
Abstract
Previous microarray analyses of RNAs from 8-cell (8C) human embryos revealed a lack of cell cycle checkpoints and overexpression of core circadian oscillators and cell cycle drivers relative to pluripotent human stem cells [human embryonic stem cells/induced pluripotent stem (hES/iPS)] and fibroblasts, suggesting growth factor independence during early cleavage stages. To explore this possibility, we queried our combined microarray database for expression of 487 growth factors and receptors. Fifty-one gene elements were overdetected on the 8C arrays relative to hES/iPS cells, including 14 detected at least 80-fold higher, which annotated to multiple pathways: six cytokine family (CSF1R, IL2RG, IL3RA, IL4, IL17B, IL23R), four transforming growth factor beta (TGFB) family (BMP6, BMP15, GDF9, ENG), one fibroblast growth factor (FGF) family [FGF14(FH4)], one epidermal growth factor member (GAB1), plus CD36, and CLEC10A. 8C-specific gene elements were enriched (73%) for reported circadian-controlled genes in mouse tissues. High-level detection of CSF1R, ENG, IL23R, and IL3RA specifically on the 8C arrays suggests the embryo plays an active role in blocking immune rejection and is poised for trophectoderm development; robust detection of NRG1, GAB1, -2, GRB7, and FGF14(FHF4) indicates novel roles in early development in addition to their known roles in later development. Forty-four gene elements were underdetected on the 8C arrays, including 11 at least 80-fold under the pluripotent cells: two cytokines (IFITM1, TNFRSF8), five TGFBs (BMP7, LEFTY1, LEFTY2, TDGF1, TDGF3), two FGFs (FGF2, FGF receptor 1), plus ING5, and WNT6. The microarray detection patterns suggest that hES/iPS cells exhibit suppressed circadian competence, underexpression of early differentiation markers, and more robust expression of generic pluripotency genes, in keeping with an artificial state of continual uncommitted cell division. In contrast, gene expression patterns of the 8C embryo suggest that it is an independent circadian rhythm-competent equivalence group poised to signal its environment, defend against maternal immune rejection, and begin the rapid commitment events of early embryogenesis.
Introduction
C
Cells of the early human embryo, ∼10,000 times larger than somatic cells, are totipotent and appear capable of guiding their cleavage stages without need for external growth factor stimulation, perhaps because cell growth is not needed and key cell cycle checkpoints are not expressed [1,2]. Each cleavage divides the blastomeres into two daughter cells, half the size of the precursor, and without an increase in embryo mass. At the blastocyst stage (∼100 cells), the embryonic cells are about the size of somatic cells and they need to enlarge for each subsequent cell cycle.
Many studies to measure the expression of various growth factors and their receptors, and the influence of growth factor addition to culture systems for early cleaving embryos, have been reported in animal model systems ([3 –6] and [7] for reviews), especially mouse, but only a few studies have focused on early human embryos, most of which rely on reverse transcription–polymerase chain reaction (RT-PCR) amplification of specific mRNAs [8 –10] or immunostaining for proteins [11,12]. Innovative methods of linear amplification of small quantities of mRNA [1,13 –15], improved whole human genome microarrays [16 –18], and RNA deep-sequencing methods for single cells [19] have allowed for more global in-depth analyses of gene expression patterns of preimplantation human embryos.
We have reported that noncryopreserved, normal appearing 8-cell (8C) embryos overexpress circadian oscillators, CLOCK, period, cryptochrome, and ARNTL(BMAL), and cell cycle drivers, Cyclins A, -B, -E and Myc, and underexpress key cell cycle checkpoints, Rb and Wee1 [1,2], relative to pluripotent human embryonic stem (hES) cells, induced pluripotent stem (iPS) cells, and human fibroblasts.
The silence of Rb is in keeping with a lack of growth factor dependence to stimulate early embryo cleavages following fertilization, but the silence of Wee1 heightens the following questions: What cellular controls are in place to ensure accurate DNA replication and chromosome segregation? Is euploid human blastomere cleavage dependent on cyclic overexpression of key proteins rather than on cell cycle checkpoints imposed by growth factor dependency and Gap 2 [20]? The silence of Rb in the human 8C [2], and the human oocyte [14], is in contrast to the mouse, in which Rb is detectable in both oocytes and early cleaving embryos [21], although knockout of Rb family members blocks postimplantation, not preimplantation, mouse embryo development [22].
