
Abstract
Recent studies have shown that secretion of bioactive factors from mesenchymal stem cells (MSCs) plays a primary role in MSC-mediated therapy; especially for bone marrow-derived MSCs (BMSCs). MSCs from dental apical papilla (SCAPs) are involved in root development and may play a critical role in the formation of dentin and pulp. Bioactive factors secreted from SCAPs actively contribute to their environment; however, the SCAPs secretome remains unclear. To address this and gain a deeper understanding of the relevance of SCAPs secretions in a clinical setting, we used isobaric chemical tags and high-performance liquid chromatography with tandem mass spectrometry to profile the secretome of human SCAPs and then compared it to that of BMSCs. A total of 2,046 proteins were detected from the conditioned medium of SCAPs, with a false discovery rate of less than 1.0%. Included were chemokines along with angiogenic, immunomodulatory, antiapoptotic, and neuroprotective factors and extracellular matrix (ECM) proteins. The secreted levels of 151 proteins were found to differ by at least twofold when BMSCs and SCAPs were compared. Relative to BMSCs, SCAPs exhibited increased secretion of proteins that are involved in metabolic processes and transcription and lower levels of those associated with biological adhesion, developmental processes, and immune function. In addition, SCAPs secreted significantly larger amounts of chemokines and neurotrophins than BMSCs, whereas BMSCs secreted more ECM proteins and proangiogenic factors. These results may provide important clues regarding the molecular mechanisms associated with tissue regeneration and how they differ between cell sources.
Introduction
T
Stem cells from dental apical papilla (SCAPs) are found in the root apex of immature permanent teeth [8]. These cells appear to be the source of odontoblasts and contribute to root formation [9]. For example, interruption of SCAPs development was found to result in a halted root development [10], and in combination with tissue engineering procedures, Huang et al. used SCAPs to regenerate vascularized human dental pulp in an emptied root canal and this was accompanied by formation of new dentin on the existing dentinal wall [11]. However, the understanding of SCAPs function and relevance in a clinical setting needs to be improved. One means to achieve this is identification of the wide array of proteins they secrete.
Therefore, in this study, we investigated the secretome of human SCAPs and compared the resulting profile to that of BMSCs, with the aim of achieving insights into the functional role of SCAPs secretions and the molecular basis of their actions.
Materials and Methods
Establishment, characterization, and analysis of cell proliferation rate of SCAPs and BMSCs cultures
Normal human impacted third molars with immature roots were collected from healthy patients (aged 16–24 years) (n = 5). Bone marrows were obtained from the mandibular bone of healthy patients (aged 20–30 years) (n = 3). Sample collection was approved by the Ethics Committee of the Health Science Center of Peking University (Beijing, China; IRB00001052-11060). SCAPs and BMSCs were isolated from dental apical papilla tissues and mandibular bone marrow, respectively. Cultures of MSCs were maintained in alpha-modified Eagle's minimum essential medium (Gibco) supplemented with 10% fetal bovine serum (Gibco) in 5% carbon dioxide at 37°C. The cells were used in experiments after three to five passages, and for each experiment, SCAPs and BMSCs had the same passage number.
SCAPs and BMSCs were characterized by flow cytometry using fluorescein-isothiocyanate-conjugated or phycoerythrin-conjugated antibodies specific for CD73, CD90, CD105, CD146, and CD34 (BD Biosciences) as previously described.
MSCs were seeded into 96-well plates at a density of 1.0 × 103 cells/well and then cultured for 7 days. Cell counting kit 8 (CCK-8; Dojindo) solution was added to each well of the plate and the absorbance was measured at 450 nm, every 24 h, according to the manufacturer's protocol.
Preparation of CM and protein digestion
SCAPs and BMSCs were seeded on 100-mm plates at a density of 20,000 cells/cm2. When MSCs reached 90% confluency, they were washed five times with phosphate-buffered saline and cultured in serum-free medium for 24 h. The CM were collected after centrifugation at 130 g for 10 min to remove the cellular debris and then passed through a 0.22-μm filter. The samples were concentrated, then air-dried, redissolved in triethylammonium bicarbonate (Promega), and finally, reduced with dithiothreitol at 55°C for 1 h. Next, iodoacetamide (Promega) was added to the samples and they were left for 1 h at room temperature, protected from light. Protein concentration was determined using a bicinchoninic acid (Thermo Fisher Scientific) assay.
