
Abstract
The negative effects of space flight on cardiovascular health of astronauts have been demonstrated and documented over many years. Endothelial cells (ECs) play an important role in regulating weightlessness-induced cardiovascular dysfunction. This project seeks to study the effect of microgravity on the differentiation of endothelial progenitor cells (EPCs) into ECs and the downstream functions of the differentiated cells. Initial exposure of EPCs to microgravity indicated an inhibition of migratory, proliferative, and antithrombogenic capacity of the differentiated ECs. However, our results indicate a potential recovery of proliferative and antithrombogenic functions on prolonged exposure to microgravity.
Introduction
U
EC dysfunction has been implicated as an important factor in weightlessness-induced cardiovascular deconditioning [5]. ECs are sensitive to physiological mechanical cues, such as altered gravity conditions, which cause them to undergo significant changes in morphology and function [5 –8]. In addition to resident ECs, progenitor cells recruited from the bone marrow into circulation are also involved in maintaining vascular homeostasis [9,10]. While great attention has been placed on understanding the effects of weightlessness on the endothelium, much less work has been done to study the effect of microgravity (μg) on the “repair cells,” circulating endothelial progenitor cells (EPCs). The goal of this study is to elucidate the effect of microgravity exposure before EPC differentiation down an EC lineage and the subsequent effects on the function of the resulting ECs.
As a first step toward understanding the effect of gravitational unloading on astronauts, we modeled these mechanisms in porcine cells using a NASA-designed rotating wall vessel (RWV) bioreactor that simulates microgravity on earth (Synthecon, Houston, TX). The RWV bioreactor consists of a cylindrical vessel that maintains cells in suspension by slow rotation around its horizontal axis with a coaxial tubular silicon membrane for oxygenation [6,11]. The porcine model was chosen in this study since it is widely preferred as a model system for vascular research due to its physiological and hemodynamic similarities to the human vascular system [12]. In addition, a reproducible and consistent method to induce the differentiation of porcine blood-derived EPCs has been reported [13].
The effect of altered gravity culture conditions on the function of mature porcine ECs has been studied in the past [14]. However, to our knowledge, the effects of short-term hypogravity or microgravity exposure on circulating progenitor cells have not been reported before. This study describes the effect of simulated microgravity on the differentiation of circulating progenitor cells derived from pig blood along an endothelial pathway. This study also describes the effect of short-term exposure to simulated microgravity, on the downstream functionality of the differentiated ECs such as release of soluble factors, prostaglandin E2 (PGE2) and nitric oxide (NO), proliferation, migration, and ability to regulate clotting mechanisms.
Materials and Methods
Isolation of peripheral blood mononuclear cells from porcine blood
About 50 mL of circulating peripheral blood was collected from the exterior jugular vein of domestic pigs (Department of Animal Sciences, University of Florida) in the presence of acid citrate dextrose anticoagulant. All blood collection procedures were performed in accordance with the rules and regulations of the Institutional Animal Care and Use committee at the University of Florida. Within 4 h of blood collection, the peripheral blood mononuclear cells (PBMNCs) were isolated by density gradient centrifugation as previously described [13,15]. An average of 140–160 million mononuclear cells obtained from the isolation process were suspended in endothelial cell growth media 2 (EGM-2; Lonza, Walkersville, MD), supplemented with 20% fetal bovine serum. Porcine aortic endothelial cells (PAEC; Cell Applications, San Diego, CA) between passages 4 and 6 cultured in the EGM-2 serve as a positive control for mature EC function and response.
Rotating wall vessel bioreactor system
The culture of cell systems in actual microgravity can be achieved only through parabolic flights, rockets, space crafts, or laboratories available on the International Space Station. However, the ability to culture various cell systems in real microgravity is limited by the cost involved in performing these experiments on space missions. Therefore, a variety of devices have been designed to mimic some aspects of real microgravity on earth [11]. In this study, some aspects of simulated microgravity (μg) conditions were established using a RWV bioreactor consisting of a rotor base and detachable cell culture chambers (Fig. 1A). The chamber consists of a silicone membrane for gas exchange as shown in Fig. 1B. Using this setup, the time-averaged gravitational vector acting on these cellular assemblies is reduced to about 10−2 g. For simplicity, this condition is referred to as simulated microgravity. The validity of the RWV model to simulate weightlessness has been verified and reported extensively in literature [11,16,17]. In addition, the use of RWV to simulate aspects of microgravity has been confirmed and validated by experiments in real microgravity in the past [7,18 –20].

