimulate the ovaries, retrieve the oocytes, vitrify, and store them for later in vitro fertilization [103,104]. Although early menopause frequently occurs in young female cancer patients after chemotherapy [105], bone marrow transplantation has been related to an unexplained return of ovarian function and fertility in some cancer survivors [106 –109]. Some studies modeling this process in mice have shown that bone marrow transplantation generates donor-derived oocytes in chemotherapy-treated recipients.
Tilly's research group has demonstrated that bone marrow transplantation can restore oocyte production in wild-type mice, presterilized by chemotherapy, as well as ataxia telangiectasia-mutated gene-deficient mice, which are otherwise infertile and incapable of producing oocytes [110]. Moreover, the donor-derived oocytes have also been observed in female mice after peripheral blood transplantation. The fertilization potential and developmental competence of bone marrow- and peripheral blood-derived oocytes have not been tested, although their morphology, enclosure within follicles, and expression of oocyte-specific markers strongly support the idea these cells had developed toward becoming oocytes. These results identified for the first time that bone marrow as a potential source of female germ cells could sustain oocyte production and fertility in adulthood [110]. In addition, in mice receiving no prior conditioning regimen, the once-monthly infusions of BMDCs retrieved from young adult female donors, marked with a green fluorescent protein, sustained the fertility of aging wild-type females [111].
All this work has been criticized by other researchers [112 –114], but Tilly and colleagues have provided further evidence that immature and nonovulated oocytes can be derived from bone marrow transplantation in a preclinical mouse model of chemotherapy-induced premature ovarian failure (POF) [115] and not from misidentified immune cells, as proposed in others' critiques. Surprisingly, all offspring were derived from the recipient germline and not transplanted one, thus indicating that in the ovaries of recipient mice, there were some oocyte progenitor cells that survived the chemotherapy. We suggest that these cells were VSELs, based on the finding that gonadal VSELs can survive chemotherapy, and are involved in primordial follicle assembly and oogenesis [96,97]. Despite that, more offspring have been retrieved after bone marrow transplantation. Thus, bone marrow transplantation provided some signals for chemoablated ovaries to regenerate themselves by possible involvement of mesenchymal stem cells and improvement of the ovarian niche, as found in other studies [96,116 –119].
Despite such criticism, some other groups have confirmed the above findings. In the mouse model, allogeneic bone marrow transplantation has been examined to determine whether it can improve POF induced by radiation and whether allogeneic ovarian transplantation can modulate the adverse effects of radiotherapy [116]. Transplantation of bone marrow and simultaneous transplantation of ovarian allografts under the kidney capsules have been shown to be efficient in mice, which could be advantageous for young women with POF and osteopenia after chemotherapy and radiotherapy for malignant diseases [116]. In addition, some other studies demonstrated that transplanted bone marrow cells do not directly provide new oocytes but rescue fertility in female mice after treatment with chemotherapeutic agents [117]. The authors have not found any evidence that transplanted bone marrow cells provide new fertilizable oocytes in SCID mice treated with chemotherapeutic agents. However, transplanted bone marrow cells have improved the fertility of SCID mice pretreated with chemotherapeutic agents and positively influenced the ovarian physiology.
Our opinion is that the confusion about the oocytes from bone marrow results from the fact that there are different populations of stem cells that are present in adult mammalian bone marrow (including humans): from pluripotent VSELs [5,6] to classical mesenchymal stem cells [118]. From the literature, it is known that transplantation of mesenchymal stem cells from different origins, including bone marrow, may improve the function of presterilized ovaries in animal models [119]. In different studies, researchers have possibly focused on different populations of bone marrow-derived stem cells; therefore, the general conclusion on germ cell generation from bone marrow-derived stem cells is not possible at present. Further studies are needed with clear characterization of bone marrow-derived stem cells to elucidate the important question regarding the link between germinal and hematopoietic lineages, with special emphasis on bone marrow VSELs.