TMT and HPLC-MS/MS analysis
Samples were digested with sequence-grade modified trypsin (Promega) and then labeled using the TMT reagent kit (Thermo Fisher Scientific) [12]. Ten fractions were collected by high-pH separation using an Aquity UPLC system (Waters Corporation) [13].
The fractions were resuspended in formic acid, separated by nanoLC, and analyzed by online electrospray tandem mass spectrometry. The analysis was performed with nanoAquity UPLC system (Waters Corporation), which was equipped with an online nanoelectrospray ion source and connected to a Q-Exactive hybrid quadrupole-orbitrap mass spectrometer (Thermo Fisher Scientific). Samples were separated by analytical column (Thermo Scientific Acclaim PepMap C18, 75 μm × 50 cm) with a linear gradient from 5% D to 30% D in 95 min. Full-scan MS spectra (m/z 350–1200) were acquired and three MS/MS events were analyzed. The samples were analyzed in triplicate.
Database searching and quantitative data analysis
MS was extracted by ProteoWizard version 3.0.5126 (Thermo Fisher Scientific) and analyzed using Mascot version 2.3 (Matrix Science). The human Uniprot-SwissProt database (release 2014_04_10; 20,264 reviewed entries) was used to search. A fragment-ion mass tolerance of 0.050 Da and a parent-ion tolerance of 10.0 ppm were equipped for Mascot.
Scaffold Q+ version 4.4.1.1 (Proteome Software, Inc.) was used to assess label-based peptide quantification and protein identification. Peptide identifications were accepted if the false discovery rate (FDR) was less than 1.0%, and protein identifications were accepted if the probability was more than 66.0% and at least one identified peptide was contained. Acquired intensities in the experiment were globally normalized across all acquisition runs. Individual quantitative samples were normalized within each acquisition run. The intensity of each peptide identified was normalized within the assigned protein. The reference channels were normalized to produce a 1:1 ratio. All normalization calculations were performed using medians to multiplicatively normalize the data. Differentially expressed proteins were determined using a Kruskal–Wallis test. String software (
Western blotting
Thirty micrograms of secretory protein were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred on a polyvinylidene fluoride membrane. The membrane was blocked with 5% (w/v) nonfat dried milk, incubated overnight at 4°C with primary antibodies against midkine (MDK) and angiopoietin-related protein 2 (ANGPTL2; Abcam), and then reacted with horseradish-peroxidase-conjugated secondary antibodies (Origene). Immunoreactive bands were visualized by enhanced chemiluminescence (Cwbiotech) at room temperature and then digitized using the Fusion FX image analyzer (Viber Loumat).
Real-time reverse transcription-polymerase chain reaction analysis
Total RNA was extracted from SCAPs and BMSCs (after the culture procedure described above) using TRIzol Reagent (Invitrogen) according to the manufacturer's instructions. cDNA was synthesized with oligo(dT) primers using a reverse transcriptase kit (Promega). The primer sequences were designed by primer bank (Supplementary Table S1; Supplementary Data are available online at
Statistical analysis
Statistical analysis was performed using SPSS version 15.0 (SPSS). Student's t-tests were used to assess the significance of differences between groups; a P value of <0.05 was considered an indicator of statistical significance. All experiments were repeated at least thrice (n ≥ 3).
Results
Proliferation of SCAPs and BMSCs
Cell proliferation was monitored over a period of 7 days postseeding, and we found that SCAPs showed an enhanced proliferation capacity versus BMSCs (Fig. 1A).
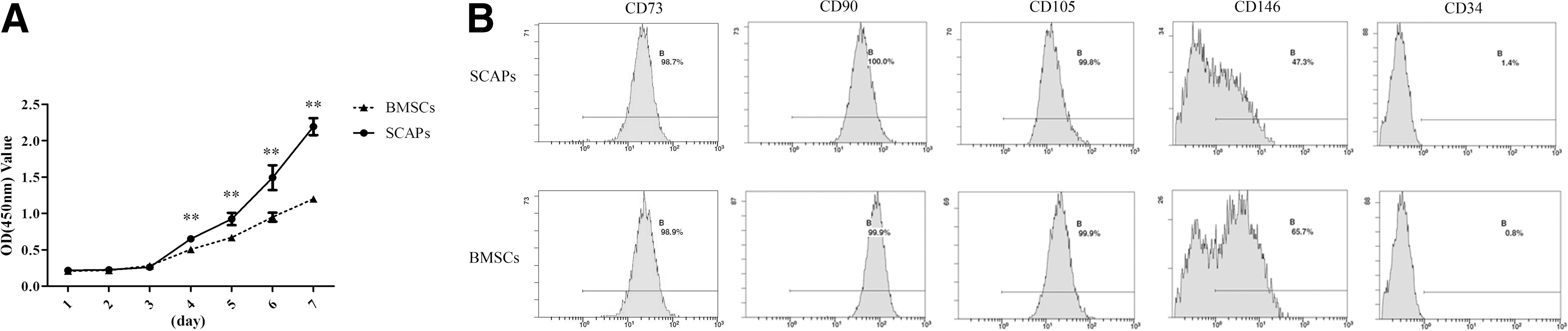
SCAPs exhibited increased cell proliferation compared with BMSCs.