Perhaps the need for gene amplification to support timely signals to the mother to prevent miscarriage outweighs the need to maintain accurate ploidy during preimplantation development [20], or perhaps heretofore unrecognized intracellular signals, such as the circadian oscillators, are guiding early embryonic cleavages. Growth factors and cytokines expressed and secreted by the embryo itself, perhaps influenced by circadian transcription factors, which would limit their availability to certain time periods, could exert autocrine control on embryo development and paracrine stimulation of endometrial receptivity [4 –6,8].
The goal of the present study is to begin to understand the repertoire of growth factors and their receptors expressed in normal 8C human embryos not previously cryopreserved. We have compared microarray data from the 8C embryos, hES cells, and human fibroblasts before and after induced pluripotency (iPS cells) to determine differences between the totipotent embryo cells, the pluripotent hES and iPS cells, and the lineage-restricted fibroblasts. The results provide clues about the unique molecular mechanisms that guide early human development and participate in the complex embryo–endometrium dialog. We analyzed two pools of five embryos each to avoid individual embryo and blastomere bias in an effort to focus on overarching gene expression and pathway differences between the totipotent 8C embryos and the pluripotent stem cells.
Using public databases, we compiled a list of 487 growth factors and receptors and grouped them according to Table 1, with underlining to indicate the elements that are expressed with a circadian rhythm in mouse tissues according to CircaDB [23]. The epidermal growth factor (EGF) family consists of several structurally homologous polypeptides that act by binding to the EGF receptor (EGFR) or the erbB family of receptors (ERRB2–4) [24 –26]. The fibroblast growth factor (FGF) family, known to play numerous essential roles in development and cell proliferation, consists of about 23 structurally related proteins, 18 of which act by binding to FGF receptors (FGFRs), and four of which, FGF homologous factors, FGF11–14(FHF1–4), act intracellularly [27,28].
Underline denotes gene elements reported to be circadianly expressed in some mouse tissues (
The insulin and insulin-like growth factor (IGF) family consists of insulin (INS) and two IGF peptides (IGF1 and IGF2) that share structural similarity with insulin, plus a group of six high-affinity binding proteins (IGFBP1–6), IGFBP proteases, and three receptors, insulin receptor (INSR) and IGF1R and IGF2R.
The transforming growth factor beta (TGFB) superfamily includes at least 38 structurally related proteins that share sequence homology with TGFB, such as activins, anti-Mullerian hormone, bone morphogenetic proteins (BMPs), endoglin, growth differentiation factors (GDFs), left–right determination factors (LEFTY1, -2), Nodal, and teratocarcinoma-derived growth factors (TDGF) [29]. Two families of TGFB receptors have been described, type I receptors (TGFBR1) and type II receptors (TGFBR2), with counterparts for other family members, such as activin receptor type I and type II and BMP receptor type I and type II, as well as Endoglin (ENG), an auxiliary receptor that modulates TGFB signaling.
Nine different genes (PDGF1–4 and VEGF1–5) encode platelet-derived growth factors and vascular endothelial growth factors, respectively, which form hetero- or homodimers, binding to their receptors (PDGFRA–D) and (FLT1–4). There are four members of the nerve growth factor family, nerve growth factor (NGF), neurotrophins, neuron-derived neurotrophic factor, and brain-derived neurotrophic factor (BDNF), which act mainly through their respective receptors. The cytokines are a large family of interleukins, interferons, inducible factors, and regulatory factors, including colony-stimulating factors (CSFs) and leukemia inhibitory factor (LIF), tumor necrosis factor (TNF), and FAS ligand, thought to act principally through their cognate receptors.
The CCN family consists of six members: CYR61, connective tissue growth factor (
For purposes of discussion, the microarray results are grouped according to the level of detection in each cell type and growth factor superfamily, subjected to GenBank descriptions and pathway analyses using DAVID and GeneMANIA, and queried for reported circadian expression according to CircaDB [23]. Gene elements reported to be expressed in a circadian pattern in mouse tissues are underlined throughout this report.