Microgravity culture of PBMNCs
The bioreactor setup as described above was placed in a cell culture incubator maintained at 37°C and 5% carbon dioxide. Our experimental setup included (1) PBMNCs suspended (18 × 106 cells in 50 mL of EGM2) inside the bioreactor chamber for microgravity exposure and (2) PBMNCs seeded at a density of 2–3 × 105 cells/cm2 on fibronectin-coated tissue culture polystyrene (TCP) multiwell plates (BD Biosciences, Franklin Lakes, NJ) to be cultured statically with no micrograviy exposure. To maintain conditions of microgravity, the bioreactor chambers containing PBMNCs were rotated at a minimum speed of 5 rpm. Following 6, 12, or 18 h μg exposure, the PBMNCs were collected, then seeded on fibronectin-coated TCP plates, and cultured statically for 7–10 days.
Differentiation, colony formation, and immunocytochemistry
The statically cultured cells, both those preexposed to microgravity and those in normal gravity for the entirety, were visually assessed to observe colony formation every day for 8 days using an inverted phase-contrast microscope (Nikon TE2000U, Melville, NY). The first appearance of densely packed, isolated, cobblestone morphology, endothelial-like cells in a given field of view was recorded as indicative of EC differentiation. The number of colonies and the rate of new colony formation were recorded.
Porcine endothelial (PE) like cells were examined using immunohistochemical analysis to confirm EC phenotype. Briefly, cells were fixed in 4% paraformaldehyde, permeabilized, if required, using 0.2% Tween-20, and then blocked using 3% bovine serum albumin. Following which the cells were incubated overnight at 4°C with a primary antibody (diluted 1:100) against endothelial nitric oxide synthase (eNOS; Abcam, Cambridge, United Kingdom) and CD31 (diluted 1:100; AbDSerotec, Raleigh, NC). Subsequently, the probed cells were incubated with a fluorescein isothiocyanate tagged isotype-matched secondary antibody (Santa Cruz Biotechnology, Santa Cruz, CA) and nuclei were stained using Hoechst 33342 (ThermoScientific, Waltham, MA). Fluorescent images were captured using an inverted fluorescent microscope (Nikon TE2000U).
Endothelial-like cell migration and proliferation
Cell migration was assessed using the established scratch assay [21]. Briefly, PE-like cells differentiated after 6, 12, and 18 h of preexposure to simulated microgravity as well as those cells differentiated in normal gravity, were cultured to ∼90% confluence on multiwell TCP plates in the culture media as described above. A linear scratch was made through the monolayer using a pipette tip to create a “cleared” area, then the cells were incubated at 37°C for 18 h to allow for migration. Digital images of the scratch surface were taken before and after incubation and the percentage area covered by the migrating cells was quantified using ImageJ software version 1.46.
Cell proliferation was assessed by changes in DNA concentration from cultured cells over time and quantified using the Quant-it picogreen dsDNA assay kit (ThermoScientific). Briefly, PE-like cells differentiated after 6, 12, and 18 h of preexposure to microgravity as well as normal gravity-cultured controls, were seeded on 12-well TCP plates at a seeding density of 2,500 cells/cm2. At 24-h increments over 4 days, the cells were lysed to collect and quantify the intracellular DNA. The concentration of DNA contained within the lysates was normalized by subtracting the initial seeded DNA concentration from the concentration at each time point for each sample.
NO and prostaglandin secretion
NO and prostaglandin E2 secretion was evaluated in PE-like cells differentiated after 6, 12, and 18 h of preexposure to simulated microgravity as well as the cells differentiated under normal gravity. The amount of total NO and PGE2 secreted by the cells into the culture medium over a period of 24 h for each cell population was determined using the total NO assay kit and Prostaglandin E2 Assay kit (R&D systems, Minneapolis, MN) following manufacturer protocols. Briefly, the concentration of NO measured is based on the enzymatic conversion of nitrates to nitrites and the detection of total nitrite concentration by the Greiss reaction. PGE2 concentration was assessed by enzyme-linked immunosorbent assay. The concentration of both NO and PGE2 was normalized to cell number and expressed in terms of concentration per cell.
Clotting kinetics
Following previously reported methods, the kinetics of both plasma and whole blood clot formation was assessed [13,15]. Briefly, plasma and whole blood were exposed to monolayers of PE-like cells differentiated after 6, 12, and 18 h of preexposure to simulated microgravity as well as cells differentiated under normal gravity, and the kinetics of clot formation were reported. In this study, PAEC served as a positive control for antithrombogenic EC function, and bare TCP serves as a negative control representing the kinetics of uninhibited plasma and blood clotting.