Cell surface-marker analysis by flow cytometry showed that both stem cell types were positive for CD73, CD90, CD105, and CD146 and negative for CD34 (Fig. 1B).
Quantitative analysis of low-pH nano-HPLC-MS/MS
Cell death was rarely detected during the incubation period, as determined by microscopy. A total of 2,046 proteins were detected from the CM of the SCAPs, and the FDR was less than 1.0%. In addition, 74.3% of the proteins (1,520/2,046) were identified in at least two of the replicate experiments, including 1,172 proteins that were detected in all three experiments.
In the proteins that were identified in all three experiments, 296 (25.3%) proteins matched the GO terms “extracellular region” and 110 (9.5%) proteins matched the GO terms “extracellular matrix (ECM)” using DAVID Bioinformatics Resources. The extracellular region is the space external to the outermost structure of a cell, while ECM is a structure generated and secreted by cells to provide structural and functional support for cells or tissues. The identified ECM proteins contained a noticeable number of structural proteins like collagens and fibronectin, as well as multiple growth factors such as transforming growth factor beta 1 (TGFβ1) and ANGPTL2 (Supplementary Table S2). To facilitate the interpretation of the ECM proteins, a functional interaction network of the ECM proteins was searched using the String software. As illustrated in Fig. 2, those ECM proteins secreted by SCAPs belong to eight major functional classes: cell adhesion, cell proliferation, collagen organization and proteases, immune system, odontogenesis, angiogenesis, neuron development, and signaling molecules.

Functional interaction network of the ECM proteins in the SCAPs sectetome. String software was used to analyze the ECM proteins listed in Supplementary Table S2. ECM, extracellular matrix.
Then we analyzed the secretome of BMSCs (Supplementary Table S5), it is noticed that the identified secretory proteins from SCAPs and BMSCs, both included factors involved in angiogenesis, immunomodulation, chemotaxis, neuroprotection, antiapoptosis, and ECM formation; some of these are listed in Table 1. Most of the angiogenic factors and ECM proteins were significantly lower in SCAPs, when compared to BMSCs, whereas SCAPs exhibited significantly elevated secretion of chemokines and neurotrophins. On the whole, there were no significant differences for immunomodulatory and antiapoptotic factors.
BMSC, bone marrow-derived mesenchymal stem cells; PSM, peptide-to-spectrum matches; SCAP, stem cells from dental apical papilla.
When comparing the secretomes of SCAPs and BMSCs, we found significant differences of at least twofold in magnitude (P < 0.05; fold change ≥2 for each replicate) for 151 proteins. Of these, 83 were higher in SCAPs (Supplementary Table S3) and 68 were lower (Supplementary Table S4).
Bioinformatic analysis of the secretome
To further investigate the different functions of secretory proteins between SCAPs and BMSCs, we analyzed the ontology of the identified 151 significant different proteins (P < 0.05; fold-change ≥2 for each replicate) based on “cellular component” and “biological process” using the PANTHER classification system. After carrying out a GO analysis (Fig. 3A) in the cellular-component domain, the vast majority of proteins that were less secreted in SCAPs could be mapped to the GO terms (categories), “extracellular matrix” (35%) and “extracellular region” (61%). Of the 83 proteins significantly increased in SCAPs, 36% mapped to “cell part” and 21% mapped to “macromolecular complex”; these two categories included vimentin, ATP-dependent RNA helicase A, regulators of G-protein signaling, and eukaryotic translation initiation factors. In addition, 7% of the increased proteins were ECM proteins, such as stromal cell-derived factor 1 (SDF1), periostin, latent TGFβ-binding protein 4, and mimecan.