Materials and Methods
Supernumerary embryos were donated by nine Greek couples undergoing assisted reproduction at the 1st Obstetrics and Gynecology Department of the University of Athens, “Alexandra” Maternity Hospital, Athens, Greece, as reported [1,2]. The study protocol and written informed consent process were reviewed and approved by the ethics research committees of “Alexandra” Hospital and Bedford Research Foundation, as described (Supplementary Data; Supplementary Data are available online at
Using gene ontology (GO) terms (
Array signals ranged from 20 fluorescence units (FUs) to >740,000 FUs. For purposes of discussion, we set 500 FUs as the upper limit for off/marginal expression, 500–5,000 FUs as the range for moderate expression, and greater than 5,000 as the threshold for high expression. The highest signal was used to group genes with multiple probes. We chose a conservative sevenfold difference in detection levels to designate over- or underdetected in the tables and discussion. This is two standard deviations from the mean of the variation between the microarray elements on the two 8C embryo arrays [1,2]. This approach does not distinguish between gene variants and is not meant to be a comprehensive analysis; the raw data are presented in Supplementary Table S1 for use in other analyses.
Microarray detection levels were further evaluated by real-time PCR analyses of eight additional 8C embryos whose RNA was individually extracted and analyzed for selected mRNAs (Supplementary Data), the relative copy numbers for which agreed in all instances with the microarray results.
Groups of gene elements were further analyzed for common pathways by GeneMania (
Results
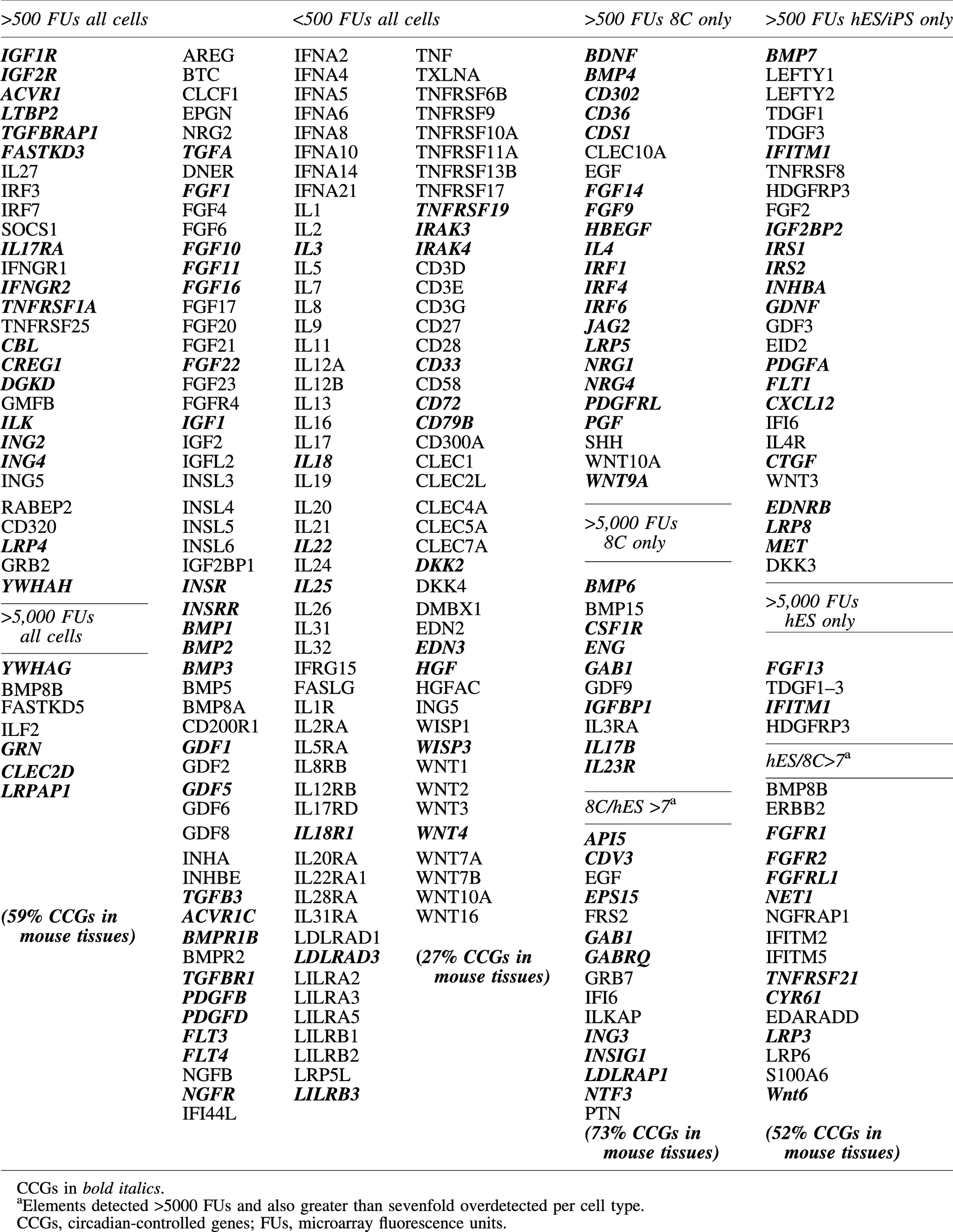
The list of 487 growth factors and receptors (Table 1) identified 1,044 gene elements in 8CFES, listed in Supplementary Table S1. Two hundred and twenty (21%) of the gene elements were off/marginal in all the cells in 8CFES (Table 2, Supplementary Table S1), with only 34 (3.2%) gene elements detected above off/marginal on all HCFES arrays (Tables 3 and 4). A comparison of the percentages of gene elements in each category is listed in Supplementary Table S2.
Bold italics denotes gene elements detected less than 7-fold lower than hES cells; bold italics underlined less than 70-fold lower than hES cells.
FUs, fluorescence units from Supplementary Table S1.
8C, 8-cell; GF, growth factor; hES, human embryonic stem; iPS, induced pluripotent stem; Rec, receptor.
