
Abstract
Alport syndrome (AS) is a hereditary glomerulopathy caused by a mutation in type IV collagen genes, which disrupts glomerular basement membrane, leading to progressive glomerulosclerosis and end-stage renal failure. There is at present no cure for AS, and cell-based therapies offer promise to improve renal function. In this study, we found that human first trimester fetal chorionic stem cells (CSC) are able to migrate to glomeruli and differentiate down the podocyte lineage in vitro and in vivo. When transplanted into 7-week-old Alport 129Sv-Col4α3tm1Dec/J (−/−) mice, a single intraperitoneal injection of CSC significantly lowered blood urea and urine proteinuria levels over the ensuing 2 weeks. In addition, nearly two-thirds of transplanted −/− mice maintained their weight above the 80% welfare threshold, with both males and females weighing more than age-matched nontransplanted −/− mice. This was associated with less renal cortical fibrosis and interstitial inflammation compared to nontransplanted mice as shown by reduction in murine CD4, CD68, and CD45.2 cells. Transplanted CSC homed to glomeruli, where they expressed CR1, VEGFA, SYNAPTOPODIN, CD2AP, and PODOCIN at the RNA level and produced PODOCIN, CD2AP, and COLIVα3 proteins in nontransplanted −/− mice, indicating that CSC have adopted a podocyte phenotype. Together, these data indicate that CSC may be used to delay progression of renal pathology by a combination of anti-inflammatory effects and replacement of the defective resident podocytes.
Introduction
A
The only current treatment for AS is blockade of the renin–angiotensin system, and proposed treatments include collagen receptor blockade, anti-microRNA therapy, and stem cell therapy [2]. The rationale of stem-based therapy is that stem cells isolated from healthy donors will migrate and engraft in renal glomeruli, where they may differentiate into functional podocytes producing a new functional GBM. We previously showed that human first trimester fetal blood-derived mesenchymal stem cells (MSC) injected intraperitoneally into fetal mouse recipients migrated to the kidneys where they engrafted in renal glomeruli [3]. However, phenotype rescue by direct cell replacement is challenged by the low level of donor cell engraftment and poor differentiation capacity of the donor cells [4].
Col4α3tm1Dec/J mice are deficient in collagen type IV alpha 3 (Col4α3) chains and suffer from progressive glomerulosclerosis, with thickening and lamellation of the GBM, segmental glomerular scarring, tubular atrophy, tubulointerstitial fibrosis, and inflammation [5]. The rate of disease progression depends on the genetic background. On a 129Sv background (129Sv-Col4α3tm1Dec/J mice), inactivation of col4α3 leads to proteinuria by 35 days, elevated blood urea from 50 days onward, and end-stage renal failure by 66 days; whereas on a C57BL/6 background (C57BL/B6-Col4α3tm1Dec/J mice), these events occur later at 110, 150, and 194 days, respectively [6].
We previously showed that whole bone marrow from wild-type Col4α3+/+ mice transplanted into Col4α3−/− mice produced the missing Col4α3 chain and contributed to improved renal function. However, transplantation of expanded MSC from +/+ mice into −/− mice failed to improve renal function, suggesting either that culture conditions did not maintain cellular plasticity of bone marrow MSC or that other cells, such as hematopoietic stem cells, were involved in restoring renal function [7]. Using a similar model, Sugimoto et al. reported partial restoration of Col4α3 chain expression as well as improvement of glomerular structure and kidney histology following wild-type bone marrow transplant [8]. LeBleu et al. also showed that improvement of renal function in Col4α3−/− mice was associated with the expression of the missing α3 chain of type IV collagen [9].
In all cases, the origin of the cells responsible for the improved renal function was not established, but the results suggest that repair of GBM architecture and glomerular integrity is attributable to expression of Col4α3 chain from podocyte-differentiated donor cells. Using a different AS mouse model, that is, ColIVα5−/− mice, Sedrakyan et al. suggested that mouse stem cells isolated from amniotic fluid delayed interstitial fibrosis and progression of glomerular sclerosis, ameliorating the decline in kidney function [4]. However, donor cells failed to differentiate into podocytes and produce the collagen IVα5 chain, suggesting that improved renal function may have been achieved by production of anti-inflammatory cytokines.
The placenta is a potential source of readily obtainable stem cells throughout pregnancy. We recently isolated and characterized human fetal stem cells derived from first-trimester chorion [ie, chorionic stem cells (CSC)] 10 . CSC have a spindle-like morphology, are capable of trilineage differentiation (osteogenic, adipogenic, and chondrogenic), and demonstrate high tissue repair in vivo [10]. Over 95% of the cell population lack expression of CD14, CD34, and CD45, but express the mesenchymal markers CD105, CD73, CD44, vimentin, CD29 and CD90, with a subset of cells also expressing the pluripotency markers NANOG, SOX2, cMYC, KLF4, SSEA4, SSEA3, TRA-1-60, and TRA-1-81, and being able to form embryo bodies containing cells from the three germ lineages.
In this study, we provide evidence that human CSC can be differentiated down the podocyte lineage in vitro and in vivo, and delay progression of renal pathology when injected in Col4α3−/− mice, preventing weight loss and decreasing levels of cortical fibrosis and interstitial inflammation.