Bioinformatic analysis of proteins secreted in significantly different quantities by SCAPs (P < 0.05; ≥2-fold change vs. BMSC value).
Proteins in the biological process domain are listed in Fig. 3B and of the increased proteins 45 mapped to “metabolic process” and 33 mapped to “cellular process”; these two categories included 1-phosphatidylinositol 4-kinase, 5-bisphosphate phosphodiesterase, KH domain-containing RNA-binding signal transduction-associated protein 1 (KHDRBS1), Rho GDP-dissociation inhibitor 1, SWI/SNF complex subunit, and stress-70 protein (HSPA9). The category, “developmental process” covered 7% of the increased proteins and decreased proteins and 9% came under “immune system process.”
Validation of several differentially secreted proteins
To validate our proteomic findings, we selected several proteins that were secreted in quantities that differed between SCAPs and BMSCs, including neurotrophins [MDK; pleiotrophin (PTN)], development-related proteins [retinol-binding protein 1 (RBP1)], an angiogenic factor (ANGPTL2), antiapoptotic factors [insulin-like growth factor-binding protein 7 (IGFBP7); metalloproteinase inhibitor 4 (TIMP4)], immunomodulatory factors [follistatin-related protein 3 (FSTL3)], and cell cycle regulator (KHDRBS1). The expression of the MDK, PTN, RBP1, and KHDRBS1 genes were significantly higher in SCAPs, as shown by qRT-PCR (Fig. 4A). Western blotting indicated that SCAPs had a higher secretion of MDK proteins and a lower secretion of ANGPTL2 proteins compared with BMSCs (Fig. 4B). These results were consistent with our proteomic analysis. Also, in agreement with the proteomic results was the finding that IGFBP7, TIMP4, ANGPTL2, and FSTL3 gene expression were all significantly decreased in SCAPs.
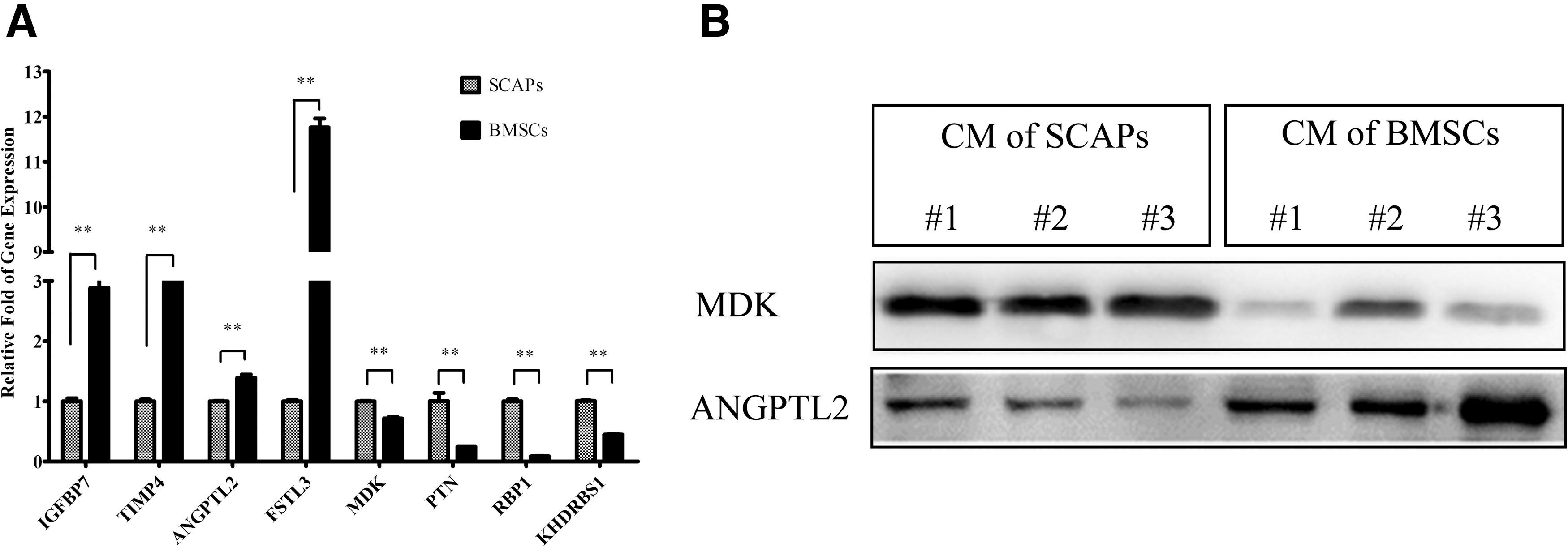
Verification of proteins secreted in significantly different quantities by SCAPs and BMSCs (P < 0.05; ≥2-fold change).