Bold denotes gene elements detected greater than 7-fold higher than hES cells; bold underlined greater than 70-fold higher than hES cells; bold italics denotes gene elements detected less than 7-fold lower than hES cells; bold italics underlined less than 70-fold lower than hES cells.
FUs, fluorescence units from Supplementary Table S1.
Bold denotes gene elements detected greater than 7-fold higher than hES cells; bold underlined greater than 70-fold higher than hES cells; bold italics denotes gene elements detected less than 7-fold lower than hES cells.
FUs, fluorescence units from Supplementary Table S1.
The lineage-restricted fibroblasts have more off/marginal (43%) and more high, (12%), whereas the totipotent 8C embryos have fewer off/marginal (30%) and fewer high (5%). Fifty-one gene elements were overdetected on the 8C arrays, at least sevenfold higher than the pluripotent cells, and 44 were underdetected, at least sevenfold lower (Tables 3 and 4). These groupings are not meant to indicate the level of protein expression, nor cellular importance, but as a starting point for discussion of which growth factor/receptor pathways might function in each cell type for autocrine stimulation and/or for paracrine stimulation between cell types frequently cocultured, such as embryonic stem cells with a fibroblast feeder layer, and iPS cells derived in a background of fibroblasts. Except as noted in a special section at the end of Results, iPS cells more closely reflected the microarray detection patterns of hES cells than their fibroblast precursors.
EGF family
Several EGF family members were silent or marginal on all 8CFES arrays, including
FGF family
Eleven FGFs and FGFR4 were silent or marginal on all 8CFES arrays (Table 2). Six (
INS/IGF family
INS,
TGFB family
Many members of the TGFB superfamily were off/marginal on the arrays of all cells, including several BMPs, several GDFs, inhibin (
PDGF/VEGF family
Two PDGFs (
NGF family
Nerve growth factor (NGFB) and nerve growth factor receptor (
Cytokines
Many members of the cytokine superfamily were off/marginal on all the arrays, including the interleukin-1 family (IL1A, -1B, IL1R1, -R2), interleukin-3 (
Ten members of this family were at least sevenfold overdetected on the 8C arrays relative to hES/iPS cells: interferon inducible factor 6 (IFI6), three interferon regulatory factors (
CNN family
No WISPs were above off/marginal on any 8CFES arrays, except WISP2, which was detected on the fibroblast arrays.