Materials and Methods
Animals
Alport (129-Col4α3tm1Dec/J) mutant (Col4α3−/− ) and wild-type (Col4α3+/+ ) mice (Jackson Laboratory) were housed in filter cages with a 12-h light–12-h dark cycle (21°C), with water and wet chow (Purina) ad libitum to avoid dehydration, which can affect blood urea measurements. Mice were weaned at 30 ± 1 days and culled at 9 weeks of age. All animals were handled in accordance with good animal practice as defined by the British Home Office Animal Welfare Legislation, and animal work was approved by the Institutional Research Ethics Committee (Imperial College London, London, United Kingdom).
Glomeruli isolation
Kidneys from wild-type and Alport mice were pushed through a series of sieves (pore sizes 150, 106, and 45 μm) using the plunger of a 20-mL syringe. Glomeruli retained on the 45-μm sieve were collected into a tube and centrifuged at 1,000 rpm for 10 min.
Cell culture
Collection of surplus human chorionic villi samples was approved by the Research Ethics Committees of Hammersmith & Queen Charlotte's Hospital and of University College London Hospital (UCLH) in compliance with national guidelines (Polkinghorne). CSC were selected by adherence to plastic, further expanded at 10,000 cells/cm2 at 37°C in 5% CO2 incubator, and studied at passage 4–8. Their fetal origin was confirmed by FISH for X and Y chromosomes on male samples. The cells were fully characterized as previously reported [10], showing the characteristics of stromal MSC-like cells, that is, capacity to differentiate down the osteogenic, adipogenic, and chondrogenic pathway, and expression of CD73, CD90, and CD105. CSC were cultured for 3 weeks in Dulbecco's modified Eagle's medium (DMEM)–high glucose (Invitrogen) supplemented with 10% fetal bovine serum (Biosera), 100 IU/mL penicillin, and 100 μg/mL streptomycin (Invitrogen), that is, growth medium (D10) on noncoated plastic dishes, or on plastic dishes coated with human type IV collagen (Sigma).
Temperature-sensitive conditionally immortalized human podocyte cell line, derived by Saleem et al. from fresh normal human pediatric kidney specimens, were used as positive controls (gift from Moin Saleem, University of Bristol, Bristol, United Kingdom). These cells were originally by incorporating a temperature-sensitive SV40 gene that enables the cells to proliferate at a permissive temperature (33°C) and differentiate at a nonpermissive temperature (37°C), as evidenced by cell morphology and upregulation of nephrin, synaptopodin, podocin, and VEGFA expression [11]. The podocytes were cultured for 21 days at 37°C in six-well plates in RPMI 1640 medium supplemented with glutamine (Invitrogen), 10% fetal calf serum (Biosera), antibiotics, and 1% insulin transferrin sodium selenite (Sigma).
Fluorescence immunostaining and confocal microscopy
CSC were grown exponentially on 10-mm coverslips before being fixed in 4% PFA, 250 mM HEPES (pH 7.6; 10 min, 4°C), refixed in 8% PFA, 250 mM HEPES (pH 7.6; 50 min, 4°C), and rinsed thrice with phosphate-buffered saline (PBS). After fixation, the cells were incubated (30 min) with 20 mM glycine in PBS, blocked (1 h) with PBS+ [PBS supplemented with 1% bovine serum albumin (BSA), 0.2% fish skin gelatin, 0.1% casein; pH 7.6], incubated (2 h) with anti-NHPS2 or podocin (1:1,000; Sigma) in PBS+, washed (5× over 1.5 h) in PBS+, incubated (1 h) with secondary antibodies (Alexa 488 goat anti rabbit) in PBS+, rinsed (overnight, 4°C) in PBS+, and mounted in VectaShield labeled with DAPI (Vector Laboratories). Fluorescence confocal laser-scanning microscopy images were collected on a Leica TCS SP5 (×400 PL APO oil objective) and transferred to Adobe Photoshop (Adobe Systems).
Chemotaxis assay
CSC suspension (100 μL of 107 cells/mL in DMEM-0.5% BSA) was placed in the upper compartment of a chemotaxis chamber. Chemoattractants (glomeruli from +/+ or −/− mice, or DMEM-0.5% BSA) were placed in the lower compartment, separated by an 8 μm polycarbonate filter (Neuroprobe). The cells were allowed to undergo chemotaxis (1 h). The filter was then removed, washed, fixed, and stained (1% hematoxylin; Sigma). Ten random fields were counted at × 40 magnification by a blinded observer (triplicates). The migration index (MI) was calculated as the ratio of the number of cells migrating toward the chemoattractant to the number of cells migrating toward media alone.
Cell transplantation
Cells (106 in 10 μL PBS, pooled from five different donors to reduce interdonor differences) were injected intraperitoneally in 7-week-old Col4α3−/− or wild Col4α3+/+ (n = 25/group). Animal weight was recorded thrice per week and mice were culled 2 weeks after transplantation or when weight loss exceeded 20% of the maximum previously achieved weight, as mandated by the British Home Office.