Sperm-like cells from bone marrow
A small population of BMDCs has been shown to transdifferentiate not only into female germ cells but also into male germ cell-like cells [120,121]. These cells expressed some early germ cell markers, such as Oct4, Figla, Dppa3/Stella, Ddx4/Vasa, and Rnf17, as well as male germ cell-specific markers, such as Dazl, TSPY, Piwil2, Rbm, c-Kit, Tex18, and Stra8. Moreover, it has been evidenced that retinoic acid induces mouse bone marrow-derived CD15, Oct4, and CXCR4-positive stem cells into male germ cell-like cells in a two-dimensional cell culture system [122]. The germ cell-like cells expressed several genes related to male germ cells, such as Mvh, Dazl, Piwil2, Dppa3, and Stra8. These data provided direct evidence that human bone marrow cells can differentiate into putative male germ cells and identify bone marrow as a potential source of male germ cells.
Germ Cell Characteristics of Tumors
Malignant tumors express not only the markers related to pluripotency but often also some embryonic/germ cell traits that are common in malignant tumors and are thought to be involved in malignant tumor behavior. The speculations on why malignant tumors show strong embryonic/germline traits are controversial. It could be a consequence of displaced germ cells or gametogenic programming reactivation.
Recent research demonstrated that a chemical carcinogen, 3-methylcholanthrene (3-MCA), can trigger the germ cell potential of human BMDCs (hBMDCs) [123]. The chemical 3-MCA promoted the generation of germ cell-like cells from induced hBMDCs that had, interestingly, undergone malignant transformation, whereas similar results were not observed in the parallel hBMDCs control cultured without treatment with 3-MCA. The malignant-transformed hBMDCs spontaneously and efficiently developed into oocyte-like cells even at the single-cell level. It was surprising that germ cell-like cells from induced hBMDCs were comparable to natural germ cells in many aspects, including morphology, gene expression, proliferation, migration, further development, and potential of teratocarcinoma formation [123]. The authors concluded that a chemical carcinogen can reactivate the germline phenotypes/properties of human bone marrow (somatic tissue)-derived cells which might provide a novel idea and intervention in tumor biology and therapy.
Some studies indicated that pluripotent cancer-initiating cells existed in chemical carcinogen-transformed mouse BMDCs [124]. These cells were named bone marrow-derived pluripotent cancer-initiating cells and showed properties of embryonal carcinoma cells/ESCs in terms of morphology and gene expression, differentiated into multiple somatic tumor cells, generated oocyte-like cells, and developed into blastocyst-like structures in vitro while causing teratocarcinomas in vivo. These findings demonstrated that pluripotent cancer-initiating cells can originate from BMDCs.
Finally, an early study provided evidence of the presence of PGCs and oogonia in tissue squash preparations, including bone marrow, and sections from hematological malignancies [125]. PGCs were identified by their morphology, intense periodic acid–Schiff (PAS), PAS-D reaction, and presence of calcium-activated neutral proteinase. The bone marrow oogonia were also found to be PAS and PAS-D positive.
Conclusion
This review summarizes several studies, which from different aspects showed the potential link between the germinal and hematopoietic lineage in mammals, including humans. This link may be represented by small stem cells that were discovered in adult human bone marrow, peripheral blood, and umbilical cord blood and were termed VSELs. Very similar populations of VSEL-like cells have also been found in adult human gonads, ovaries, and testicles, thus indicating that these cells may persist in adult reproductive tissues from the stage of indifferent gonads in the embryo. The VSELs from bone marrow and small stem cells, similar to VSELs from gonads, express a specific pattern of pluripotency and several genes related to germinal lineage, especially to PGCs, thus indicating their potential common origin in the epiblast of the embryo. Along with a degree of pluripotency, the VSELs also express some markers of hematopoietic cells; therefore, they may represent a developmental link between the germinal and hematopoietic lineage. The ease with which VSELs spontaneously differentiate into cells of germinal lineage is because they are equivalent to PGCs, whereas it is evidently very difficult to obtain PGCs from ES/iPS cells [90]. Undoubtedly, the VSELs present a great scientific challenge to better understand the development of the human being, some diseases, including complicated cancers, and their potential role in regenerative medicine.
Footnotes
Acknowledgments
The author would like to thank all persons and institutions supporting the ovarian stem cell research of her group, as well as Martin Stimpfel who critically read the article.
Author Disclosure Statement
No competing financial interests exist.