Others
Many factors and receptors grouped in this category were not above off/marginal on any 8CFES arrays, including several members of the WNT and Dikkkopf pathways (Table 2, 6 and Supplementary Table S1). Twenty-one members were detected on all 8CFES arrays (Tables 3, 4, and 6):
iPS cells and hES cells
As has been previously reported, the iPS cells generated by Yamanaka and colleagues [31] from fibroblasts exhibit microarray results similar to the hES cells. Interesting exceptions noted are CD4 remained 13-fold lower in iPS cells than hES cells and 20-fold lower than 8Cs; activin receptor, ACVR1B, was 7-fold lower than hES cells and 19-fold lower than 8Cs; LEFTY2 was 12-fold lower in the iPS cells than hES cells, but not lower than 8Cs; IFITM5 was 9-fold lower than hES cells, but not lower than 8Cs; FGF3 was 7-fold lower than hES cells and 3-fold lower than 8Cs. The only gene not downregulated in the iPS cells was S100A6 (a high value on all the arrays at >100,000 FUs), 9-fold higher than the hES cells and 88-fold higher than the 8Cs. Interestingly, NODAL appeared only partially upregulated on the iPS arrays.
Pathway analyses
To query the gene sets for cell pathway enrichment, we submitted gene groups to GeneMania [32] and to DAVID [33]. The 34 gene elements detected on all 8CFES arrays (Tables 3, 4 and 6) revealed no particular pathway enrichments and the enzyme-linked receptor protein signaling pathway (CBL, ACVR1, DGKD, GRB2, IGF1R, LTBP2, TGFBRAP1) was the top functional annotation clustering category in DAVID.
Half of the 51 genes overdetected specifically on the 8C arrays were DAVID functional annotation clustering growth factor and signal, with subcategories enriched for various developmental processes (Table 5) because of detection of BMP4, -6, SHH, EGF, and ENG along with developmental-specific factors such as BDNF and PGF. GeneMANIA analysis of the same set emphasized leukocyte differentiation, (SHH, BMP4, IL23R, CSF1R, GAB2,
GO, gene ontology.
Over half of the 44 gene elements underdetected on the 8C arrays relative to hES cells were DAVID functional annotation clustering signal/secreted (Table 5), 13 were growth factor activity, 12 were extracellular space, 7 were positive regulation of cell motion, 16 were regulation of cell proliferation (Table 5), and 7 were blood vessel morphogenesis. The top functions enriched in GeneMANIA were regulation of ERK1/2 cascade (CYR61, KDR, FGFR2, FGF2, ERBB2, PDGFA), morphogenesis of an epithelium (CYR61, WNT6, FGFR2, FGF2, GDNF, TDGF1, LRP6, MET, PDGFA), phosphatidylinosital-mediated signaling (IRS1, -2; ERBB2; FGFR1; PDGFA; FLT1), and tube development (MET, GDNF, LRP6, WNT6, BMP7, FGF2, FGF2R, PDGFA).
Circadian-controlled genes
Gene element groups were submitted to CircaDB for assignment to reported circadian expression in some mouse tissues (Table 6). Of the 34 gene elements above off/marginal on all cell arrays, 59% were reported to exhibit circadian patterns of expression, peaking every 24 h. In contrast, only 27% of the 150 gene elements below off/marginal on all cell arrays were reported to display circadian expression. Of the 48 gene elements specific to the 8C arrays, 73% exhibited circadian expression in mouse tissues in CircaDB, whereas of the 47 gene elements specific to the hES/iPS cells, only 51% exhibited circadian expression (Table 6).
Discussion
The capacity for hES/iPS cells to maintain uniform pluripotency during expansion for many generations in culture is a remarkable artificial state that shows great promise for stem cell therapies. Suppressing commitment, while encouraging expansion and maintaining normal karyotype and commitment potential, was an early challenge of hES/iPS cell culture that has now largely been met. In contrast, embryonic development is a dynamic process of commitment, expansion of committed cells, and patterned differentiation.