Statistical analysis
All data are reported as means ± standard deviations (SD). Numerical means were compared using a student two-sample t-test. Analysis of variance (ANOVA) with Newman–Keuls multiple comparison test post hoc analysis was employed to identify significant differences between three or more means. P values <0.05 were reported as significant.
Results and Discussion
This study reports the effect of early exposure to simulated microgravity on the differentiation and downstream function of porcine blood-derived progenitor cells. Although multiple studies have investigated and reported the effect of altered gravity conditions on the behavior of adult ECs, there are limited reports on its effect on circulating progenitor cells [5]. Since these cells have been shown to be involved in vascular repair mechanisms in the body, it is important to understand the functionality of these cells when exposed to short-term microgravity, to better understand effects of cardiovascular deconditioning in space. Previous studies have reported significant alterations to morphological characteristics and changes to gene and protein expression in mature ECs within a few minutes to a few days of microgravity exposure [5,22 –24]. Moreover, the changes in the expression of immunological cytokines in PBMNCs and recovery of their functions on short-term exposure to simulated microgravity ranging from within a few hours to up to 3 days have been reported before [25].
In this study, circulating progenitor cells contained within the PBMNC fraction of porcine whole blood were exposed to simulated microgravity, using an RWV bioreactor designed originally by NASA to simulate the reduction in earth's gravity (1g) to microscale (μg) on cultured cells. The time-averaged gravitational vector acting on these cells is reduced to about 10−2 g using this setup. After exposing the circulating progenitor cells to 6, 12, and 18 h of microgravity, the cells were then cultured on fibronectin-coated TCP under static conditions and their ability to differentiate into ECs was assessed by the formation of colonies with EC-specific “cobble-stone” morphology (Fig. 2A). In all cases, the observation for colonies was concluded after 8 days after which it was difficult to discern discrete isolated colonies. Cells exposed to microgravity for longer than 18 h showed significantly reduced or no colony formation when cultured on fibronectin-coated TCP. This indicates the existence of a possible threshold time of microgravity exposure beyond which the differentiation ability of these PBMNCs may be permanently impaired.

On assessing the kinetics of EC colony formation, it was observed that the average rate of EC colony formation decreased with increased time of exposure to microgravity between 3 and 6 days of culture (Fig. 2B), while the normal gravity showed the highest rate of colony formation as indicated by an average of 10 colonies per day. The control population of PBMNC, cultured under normal gravity conditions formed colonies at the earliest time period of 3 days compared to microgravity-exposed populations, which did not form any colonies as early as the third day of culture. The endothelial phenotype of both the microgravity-exposed and normal-gravity cultured cells was confirmed by their characteristic cobble stone morphology (Fig. 2C) and positive staining for eNOS (Fig. 2D, E) and CD31 (Fig. 2F, G). In the past, the decrease/delay in colony formation of ECs from EPCs has been associated to preexisting cardiovascular complications [26,27]. This delay in differentiation indicates a possible detrimental effect of simulated microgravity on the onset of differentiation of EPCs to ECs that may affect the regulation of cardiovascular health by these cells. In addition, the ability of simulated microgravity to inhibit bone marrow hematopoetic stem cell differentiation has been confirmed by Plett et al. [28].
Furthermore, we investigated the effect of this altered condition on the downstream functional abilities of the differentiated endothelial-like cells that were preexposed to microgravity. The differentiated cells were assessed for their proliferation and migratory ability. Migration was assessed by observing and quantifying the migration of cells into an area cleared of cells (Fig. 3A). The quantification of percentage area of the scratch covered by the migrating cells (Fig. 3B) showed that the 6 h μg exposed and normal gravity-cultured PE-like cells migrated the most followed by 12 h μg exposed cells (P < 0.05). Finally the 18 h μg exposed PE-like cells migrated the least (P < 0.05), indicating a direct decrease in the migration ability of the EC with increased time of exposure of the progenitor cells under simulated microgravity.

The effect of preexposure to μg on the proliferative capacity of the EC-like cells was also assessed by quantifying the change in DNA over time in cultured cells. The data show that 6 and 12 h of preexposure to μg altered the initial rate of proliferation most significantly (Fig. 3C). In contrast, the PE-like cells preexposed to microgravity for 18 h proliferated faster than both (Fig. 3C, P < 0.05). As expected, the PE-like cells differentiated under normal gravity showed no inhibition in proliferation, but rather showed the greatest amount of proliferation (P < 0.05). These data collectively show that both the migratory and proliferative ability of ECs are impaired by preexposure of the EPCs to microgravity. While increased exposure time to μg impaired the migration of the microgravity-exposed cells, the proliferation of these cells seems to be enhanced by increased time of microgravity preexposure. This indicates a possible recovery of the proliferative ability of the PE-like cells on longer times of exposure of the EPCs to simulated microgravity. A similar recovery of immune functions of human PBMNCs after their short-term exposure to simulated microgravity has been reported [25]. Further investigation of this recovery mechanism may result in the development of possible countermeasures for treating cardiovascular deconditioning to enhance recovery [25,29,30].