Quantitative real-time reverse-transcription polymerase chain reaction
Total RNA (n = 8 mice/group) was extracted from the glomeruli using TRIzol (Invitrogen) and cDNA synthesized using random primers and 1 μL of 200 U M-MLV Reverse Transcriptase in the presence of dNTPs (Promega Corp.) (10 min, 75°C; 120 min, 42°C and 10 min, 75°C). Quantitative real-time RT-PCR (QRT-PCR) was performed with the ABI Step-One Plus Sequence Detector (Applied Biosystems). We used primers amplifying sequences of the β-actin gene (Accession No. NM_001101) present in humans, but not in mice, to determine the amount of human cellular cDNA in samples (primer specificity confirmed by absence of amplification of mouse cDNA), and primers common to both human and mouse to determine the total cDNA in each sample, as previously described [12]. For both sets, the absence of dimer formation was confirmed using Dissociation Curves 1.0 software (Applied Biosystems). Human–mouse chimerism was estimated as a ratio. Serial dilution of human cDNA in mouse cells formed the calibration curves. The primer sequences are shown in Table 1.
RT-PCR, reverse transcription polymerase chain reaction.
Histology
Fresh kidneys (n = 6 mice) were fixed in Bouin's fixative (Sigma) for 4 h, dehydrated using serial dilutions of ethanol, embedded in wax, sectioned, and stained with picrosirius red stain. Five random nonoverlapping fields were assessed at ×200 magnification by a blinded observer, under polarized or white light with an Olympus BX51 microscope.
Immunohistology
Fresh kidneys were removed and fixed in a solution containing 1% PFA, 0.075 M L-lysine, 0.01 M sodium periodate, and 0.037 M phosphate buffer (all reagents from Sigma) for 4 h at 4°C. Tissues were then stored overnight into 7% sucrose in PBS (Sigma) at 4°C, immersed in OCT compound (VWR), and snap frozen before being stored at −80°C for further analysis. For immunofluorescence, 4-μm sections were air dried (2 h), fixed in acetone (10 min, 4°C), air dried for a further 2 h, and denatured (1 h, 4°C) using a solution of 6 M urea and 0.1 M glycine in PBS (pH 3.5, all reagents from Sigma). Slides were then washed with PBS and incubated overnight at 4°C with a Col4a3 primary antibody diluted 1:2,000 in 7% nonfat dry milk. The Col4a3 antibody (kindly donated by Dr. Dominic Cosgrove, Boys Town National Research Hospital, Omaha, NE) is an affinity-purified rabbit polyclonal antibody raised against a peptide mapping the NC1 region of Col4α3. This antibody has been tested for cross-reactivity by the provider and reacts with human and mouse [13]. The presence of donor cells was visualized using a rabbit monoclonal antibody raised against human vimentin (ab137867; Abcam). Slides were then washed in PBS, incubated at room temperature for 1 h with an FITC-conjugated secondary antibody, washed with PBS, and mounted with Vectashield containing DAPI (Vector Laboratories) for visualization using a confocal laser-scanning microscope Leica TCS SP5 (×1,000 PL APO oil objective; Leica).
Immunohistochemistry
Immunoperoxidase staining for the T-helper cell marker CD4 (BD Biosciences), the leukocyte marker CD45.2 (eBiosciences), and the macrophage marker CD68 (Abcam) was performed on periodate-lysine-paraformaldehyde (PLP)-fixed kidney cryostat sections (4 μm). Sections were incubated overnight at 4°C with primary antibody. For the biotin-conjugated CD45.2 antibody, endogenous biotin and avidin were blocked before the addition of the primary antibody, using biotin and avidin block solutions, respectively (Vector Laboratories). Sections were then washed with PBS, incubated with the appropriate horseradish peroxidase (HRP)-conjugated secondary antibody (Golden Bridge International), washed with PBS, and visualized using DAB. The sections were then counterstained with hematoxylin, dehydrated, and mounted using DPX (Sigma).
Blood urea analysis
Blood samples were centrifuged at 1,300 g (10 min, 4°C) and the supernatant was stored at −80°C until analysis. Urea was measured using a urea/ammonia detection kit (R-Biopharm), according to the manufacturer's instructions. All samples were analyzed at the same time to avoid batch variation.
Measurement of proteinuria/hematuria
Urine was collected from 9-week-old mice and proteinuria was quantitatively measured using mouse albumin and creatinine ELISA (albumin urinary level/creatinine urinary level) (Exocell), according to the manufacturer's instructions. Analysis was performed by two observers blinded as to whether each sample was from transplanted or nontransplanted −/− or +/+ groups.
Western blotting
Total protein was extracted using the RIPAE buffer containing protease inhibitor cocktail and PMSF (Sigma). Protein concentrations were determined using the BCA assay (Thermo Scientific) with BSA as standard. Proteins were run on 8% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), transferred to nitrocellulose membranes, blocked with milk, and incubated with primary antibodies for human-specific COLIVa3 (160–190 kDa), PODOCIN (42 kDa; Santa Cruz), and CD2AP (71 kDa; Millipore). Membranes were incubated with secondary HRP-conjugated anti-goat IgG (Santa Cruz) and proteins detected using enhanced chemiluminescence (Thermo Scientific). GAPDH was used as a loading control (Millipore).