The 8C stage of human embryo development is a fleeting period of totipotency for what is essentially a developmental equivalence group that will undergo its first commitment event to trophoblast and inner cell mass within the next couple of cell cycles. In contrast to hES/iPS cell culture, the goal of human embryo culture is to fully support natural commitment events to the blastocyst stage in an in vitro environment.
Heuristically, it seems most likely that the early embryo is in charge of its development within a hospitable maternal environment. For the first few weeks, the embryo must signal the mother that it is developing to avoid a miscarriage. Whether or not its early in vivo development is augmented by specific maternal cues has thus far not been determined, but since most in vitro fertilization (IVF) conceptions take place in the absence of growth factors, maternal signals during the first few days do not appear to be essential for successful pregnancy. The high variability of successful in vitro embryo development among mouse strains [34 –36] as well as human couples supports the concept, however, at least some fertilized human eggs might benefit from as yet undetermined growth factors while in culture.
One advantage of in vitro culture is that the early embryo avoids the threat of immune rejection by the maternal reproductive tract, but that possibility is presented abruptly at the time of embryo transfer. Whether or not further embryonic development would be enhanced if the maternal tract were preconditioned to receive the embryo, and/or if the embryo were preconditioned to the maternal tract, is urgently needed information for programs of assisted reproduction.
The concept of an equivalence group implies that each of the 8C stage blastomeres is equivalent to the others and poised for the next developmental event, but this has not been demonstrated, even for mouse embryos. Attempts to derive stem cell lines from individual 8C blastomeres from the mouse, cow, and human essentially fail, in that only a small percentage of the blastomeres continue to divide when separated from sisters [37 –39]. This suggests either that paracrine signaling is essential for further cell division at this stage or that aneuploidy incompatible with independent continued cleavage is common in blastomeres in early embryos, as has been suggested [20,40], or both.
Our present comparison of growth factor/receptor gene expression in 8C embryos with pluripotent hES/iPS cells and committed fibroblasts has revealed insight into potentially active, and inactive, growth factor pathways in early human embryos derived in vitro as well as which pathways are artificially balanced differently in cultured hES/iPS cells to stabilize robust multiplication and pluripotency.
EGF family
More members of the EGF family were detected on the 8C arrays than on the other arrays and at markedly higher levels, including EGF and
The robust expression on the 8C arrays of the EGFR adaptor protein, GRB, and its cognate docking proteins,
This suggests that early cleaving embryos have the capacity to carry out important downstream cellular functions without the growth factor/receptor interaction required by somatic cells. This type of innate pathway potential may serve to augment their independence during the preimplantation period. The robust detection of the signaling molecule, neuregulin (
FGF family
Detection of three of the four FGFRs suggests 8C embryos and hES/iPS cells may be receptive to multiple members of the FGF superfamily, although receptor detection on the 8C arrays was low. The lack of detection of FGF4 (essential for blastocyst formation in the mouse) and FGF8 (essential for gastrulation in the mouse) on any of the 8CFES arrays is surprising and suggests that they do not play similar roles in human embryo development and are suppressed in the cultured hES/iPS cells to maintain pluripotency. The lack of FGF2 detection on the 8C arrays is consistent with the lack of FGF2 detection in mouse embryos (
Taken together, the data suggest that the pluripotent cells and the fibroblasts are capable of autocrine FGF2 and FGF3 signaling, but the 8Cs only autocrine FGF3 signaling.
The differential expression of
This suggests that the robust expression of FGF14(FHF4) in the 8Cs relates to a novel role for this protein in early embryo sodium channel regulation and/or demonstrates how poised the embryo is to begin neuronal differentiation. Moreover, the stimulation of the FGF14(FHF4) sodium channel complex by GSK3 [53] is especially intriguing given that inhibition of GSK3 inhibits spontaneous differentiation in pluripotent stem cells [54,55].
The detection of API5 in all cells, and at highest levels on the 8C arrays, suggests that mechanisms to block apoptosis are important to early development as well as to cells in culture.