A healthy endothelium is able to control and regulate thrombogenic properties and clotting cascades in the vasculature by releasing antithrombogenic factors into the blood stream. The ability of porcine EPC-derived ECs to inhibit clotting mechanisms has been shown before [13,15,31,32]. Hence, it is important to assess the ability of ECs derived from microgravity-preexposed progenitors to regulate clotting mechanisms. To evaluate the antithrombogenicity of these cells, we first investigated their ability to release soluble factors such as NO and PGE2 into the surrounding medium.
Prostacyclin and PGE2 have been known to be released from the endothelium after stimulation by vasoactive agents and thrombin [33]. Prostacyclin is predominantly released by the endothelium of large-diameter blood vessels, while the ECs from smaller diameter blood vessels predominantly release PGE2 [34]. Both of these factors contribute to platelet inactivation and regulate clotting mechanisms [33]. Another important molecule that regulates clotting mechanisms in the body is NO, which is produced by eNOS [33,35]. Both these molecules synergistically regulate antithrombogenicity in the body.
Our results show a decrease in secretion of soluble PGE2 by 6 h μg exposed cells (P < 0.05), while there was no significant difference in the levels of PGE2 between the other microgravity-exposed cells as well as the control cells cultured under normal gravity (Fig. 4A). The assessment of release of soluble NO indicated a similar trend where the amount of NO released by 6 h μg exposed cells was significantly lower (P < 0.05) when compared to other microgravity-exposed cells as well as normal gravity-cultured cells (Fig. 4B). These data show the ability of short-term microgravity exposure of EPCs to inhibit the regulation of release antithrombogenic molecules into the surrounding medium. However, with increased time of microgravity exposure, a potential recovery effect is also observed where the 12 and 18 h μg exposed cells secrete NO and PGE2 in levels comparable to the cells cultured under normal gravity. Previous reports have shown an increase in the release of both the factors by mature ECs when cultured under microgravity conditions [5]. However, the effect of short-term microgravity exposure of progenitor cells to alter the release of soluble factors by ECs derived from them has not been reported before. These data are in coherence with whole blood clotting and recalcified plasma clotting studies subsequently performed on these cells.
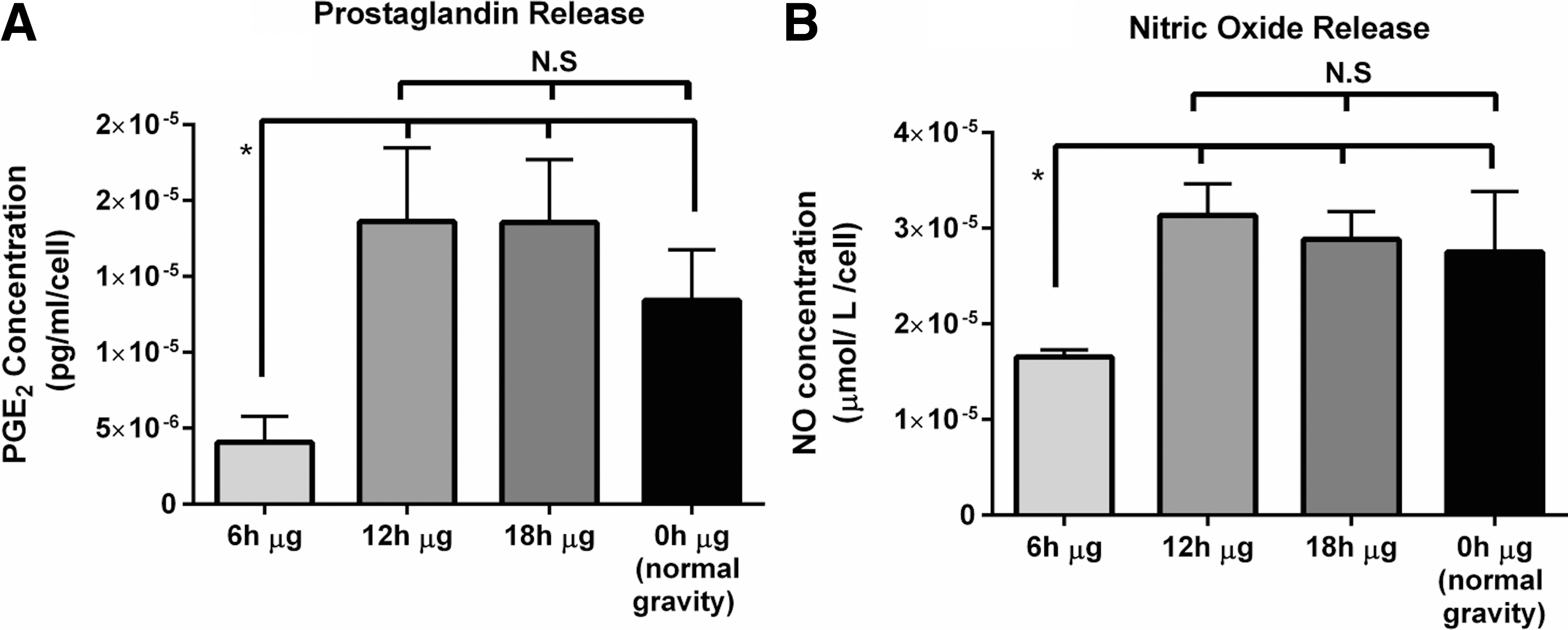
Representative results of quantification of release of
Plasma recalcification clotting profiles indicated that as expected, the TCP surface induced highest rate of plasma clotting within the first 90 s followed by 6 h μg exposed PE-like cells (Fig. 5A, P < 0.05). Following the clotting rate for 6 h μg exposed cells are the clotting rates for the 12 and 18 h μg exposed cells. Normal gravity-cultured cells clotted the next, while the last to clot was the plasma exposed to the PAEC control cells (P < 0.05).

Representative result showing quantification of