Statistical analysis
Data are expressed as mean ± SEM (standard error of the mean) or median and range. Parametric and nonparametric statistics were applied after testing distributions on histograms. P < 0.05 was considered significant.
Results
CSC cultured on human type IV collagen or cocultured with glomeruli express podocyte markers and migrate to glomeruli in vitro
CSC were cultured for 3 weeks in growth medium (D10), either on noncoated plastic dishes or on plastic dishes coated with human collagen IV (CSC on COLIV). Alternatively, CSC were cocultured without cell contact, with freshly isolated COL4α3−/− and COL4α3+/+ glomeruli (CSC-glomeruli coculture). After 3 weeks of culture, the expression level of various podocyte markers was analyzed by QRT-PCR (Fig. 1A). The podocyte line expressed NEPHRIN, a gene involved in renal filtration; PODOCIN, expression of which is restricted to mature podocytes; VEGFA, produced during kidney morphogenesis to guide endothelial cells toward glomeruli; SYNAPTOPODIN, an actin-associated gene; CD2AP, which regulates the translocation of dendrin to reorganize podocyte cytoskeleton and stabilize the slit diaphragm; and CR1, complement receptor 1 which protects podocytes from complement attack.
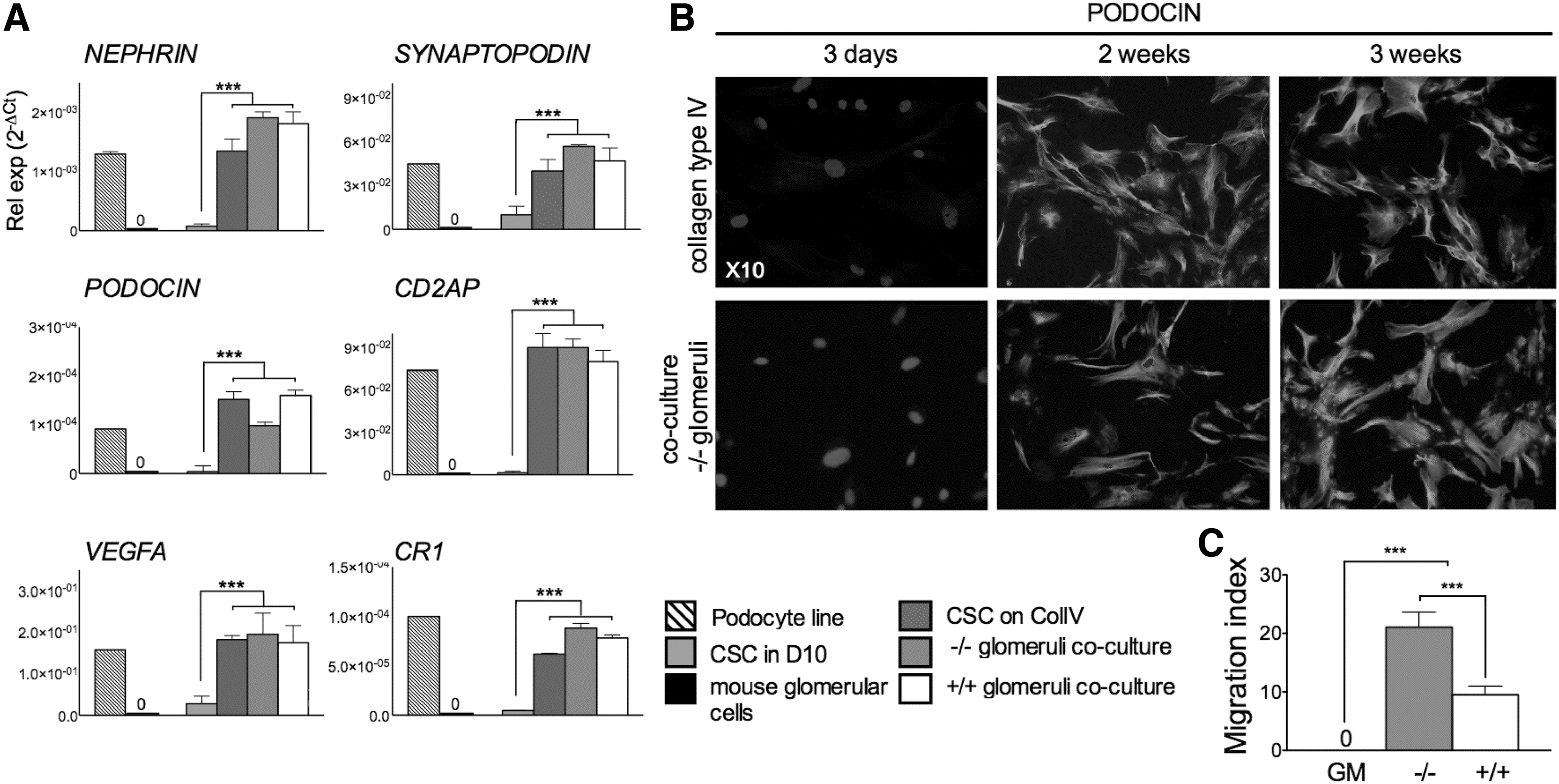
Chorionic stem cells (CSC) cultured in permissive conditions express podocyte markers and migrate to glomeruli in vitro.