INS/IGF family
Insulin is a common component of cell culture additives (eg, ITS, insulin-transferrin-selenium), although INSR was essentially silent on all 8CFES arrays, suggesting that canonical insulin signaling is lacking in these cell types. Nonetheless, the detection of
The marked overdetection of IGFBP1 on the 8C arrays suggests the possibility of a unique function in early embryo development in addition to the reported importance of IGFBPs to fetal development and the fetal/maternal interface in the placenta [57,58]. IGFBP1 is best known for its role in binding, thereby controlling the activity of
The
TGFB family
The pattern of TGFB1 and -2 and TGFB1 and -2 receptor detection suggests the possibility of autocrine stimulation of TGFB pathways in fibroblasts, but not hES/iPS cells, nor the 8C.
BMPs are important molecules in tissue differentiation, especially modulation of a variety of endocrine systems [61]. BMP8B, the only BMP family member detected on all 8CFES arrays, although at lowest levels in the 8Cs, is known to play a role not only in developing skeletal tissues [62] but also in the induction of primordial germ cells in the mouse [63].
Downregulation in the hES/iPS cells of the maternal messages overdetected in the 8Cs (
BMP15 and GDF9 are well-characterized maternal messages in several species [66] whose synergism is essential for normal fertility. Using subtractive suppression hybridization, BMP15 was reported to decrease from oocyte to the 8C stage in mice [67], but by global gene expression analysis, BMP15 increased from the 4-cell to 8-cell stage [68] and, by RT-PCR, was found to persist through the 8C stage in bovine embryos [69]. The recent finding that BMP15 is overexpressed in proliferative leukemia stem cells supports a role in growth factor-independent cell division [70]. BMP15/GDF9 heterodimers are reportedly more bioactive than homodimers [71,72] and their overdetection on the 8C arrays suggests they may function during early embryonic development in addition to regulating ovarian function.
BMP7, overdetected on hES/iPS arrays, has been reported to regulate neural progenitor cells during brain development in the mouse [73].
The detection of
In contrast, although ActivinIIB receptor was robustly detected on the 8C arrays, NODAL was only moderately detected and ACTIVIN and TDGF1(CRIPTO) were low/marginal, indicating that Activin/Nodal signaling is not functional at the 8C stage. Taken together, the data suggest that Lefty1, -2 are important pluripotency factors to suppress Activin/Nodal-stimulated A-P patterning and mesoderm/endoderm formation in cultured pluripotent stem cells, but that the totipotency of 8C embryos may be due, in part, to the absence of Activin/Nodal signaling, a possibility supported by the relative silence of LEFTY1, -2 on the 8C arrays.
The robust detection of
Also known as macrophage inhibitory cytokine-1, GDF15, most robustly detected on the 8C and fibroblast arrays, has been implicated in several pathways, including immunomodulation. It is tempting to speculate it plays a role in blocking maternal immune rejection of the newly developing embryo.
The high level of detection of GDF9 (Supplementary Table S1 and Table 4) specifically on the 8C arrays is not surprising given its known role as a maternal message important to folliculogenesis, but it has more recently been shown to be an effective antiapoptotic signal by inhibiting caspase 3, suggesting a survival factor role in early embryo development as well as a key player, along with ASF1A and OCT4, in nuclear reprogramming [80]. The silence of GDF3 on the 8C arrays, but not the hES/iPS arrays, indicates that although it has a well-described role in maintaining pluripotency in hES cells [81], it is not operational at the totipotent 8C stage.
VEGF/PDGF family
The detection of VEGFC and ECGF1(TYMP) involved in angiogenesis and induced by gonadotropins [82] suggests that the 8C is poised to initiate angiogenesis, and the pathway is silenced in the hES/iPS cells to maintain pluripotency. The exact function of PDGFRL, overdetected more than sevenfold on the 8C arrays, is unknown, although its identification as a tumor suppressor gene suggests it may assist the fidelity of blastomere cleavage in the absence of canonical cell cycle checkpoints.
NGF family
The overdetection of
Cytokines
Several prior studies have examined the expression of the IL1 and LIF families in early embryos because they are believed to play important roles in implantation [83 –85] with mixed results. The absence of a signal on the multiple probes for the IL1 family and the low/marginal detection of the Lif family on the 8C microarray in the present study suggest that they are not important pathways at the 8C stage of human embryos.
