
Abstract
Hematopoietic stem cell (HSC) engraftment has been achieved using single-cell transplantation of prospectively highly purified adult HSC populations. However, bulk transplants are still performed when assessing the HSC potential of early embryonic hematopoietic tissues such as the aorta-gonad mesonephros (AGM) due to very low HSC activity content early in development. Intra-bone marrow transplantation (IBMT) has emerged as a superior administration route over intravenous (IV) transplantation for assessing the reconstituting ability of human HSCs in the xenotransplant setting since it bypasses the requirement for homing to the BM. In this study, we compared the ability of IBMT and IV administration of embryonic day 11.5 AGM-derived cells to reconstitute the hematopoietic system of myeloablated recipients. IBMT resulted in higher levels of AGM HSC long-term multilineage engraftment in the peripheral blood, BM, spleen, and thymus of primary and secondary recipients, and in limiting dilution experiments. The administration route did not skew the multilineage contribution pattern, but IBMT conferred higher Lineage−Sca-1+c-kit+ long-term engraftment, in line with the superior IBMT reconstitution. Therefore, IBMT represents a superior administration route to detect HSC activity from developmentally early sources with limited HSC activity content, such as the AGM.
Introduction
H
In the murine and human settings, the transplantation assay is the in vivo gold standard test to study HSC function. HSC transplantation is widely used in experimental and clinical hematology, and refinement of HSC transplantation assays has resulted in improved HSC engraftment/readout. For instance, murine HSC transplantation is now optimized to be performed at the single-cell level and has been crucial in revealing the functional heterogeneity of murine and human HSC compartments. In the mouse, HSC transplantation assays are mainly performed by intravenous (IV) administration due to the intrinsic circulatory properties of hematopoietic cells. Successful engraftment then requires blood circulation and migration of HSCs into the BM, which offers a supportive niche. Alternatively, direct injection of cells within the BM has been successfully used to engraft human HSCs [5 –8], leukemia cells [9,10], and mesenchymal sarcoma cells [11,12] in immunodeficient mice. Intra-BM transplantation (IBMT) has shed light on the homing and repopulation properties of human HSCs and has been shown to result in higher levels of multilineage long-term reconstitution than IV injection. IBMT also allows detection of HSC potential from nonhematopoietic tissues, such as the liver, muscle, and human pluripotent stem cells. Interestingly, IBMT has also been used with whole murine BM cells [13,14]; however, in one study IBMT did not result in an enhanced engraftment compared with the IV technique [15].
Recently, the group of Daley tested the impact of HSC ontogeny (embryonic vs. adult) and the recipient's age (newborn or adult) on the engraftment potential upon transplantation [16]. They showed that embryonic HSCs from E9.5–10.5 preferentially engrafted neonatal mice. Despite the wide use of IBMT in the HSC xenotransplantation setting, no studies so far have assessed the engraftment ability of murine hematopoietic cells of embryonic origin by these different administration routes. We hypothesized that IBMT could represent a superior strategy to detect/readout hematopoietic reconstitution of embryonic cells, given their scarcity at early stages of development. Indeed, limiting dilution assays have reported that an AGM at E11.5 contains approximately one definitive HSC [17]. In this study, we have compared the ability of IV and IBMT administration of E11.5 AGM cells to reconstitute the hematopoiesis in a syngeneic transplantation setting. Our data show that IBMT confers superior multilineage engraftment of murine AGM cells over IV transplantation, thus offering an improved readout to study early developmental hematopoiesis function.
Materials and Methods
Mice and embryo dissection
C57BL/6J (CD45.2) and B6/SJL (CD45.1) mice were obtained from Charles River Laboratories. Both strains were housed and maintained at the Animal Facility of the Barcelona Biomedical Research Park (PRBB) following the recommendations of the Federation of European Laboratory Animal Science Associations. Timed matings were set up late in the evening, and vaginal plugs were checked in the early morning. The day of positive plug detection was considered E0.5. At E11.5, pregnant C57/BL6 females were sacrificed by cervical dislocation. The uterus was collected in phosphate-buffered saline (PBS), and embryo dissection was performed in PBS + 10% fetal calf serum (FCS) under a stereomicroscope. AGMs were dissected from each embryo, pooled and incubated for 30 min at 37°C in PBS + 10% FCS with 400 μg/mL collagenase type I. The cells were then washed four times with PBS + 10% FCS before transplantation. All experimental procedures were approved by the PRBB and Generalitat de Catalunya animal ethics committee.