The expression of these genes was almost undetectable in CSC cultured alone in growth medium on noncoated dishes. In contrast, the expression of all markers was upregulated when CSC were cultured on dishes coated with human collagen IV. Expression of NEPHRIN, SYNAPTOPODIN, and CR1 was further upregulated when the cells were cocultured with −/− glomeruli, although PODOCIN expression was lower. The expression levels of VIMENTIN and FIBRONECTIN remained unchanged in CSC cultured on collagen type IV or by coculture with glomeruli (data not shown).
Confocal immunofluorescnce showed that CSC cultured for 3 weeks on human type IV collagen or cocultured with −/− glomeruli expressed PODOCIN at a protein level (Fig. 1B). These data indicate that CSC have the potential to differentiate down the podocyte lineage. Next, we used a chemotaxis assay where CSC were allowed to migrate for 1 h toward freshly isolated Col4α3−/− or Col4α3+/+ glomeruli ex vivo. CSC did not passively migrate toward growth medium (GM) alone, but showed high chemotaxis toward +/+ glomeruli and significantly greater chemotaxis toward −/− glomeruli (9.5 ± 1.5 MI vs. 21.1 ± 2.6 MI, respectively) (Fig. 1C). These results indicate that −/− glomeruli produce soluble factors that may stimulate migration and differentiation of CSC and rationalize the use of CSC for the treatment of AS.
CSC transplanted into −/− mice engrafted into glomeruli, expressed podocyte markers, and produce PODOCIN and the missing Col4α3 protein
We injected 106 CSC intraperitoneally into 7-week-old −/− and +/+ mice and assessed the fate of donor cells 2 weeks later. CSC injection was performed at 7 weeks postnatally because analysis of the disease history in −/− mice revealed the presence of blood and protein in urine at this age, whereas the levels of blood urea, which is a robust measure of renal function in mice, was still comparable to the values found in +/+ mice. Donor cell engraftment in isolated glomeruli was determined by QRT-PCR using human-specific and nonspecific primers for the housekeeping gene β-actin, as previously described [14]. Results showed that transplanted CSC homed to glomeruli, with a 10.8-fold higher engraftment in −/− (n = 8) compared to +/+ glomeruli (n = 8) (Fig. 2A).

Transplanted CSC engrafted into −/− glomeruli and expressed podocyte markers.
Using human-specific antibody against vimentin protein, histological analysis of kidney sections from transplanted and nontransplanted −/− mice revealed the presence of donor CSC in glomeruli, with some donor cells being also visible outside the glomeruli of CSC-transplanted −/− mice (Fig. 2B). Engrafted CSC expressed the podocyte markers CR1, VEGFA, SYNAPTOPODIN, and CD2AP (Fig. 2C). Human specificity of the primers was verified using RNA from the kidneys of −/− nontransplanted mice, which showed absence of amplification. At the protein level, engrafted CSC produced PODOCIN, CD2AP, and COL4A3, which are absent in nontransplanted −/− and +/+ mice (Fig. 2D, E).
Improvement of the −/− phenotype
Nontransplanted Col4α3 −/− mice (n = 25) progressively lost weight between 7 and 10 weeks. By 9 weeks of age, 63% mice dropped their weight below the 20% endpoint level mandated by the British Home Office and had to be culled. In contrast, all CSC-transplanted Col4α3 −/− mice (n = 48) maintained their weight until 9 weeks (Fig. 3A), at which age both transplanted Col4α3 −/− males and females were heavier than age- and sex-matched nontransplanted Col4α3 −/− mice, with their weight being similar to Col4α3 +/+ mice (Fig. 3B).

CSC transplantation prevented weight loss in −/− mice and reduced levels of blood urea and cortical fibrosis.
Next, we measured blood urea in nontransplanted Col4α3 +/+ and Col4α3 −/− mice over 90 days. Levels remained under 20 mM until day 90 in +/+ mice (upper panel of Fig. 3C, n = 41), while all Col4α3 −/− mice showed elevated blood urea levels by 58 days of age (lower panel of Fig. 3C, n = 45). In contrast, urea levels were lower in 9-week-old Col4α3 −/− mice transplanted with CSC 2 weeks before than in nontransplanted Col4α3 −/− mice (18.8 ± 2.0 mM, n = 25 vs. 29.9 ± 0.7 mM, n = 10; mean ± SEM, P < 0.01), with 64% of transplanted Col4α3 −/− mice showing levels similar to Col4α3 +/+ mice (Fig. 3D). Quantification of urine proteinuria (albumin/creatinine level) revealed a significant reduction in Col4α3 −/− mice transplanted with CSC compared to their nontransplanted counterparts (35.6 ± 1.8, n = 21 vs. 47.5 ± 3.2, n = 16; mean ± SEM, P < 0.01) (Fig. 3E).