The high level of detection of ILF2(NF45) and ILF3(NF90) on the arrays of all the cells, with the 8C arrays being the highest, indicates that the alternate system of translational control, recruitment of the ribosome to the protein initiation site without the canonical 5′terminal 7-methylguanosine cap, is operational in all cell types, including the 8C embryo [86]. The system is thought to allow the cell to respond quickly to a changing environment, including upregulation of members of the inhibitor of apoptosis family in keeping with results discussed above. In addition to interleukin transcription, ILF2(NF45) may play a role in regulation of CyclinE and Survivin, previously reported to be overdetected in 8C embryos [1,2].
Although CSF1 was detected on the 8C arrays at the off/marginal level, an alternate ligand for CSF1R, IL34(C16orf77), was detected at the low/moderate level on the 8C arrays, both of which were silent on the arrays of the other cells. This suggests the possibility of autocrine signaling through the CSF1R, a pathway reported to be responsible for cell proliferation, as well as trophoblast development [87]. CSF1 and CSF1R proteins were previously reported in human embryos later than the 8C stage.
The marked overdetection of
Others
Many factors and receptors listed in this category are known members of cell differentiation and control pathways and were silent on all the arrays. Nine were overdetected on the 8C arrays, two of which, CLEC10A and CD36, were 70-fold overdetected. CLEC10A(MGL) is not well understood, but is known to be immunosuppressive [88], and CD36 is a widely expressed cell surface glycoprotein involved in a number of cell processes, including cell–cell interactions.
Circadian-controlled genes
An estimated 10% of mouse gene elements are reported to be circadian-controlled genes (CCGs) [23]. The much higher percentage of the growth factor and receptor gene groups reported here indicates circadian controls may be more focused on growth factor-responsive pathway genes than on the genome at large. This raises the provocative possibility of a circadian fine-tuning of growth factor/receptor pathway responses. Limiting the availability of growth factors and/or their receptors to defined times of day would support a more programmed, patterned cellular response, perhaps especially important during embryonic development.
The fact that the gene elements specific to the 8Cs had the highest percentage of reported CCGs (73%) is in agreement with our earlier report of enhanced expression of circadian oscillators in the 8C human embryos. The possibility of a circadian influence during early development is an intriguing concept in urgent need of additional study.
Conclusions
EGF family
8C human embryos may influence their environment by EGF and
FGF family
The 8C embryo may be capable of FGF3/FGFR3 autocrine signaling, and the ion channel regulator, FGF14(FH4), may have a novel role in early human embryos.
INS family
The detection of
TGFB family
The relative silence of LEFTY1 and -2 in the 8C embryos was surprising given their prominence as pluripotency markers in pluripotent stem cells and may be due to lack of Activin/Nodal/TDGF1 signaling. The overdetection of
VEGF/PDGF family
The detection of ECGF1(TYMP) and VEGFC only on the 8C and fibroblast arrays suggests that the 8C embryos are poised for angiogenesis.
NGF family
Cytokines
LIF appears to not be an important signaling pathway at the 8C stage, despite the well-documented importance of LIF signaling during blastocyst formation and implantation for both mouse and human embryos [89]. CSF1R overexpression could result in autocrine signaling by IL34, an alternate ligand, or could participate in paracrine signaling by CSF1 expressed by the maternal reproductive tract.
Others
The marked overdetection of CLEC10A and CD36 specifically on the 8C arrays suggests novel roles in early human development not previously reported.
Circadian-controlled genes
The variability of the repertoire of CCGs in different mouse tissues emphasizes the need for caution in extrapolating those data to human embryos and cultured stem cells. Nonetheless, given the potential importance of circadian signals to early embryo and stem cell development, this is an area in urgent need of additional study. The ambiguous and artificial state of pluripotent stem cells needs to be considered in all studies designed for them to serve as models for early human developmental pathways.
Footnotes
Acknowledgments
This work was supported by the Naylor Research fund, the Irene Solonsky Gift fund, and the National and Kapodistian University of Athens research fund. The authors are grateful to Deborah Weidman and Elizabeth Creason for assistance with CircaDB.
Author Disclosure Statement
All authors have no commercial association and no competing financial interest with the work presented here.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.