Transplantation
Before transplantation, IBMT recipients were analgesized subcutaneously (2 mg/kg meloxicam and 0.1 mg/kg buprenorphine) and anesthesized by 4% isofluorane inhalation. For IBMT injections, the femur–tibia joint was perforated with a 25G needle, and subsequently, cells were injected into the retrieved orifice with a 27G needle in a volume of 25 μL [5,18]. IV injections were performed in the lateral tail vein in a volume of 200 μL [5]. One fresh AGM embryo equivalent (ee) CD45.2 donor cells were either IV or IBMT injected along with 400,000 unfractionated adult BM cells (CD45.1) into lethally irradiated (9 Gy) CD45.1 mice (n = 12 IV, n = 10 IBMT from n = 4 independent experiments). Transplanted animals received antibiotics in drinking water for 4 weeks after transplantation. Donor chimerism in peripheral blood (PB) was assessed at 3, 8, 12 (data not shown), and 16 weeks posttransplantation. Mice with ≥1% CD45.2+ cells in PB and multilineage (myeloid and B and T cell) reconstitution at 16 weeks posttransplantation were scored as reconstituted and considered for further analysis. Recipients were sacrificed at week 18 after transplantation for tissue analysis. For secondary transplantation, unfractionated BM cells (6 × 106) from IV- and IBMT-engrafted primary recipients were IV injected as described in lethally irradiated secondary CD45.1 mice (n = 2 secondary recipients per primary recipient; total n = 12). CD45.2+ donor engraftment was assessed at 3, 9, 16, and 20 weeks posttransplantation. As an AGM-limiting cell dose approach, 1 and 0.5 AGM ee CD45.2+ cells were IV or IBMT cotransplanted with 20,000 adult unfractionated BM CD45.1+ support cells into lethally irradiated CD45.1 mice. Mice with ≥1% CD45.2+ cells were considered donor engrafted. A total of 10 recipients were transplanted for each group in n = 5 independent experiments.
Donor chimerism analysis in PB and tissues
PB was collected from the facial vein directly into EDTA tubes. Red blood cell (RBC) lysis was performed using Quicklysis (Cytognos, Salamanca, Spain). Cells were then stained in PBS + 5% FCS with fluorochrome-conjugated antibodies. CD45.1-APC (allophycocyanin) and CD45.2-APC-Cy7 were used for allotype discrimination in transplanted mice. In multilineage analysis, cells were stained with Mac-1-V450, B220-PE-Cy7, and CD4/CD8-PE for myeloid, B-cell, and T-cell analysis, respectively. Cells were analyzed using a FACSCanto II cytometer (BD Bioscience) and FlowJo software (Tree Star).
For BM analysis, femurs and tibiae were harvested and pooled from IV recipients. For IBMT recipients, injected tibiae were harvested separately from the remaining femur and from bones of the contralateral limb. A BM cell suspension was obtained by crushing the bones, and RBC lysis was performed before staining. For Lineage−Sca-1+c-kit+ (LSK) staining, cells were incubated with the following antibodies: lineage cocktail (comprising Mac-1, Gr-1, B220, CD4, CD8a, CD5, Ter-119, all directly conjugated to PE-Cy5), c-kit-PE, Sca-1-FITC, CD45.1-APC, and CD45.2-APC-Cy7. For spleen and thymus analysis, single-cell suspensions were obtained by dissociating the tissue using a 70-μm cell strainer and a syringe plunger. After RBC lysis, splenic cells were stained with B220-VioBlue, CD8-FITC, CD4-PE, CD45.1-APC, and CD45.2-APC-Cy7, whereas thymus cells were stained with CD8-FITC, CD4-PE, CD45.1-APC, and CD45.2-APC-Cy7. Further details on fluorescence-activated cell sorting (FACS) antibodies used are shown in Supplementary Table S1 (Supplementary Data are available online at
Statistical analysis
Data are expressed as mean ± SEM of independent experiments. Statistical comparisons were performed using either the paired or unpaired Student's t test as appropriate. Statistical significance was defined as P value <0.05. Kaplan–Meier plots representing donor engraftment-assisted survival were calculated using SPSS software [19].
Results
IBMT confers a superior multilineage engraftment of AGM cells over IV administration
AGM cells contribute to low levels of chimerism when transplanted into adult recipients [16] due to their low HSC frequency/potential. Because IBMT constitutes a superior administration route than IV in the xenograft setting, we compared side-by-side the repopulation ability of murine E11.5 AGM cells after IBMT or IV transplantation. One ee of E11.5 AGM cells (CD45.2) was transplanted along with support cells (CD45.1) into adult lethally irradiated CD45.1 recipients by IV or IBMT (Fig. 1A). Analysis of PB chimerism at 16 weeks posttransplantation in reconstituted mice showed that IBMT-transplanted mice exhibited almost a two-fold higher level of donor reconstitution compared with IV-transplanted mice (79% ± 11% vs. 46% ± 12%; P = 0.1, Fig. 1B). Regardless of the administration route, a donor multilineage (myeloid, B-cell and T-cell) engraftment was consistently observed (Fig. 1C). The increased engraftment observed in PB of IBMT-transplanted mice was accompanied by enhanced donor reconstitution in other hematopoietic tissues at 18 weeks posttransplantation (BM: 54% ± 7% vs. 28% ± 10%; spleen: 79% ± 10% vs. 49% ± 17%; thymus: 90% ± 9% vs. 47% ± 20%, IBMT vs. IV, respectively. P ≤ 0.05 for pooled tissues, Fig. 1D).