CSC transplantation lowered tubulointerstitial fibrosis and reversed cortical inflammation
We next measured tubulointerstitial fibrosis within the cortex using picrosirius red staining (PSR). PSR in kidneys from CSC-transplanted Col4α3 −/− mice was lower than levels found in nontransplanted Col4α3 −/− mice (0.089 ± 0.0018, n = 6 vs. 0.040 ± 0.0015, n = 6, P < 0.001), indicating a decrease in tubulointerstitial fibrosis (Fig. 4A, B). Quantification of renal inflammation showed that CSC transplantation reduced the number of glomerular T-helper cells (anti-CD4), macrophages (anti-CD68), and hematopoietic cells (anti CD45.2) in Col4α3 −/− mice indicating a marked decreased in cortical inflammation (Fig. 4C, D).
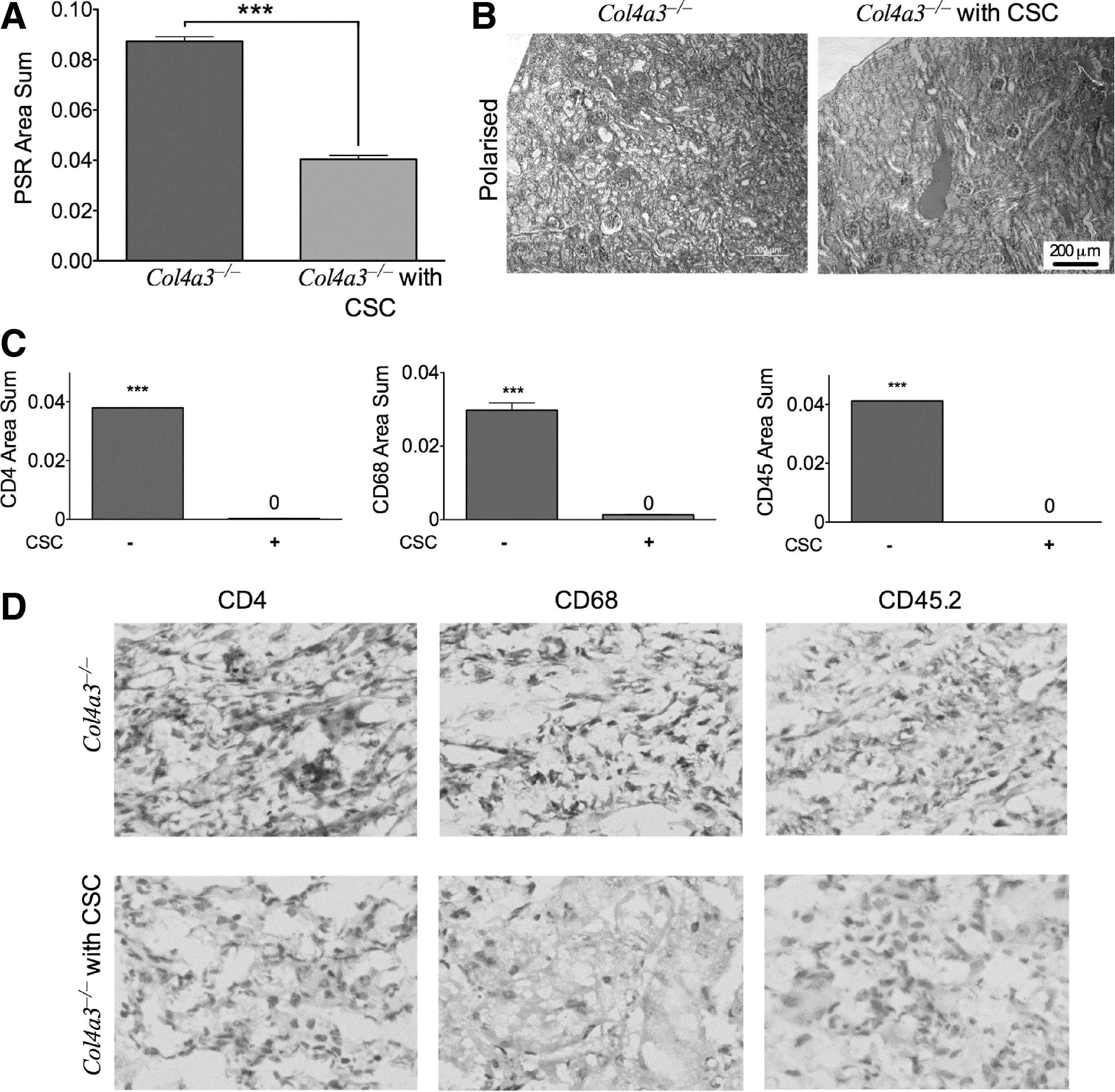
CSC transplantation reduced cortical inflammation in −/− glomeruli.