Similarly, the whole blood clotting kinetics showed that after 20 min, the average rate of whole blood clot formation was the highest for TCP surface (Fig. 5B, P < 0.05) followed by whole blood exposed to 6, 12, and 18 h μg exposed cells, whose average clotting rates were lower than TCP control (Fig. 5B). While the whole blood exposed to PAEC and cells cultured under normal gravity showed the greatest inhibition of clotting. These data collectively show that exposure to microgravity in the early progenitor stages negatively influences the antithrombogenic function of PE-like cell relative to those differentiated under normal gravity. The data show that within the three time points of μg exposure, the longer exposure times showed greater inhibition of clotting, which may be attributed to the increased ability to release soluble antithrombogenic factors, NO and PGE2.
Although previous reports have shown the ability of simulated microgravity to directly affect platelet functions [36], to our knowledge, the effect of simulated microgravity on antithrombogenic properties of ECs, such as inhibition of blood and plasma clotting, has not been reported so far. This work provides an insight into the microgravity-mediated alterations to regulation of clotting mechanisms by blood-derived ECs, which is important to assess their role in cardiovascular deconditioning during space travel.
Conclusions
The disruption of normal functioning of the endothelium has been shown to contribute significantly in weightlessness-induced cardiovascular deconditioning during space flight [37]. The focus of this study is to bridge the gap in knowledge on the effect of microgravity on circulating stem cells as they differentiate into ECs. This report also discusses the downstream function of the differentiated cells in terms of their migration and proliferative capabilities, release of soluble factors such as PGE2 and NO, and the ability to regulate clotting mechanisms in the body. While both the migration and proliferation of cells were inhibited by microgravity exposure, prolonged exposure of EPCs to microgravity resulted in decrease in the migration abilities of EPC-derived EC and the proliferative capacity of the ECs seemed to recover on longer time of exposure. The initial decrease and subsequent increase in release of soluble factors such as NO and PGE2 reaffirmed the possibility of recovery mechanisms being involved on prolonged exposure to weightlessness.
Finally, the assessment of whole blood and plasma clotting indicated that microgravity preexposure at the progenitor stage caused inhibition, in the ability of PE-like cells to delay blood or plasma clotting, while this effect was again nulled with increase in time of exposure of EPCs to microgravity. These data collectively provide some insight into the role of short-term exposure to low shear stress environment due to simulated microgravity, on cardiovascular deconditioning during space travel. Especially, the effect of microgravity on antithrombogenic properties of the endothelium has not been reported before. Future studies to understand alternate and synergistic mechanisms involved in regulations of cardiovascular complications such as thrombosis under microgravity will open new pathways for cell-based therapies for such complications.
Footnotes
Acknowledgment
This work is funded by National Aeronautics and Space Administration, Fundamental Space Biology Division, to J.B.A., grant# NNX13AM03G.
Author Disclosure Statement
No competing financial interests exist.