Discussion
The spectrum of regulatory and trophic factors secreted by MSCs is generally referred to as the MSC secretome. To define the role of MSC-secreted factors in tissue regeneration, one should start with biomolecular profiling or secretome analysis of MSCs. The effect of serum must be considered in MSC secretome analysis in vitro. Serum is often supplemented to the cell culture medium for cell growth. However, the extreme complexity of proteins in serum may overlap components of MSC secretome, which are usually secreted at low concentrations (as low as ng/mL) in the culture media [14]. The existence of serum will interfere with protein detection and make their recovery difficult. To solve this problem, MSCs can be cultured in a serum-free medium for a defined time, which has been successfully applied to secretome analysis in a number of studies [15 –17]. Thus, we collect the CM under a serum-free condition in this study.
Of the proteins that were increased ≥2-fold in SCAPs, 35.8% were intracellular proteins. There was little cell death (associated with release of cell contents) during the incubation period, but most of the identified proteins have been previously reported as present in exosomes (secreted microvesicles); thus, it is worth considering whether exosome formation and secretion is linked to the secretome differences we have observed. Exosomes often contain common cytosolic proteins, intracellular and intercellular signal transduction proteins, and various transmembrane proteins [18]. It has been found that MSCs can produce higher quantities of exosomes than other cells [19]. Several studies have reported that MSC-derived exosomes perform functions similar to those of MSCs, such as repairing tissue, suppressing inflammatory responses, and modulating the immune system [20,21].
We found that the proliferation capacity of SCAPs was significantly greater compared with BMSCs; this may be due to differences in the developmental stage of the respective cell types. Our analysis of their secretomes showed that SCAPs secreted much more proteins involved in metabolic processes, especially nucleic acid binding or transcription, such as KHDRBS1, Rho GDP-dissociation inhibitor 1, eukaryotic translation initiation factors, and heat shock protein (HSPs). KHDRBS1 is also called sam68 (the Src-associated substrate in mitosis of 68 kDa), which is involved in cell cycle regulation. It has been demonstrated that Sam68-defcient cells exhibited markedly decreased growth rate due to elongation of the G2-M phase [22]. In addition, overexpression of a Sam68 isoform with the deletion of its KH domain resulted in the suppression of cell growth and cyclin D1 expression, which could be rescued by Sam68 transfection, suggesting that Sam68 participates in the control of cell proliferation by promoting the S phase entry [23]. Rho GDP-dissociation inhibitor 1 plays a key role in the regulation of signaling through Rho GTPases. Increased expression of Rho GDP-dissociation inhibitor 1 may have important implications in cell functions involving massive cytoskeletal changes such as cell proliferation, differentiation, migration, and adhesion. It is reported that Rho GDP-dissociation inhibitor 1 was upregulated in dental pulp stem cells (DPSCs) compared to BMSCs [24]. HSPs function as molecular chaperones in the stabilization of intracellular proteins, repairing damaged proteins, and assisting in protein translocation. It was reported that the high levels of HSP90, HSC70, and HSPA9 in mouse embryonic stem cells were eliminated when the cells underwent differentiation [25].
MSCs retain the capacity to regenerate the unique microenvironments from which they are derived in vivo [24,26 –28]. In our study, 35% of proteins that were less secreted in SCAPs were ECM proteins and only 7% of the increased proteins in SCAPs were ECM proteins, which is consistent with the function in vivo. SCAPs and BMSCs contribute to dentin and bone formation, respectively. Actually, above 30% of the acellular part of bone consists of the organic components, and only 20% of dentin consists of the organic components by weight.