Col4a3 mutation downregulated endogenous murine podocyte gene expression, and CSC transplantation partially restored renal mRNA expression levels
We next assessed whether the presence of exogenous cells modified gene expression of resident podocytes. Results showed that expression of murine Nephrin, Podocin, Synaptopodin, Cd2ap, Cr1, and Vegfa was higher in the glomeruli of nontransplanted +/+ mice compared to nontransplanted −/− mice (Fig. 5). However, CSC transplantation upregulated the renal mRNA expression of Nephrin, Podocin, Synaptopodin, Cd2ap, and Vgfa, suggesting that the decrease in glomerular inflammation in transplanted mice was associated with restored podocyte activity.
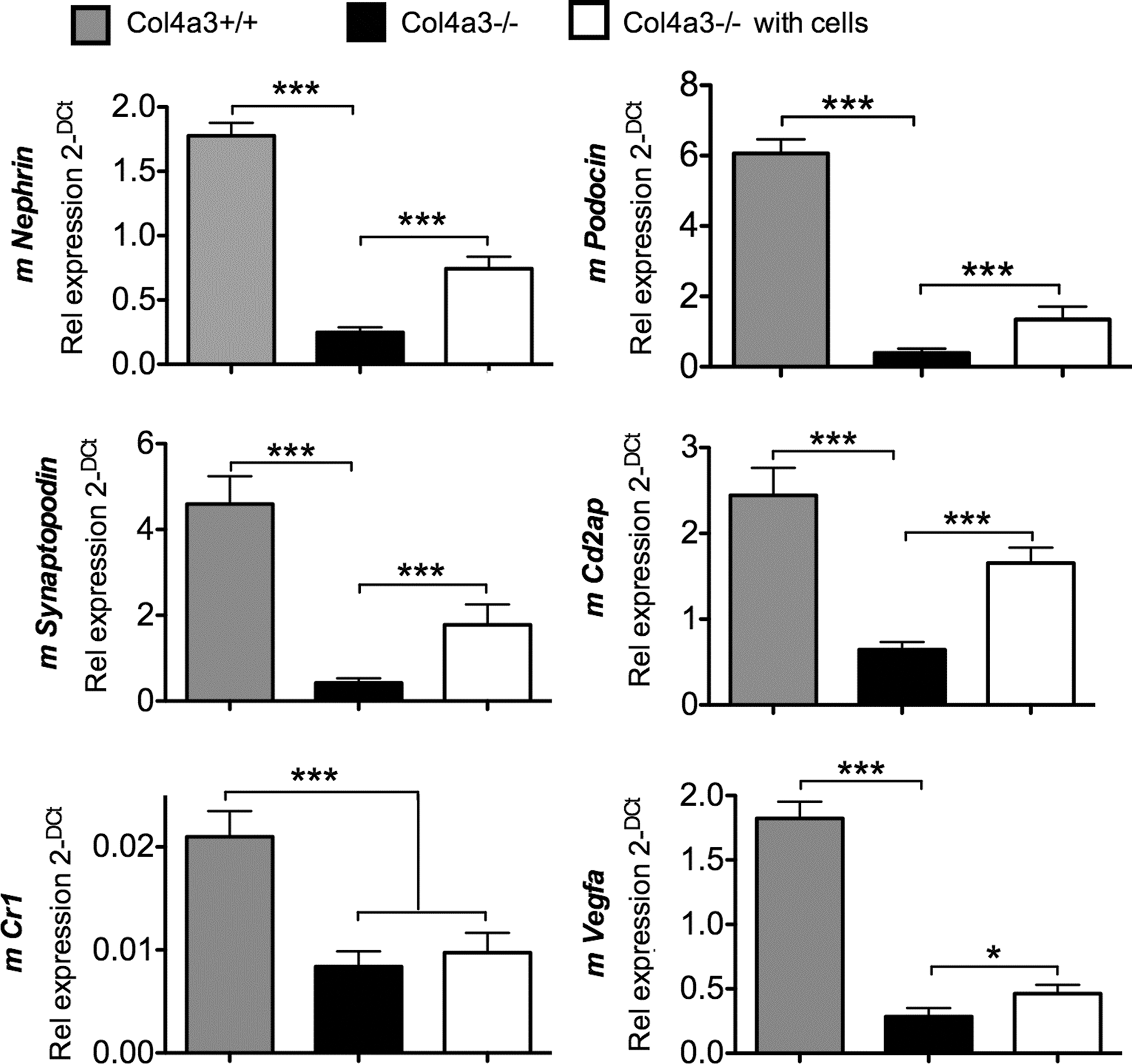
Expression of endogenous murine renal markers is modulated by CSC transplantation. QRT-PCR showing expression of podocyte markers using mouse-specific primers that do not amplify human sequences (Nephrin, Podocin, Vegfa, Synaptopodin, Cd2ap, Cr1), in the glomeruli of nontransplanted +/+ mice, nontransplanted −/− mice, and transplanted −/− mice. Values were normalized to mouse-specific cyclophilin. ***P < 0.001, *P < 0.05, n = 8 per group, error bars are SEM.
Discussion
Several studies have suggested that renal pathology in models of AS can be improved by cell therapy, although the mechanisms mediating these effects remain elusive, with conflicting results possibly attributable to variations in donor cell types. It is therefore essential to identify easily accessible sources of stem cells with high therapeutic potential for the treatment of AS [13]. In this study, we used human first-trimester fetal CSC, which are isolated from chorionic villi sampling in ongoing pregnancies and can be expanded to high numbers ex vivo, while maintaining tissue repair potential 10 . For example, when transplanted into collagen type I-deficient mice, they reduced fracture rate and increased bone plasticity and accelerated skin wound healing [10,15].