Angiogenesis, the formation of capillaries from preexisting blood vessels, is an important process in tissue engineering. SCAPs have been shown to secrete proangiogenic factors and have a predominantly proangiogenic impact on endothelial migration and tube formation [29]. In this study, proangiogenic and antiangiogenic proteins were compared between SCAPs and BMSCs. The results indicated that secretion of proangiogenic factors was lower in SCAPs, such as vascular endothelial growth factor C (VEGFC) and ANGPT1. VEGF family is the most important growth factor that regulates the process of angiogenesis. VEGFC is a ligand of the VEGFR-2 and VEGFR-3 receptors, which activates blood-vessel tip cells [30]. ANGPT1 stimulates mural coverage and basement membrane deposition, thereby contributing to vessel maturation and stability [31]. In addition, the secretion of homing- and transmigration-related factors, such as adhesion molecules and matrix metalloproteinases (MMPs), was lower in SCAPs. The adhesion molecules, integrins and vascular cell adhesion protein 1, participate in stem cell homing by adhesion to ECM and immunoglobulin superfamily molecules [32 –34]. MMPs are matrix-degrading enzymes that promote endothelial cell migration and tube formation by remodeling the basement membrane, by executing directional matrix proteolysis, or by exposing chemotactic cryptic motif sites in the ECM [31]. Meanwhile, the antiangiogenic proteins, such as TIMPs and thrombospondin, were also significantly decreased in SCAPs. TIMPs and thrombospondin function as negative regulators of angiogenesis mainly by interacting with proteases and VEGF. Thus, the effect of angiogenesis seemed to be more active in BMSCs compared to SCAPs. Given microenvironments the MSCs derived from, it is hypothesized that BMSCs may be more effective for angiogenesis. However, further study of angiogenesis is still needed for a better understanding about the differences between SCAPs and BMSCs.
Our secretome profiles indicate that neurotrophins, such as MDK and PTN, were secreted in larger amounts by SCAPs when compared to BMSCs. MDK, also known as neurite growth-promoting factor 2 (NEGF2), is a basic heparin-binding growth factor that forms a family with PTN (also known as NEGF1) [35]. They are expressed in neural precursor cells and promote their growth [36,37]. It was recently reported that the secretions of BMSCs exhibited neuroprotective effects and enhanced tissue repair in the central nervous system [6]. In addition, SCAPs secreted significantly larger amounts of SDF1, which is a member of the CXC cytokine subfamily and a widely expressed chemotactic cytokine that mediates cell migration through its binding to CXC chemokine receptor 4 (CXCR4) [38]. Our results showed that SCAPs secreted significantly greater quantities of chemokines and neurotrophins, suggesting that SCAPs may be good candidates for use in neural tissue repair.
Some proteins involved in odontogenesis or osteogenesis were secreted more in SCAPs than BMSCs, such as TGFβ, RBP1, and SPARC-related modular calcium-binding protein 1 (SMOC1). TGFβ is expressed during tooth formation and can induce preodontoblast differentiation and formation of functional odontoblast-like cells in vitro [39,40]; it has also been shown to control mouse tooth size and development in vivo [41]. The overexpression of RBP1 could promote osteogenic differentiation of BMSCs [42]. During the cap stage of crown development, RBP1-positive cells were mainly localized to the SCAPs [43]. The secretion of RBP1 was much higher in SCAPs compared to BMSCs, suggesting that RBP1 would be a good marker for SCAPs. Choi et al. showed that knockdown of SMOC1 significantly inhibited mineralization and the expression of osteoblast differentiation markers in BMSCs, while overexpression of SMOC1 substantially promoted osteoblast differentiation [44]. The condition medium of BMSCs was proved to accelerate osteogenesis in vivo and had an osteo/odontogenic inductive effect on DPSCs in vitro [5,45,46]. It is speculated that the secretome of SCAPs plays an important role in dentin regeneration and the condition medium from SCAPs may be used for the induction of dentin and bone regeneration both in vitro and in vivo.
In conclusion, our study provides insights into the function and molecular details of SCAPs secretions, and it has shown significant differences between the secretomes of SCAPs and the more widely studied and utilized BMSCs. SCAPs were found to secrete angiogenic, immunomodulatory, antiapoptotic, chemokine, and neuroprotective factors. Among the most significant differences between SCAPs and BMSCs was that SCAPs secreted considerably higher levels of chemokines and neurotrophins. These results may shed some new light on the molecular mechanisms involved in tissue regeneration, while also demonstrating the potential of SCAPs as an alternative cell source for tissue engineering and therapeutic applications.
Footnotes
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (no. 81170928 and no. 81541110) and the Peking University 985 project (no. 2013-4-01). We gratefully acknowledge the assistance of Wang Feng from BioyongTech for technical data analysis and her expertise in proteomics analysis. The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.