Col4α3−/− mice are a model of severe human AS. At 7 weeks of age, −/− Alport mice show high levels of proteinuria, but normal weight and blood urea. Over the next 2 weeks, blood urea rapidly increases, while body weight drops and mice show pronounced interstitial fibrosis and macrophage infiltration. We investigated the capacity of CSC to prevent glomerulopathy in 129Sv-Col4α3−/− mice. We show that 9-week-old Col4α3 −/− mice transplanted with CSC 2 weeks before have significantly lower blood urea and urine proteinuria, compared to their nontransplanted counterparts. Although animal welfare restrictions prevented us from studying the clinical endpoint of survival, all transplanted Col4α3 −/− mice maintained their weight until 9 weeks of age. This is important because improvement in renal histology is not necessarily associated with delay in death from renal failure [16].
However, the genetic background of the Alport mice has a strong effect on the rate of disease progression [17]. Contrary to 129Sv-Col4α3tm1Dec/J (−/−) mice, which progressively lose weight and do not survive beyond 10 weeks of age, the survival time of homozygous mutant mice is extended to about 14 weeks of age in mice maintained on a mixed genetic background or to 25–30 weeks on the C57BL/6j background. Consequently, we suggest that the elevated blood urea and 20% weight loss we report may be considered surrogates for end-stage renal failure.
Increasing evidence from stem cell transplantation in acquired injury models points to the well-characterized anti-inflammatory actions of exogenous stem cells making a major contribution to therapeutic results. For example, murine amniotic fluid cells transplanted into Col4α5−/− mice before the onset of proteinuria have been reported to modify the course of renal fibrosis, despite donor cells failing to differentiate into podocytes or produce collagen IVα5 4 . In this study, we found reduced renal fibrosis and cortical inflammation in transplanted mice, which may reflect the anti-inflammatory effect of donor cells or the replacement of defective renal cells. We also found that CSC migrated to the glomeruli, where they persisted over 2 weeks and expressed CR1, VEGFA, SYNAPTOPODIN, CD2AP, and PODOCIN at the gene level, and produced PODOCIN, CD2AP, and COLIVα3 proteins.
These data indicate that transplanted CSC have adopted a podocyte phenotype. However, focal staining for Col4a3 in the kidneys does not prove true GBM deposition, and it will be necessary to demonstrate assembly of the correct collagen type IV in the GBM to investigate whether the Col4a3 produced by CSC coassemble with Col4a4 and Col4a5 to improve GBM structure [18]. Similar to our findings, LeBleu et al. found that wild-type bone marrow-derived cells transplanted into Col4α3−/− mice improved renal histology and function, with donor cells differentiating into VEGF and collagen IV-expressing podocytes [9]; and a recent study by Lin et al. demonstrates that secretion of α3α4α5(IV) heterotrimers is sufficient to slow disease progression by partially restoring the defective collagen network [19].
Interestingly, we also found that CSC transplantation stimulated resident podocyte activity, suggesting that the production of Col4α3 from donor cells acts as a feedback to modulate podocyte activity, possibly by releasing trophic factors that promote the differentiation and regeneration of endogenous podocyte progenitors to differentiate into mature podocytes. Although endogenous podocytes remain unable to express the correct form of collagen type IV, stimulation of podocyte progenitor differentiation into podocytes by donor stem cells may contribute to the amelioration of filtration function [20]. Although it is often assumed that the presence of exogenous cells at the site of injury and their differentiation into target cell phenotypes account for the therapeutic effects observed, there is still a lack of evidence for the causality between the two.
An increasing number of studies challenge the concept of donor cells acting as building blocks to replace damaged endogenous cells and data suggest that beyond their potential as a source of cell replacement, stem cells also mediate paracrine treatment. In addition, data suggest that donor cells influence the complex cross-talk between resident cells and extracellular matrix. It is possible that exogenous stem cells reprogramme resident macrophages from an anti-inflammatory to a proinflammatory phenotype, as is the case in sepsis [21]. This mechanism might account for the therapeutic effects of wild-type bone marrow that we previously reported 7 . For example, blockade of tumor necrosis factor-alpha, a proinflammatory cytokine, has been shown to ameliorate glomerulosclerosis and proteinuria in AS mice [22].
We believe that CSC may have strong potential for the treatment of glomerulopathies and further studies are indicated to establish the precise mechanism of action of these cells in treatment of AS.
Footnotes
Acknowledgments
This research was funded by Kidney Research UK, Genzyme Renal Innovations Programme, and by the National Institute for Health Research Biomedical Research Centre at Great Ormond Street Hospital for Children NHS Foundation Trust and University College London. A.L.D. is funded by Department of Health through NIHR UCL/UCLH Biomedical Research Centre. We acknowledge support from NIHR Imperial Biomedical Research Centre.
Author Disclosure Statement
No competing financial interests exist.