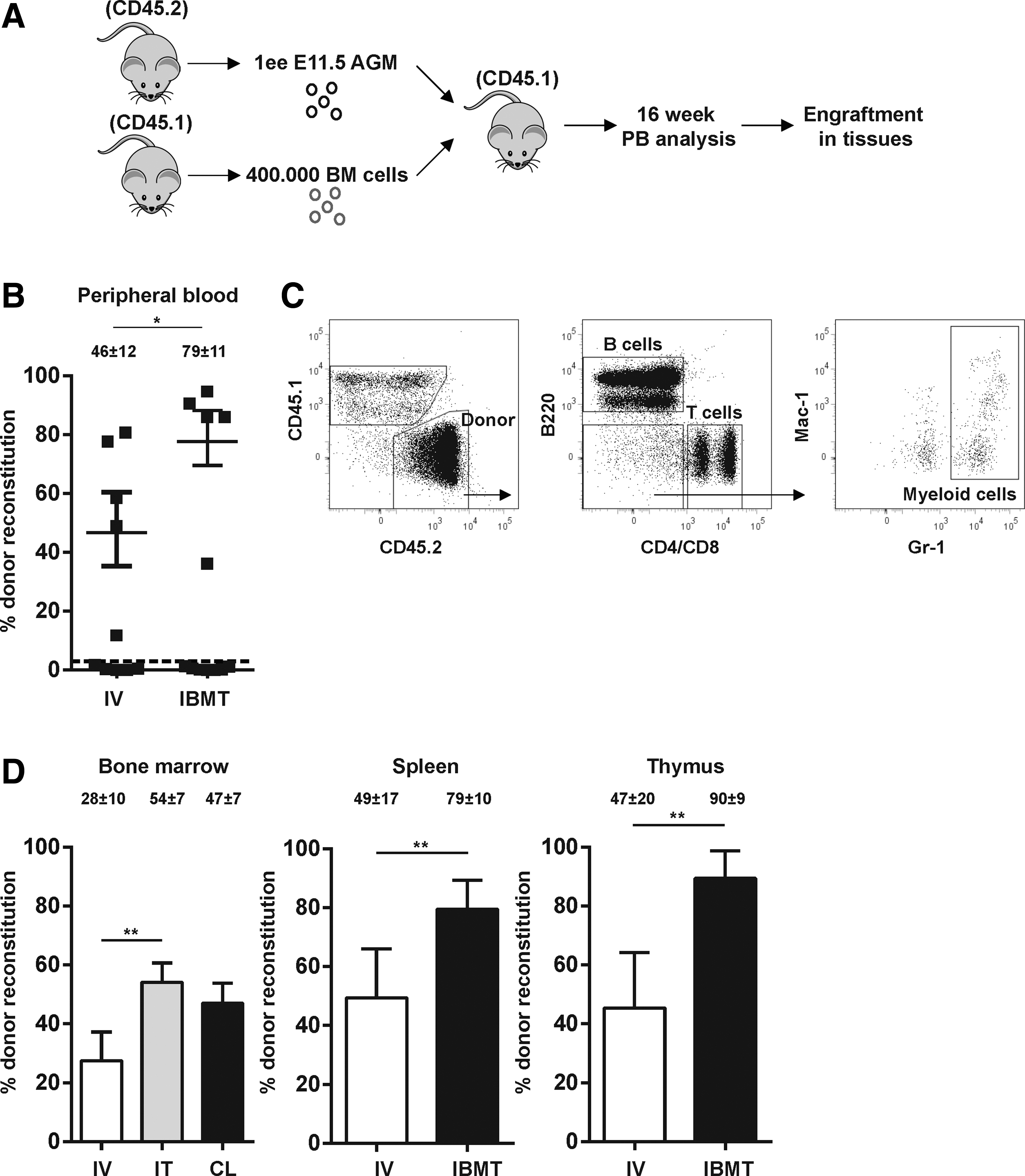
Intra-bone marrow transplantation (IBMT) confers a superior multilineage engraftment of aorta-gonad mesonephros (AGM) cells over intravenous (IV) administration.
The IBMT administration route did not skew the reconstitution pattern of the donor-engrafted cells since a similar percentage of myeloid (Gr-1+), B (B220+), and T (CD4+/CD8+) cells were found within the CD45.2+ graft in the BM, spleen, and thymus (Fig. 2A, B). However, the percentage of donor cells within the LSK compartment, which comprises the pool of immature stem and progenitor cells in the mouse hematopoietic system [20], was 50%–60% higher upon IBMT (Fig. 2C, left panel). Similarly, the percentages of CD45.2+ within the Sca-1+c-kit− and c-kit+Sca-1− compartments were higher in IBMT over IV-transplanted mice (Fig. 2C). Together, our data suggest that injection of AGM cells directly into the BM constitutes a more robust multilineage reconstitution approach than IV transplantation.
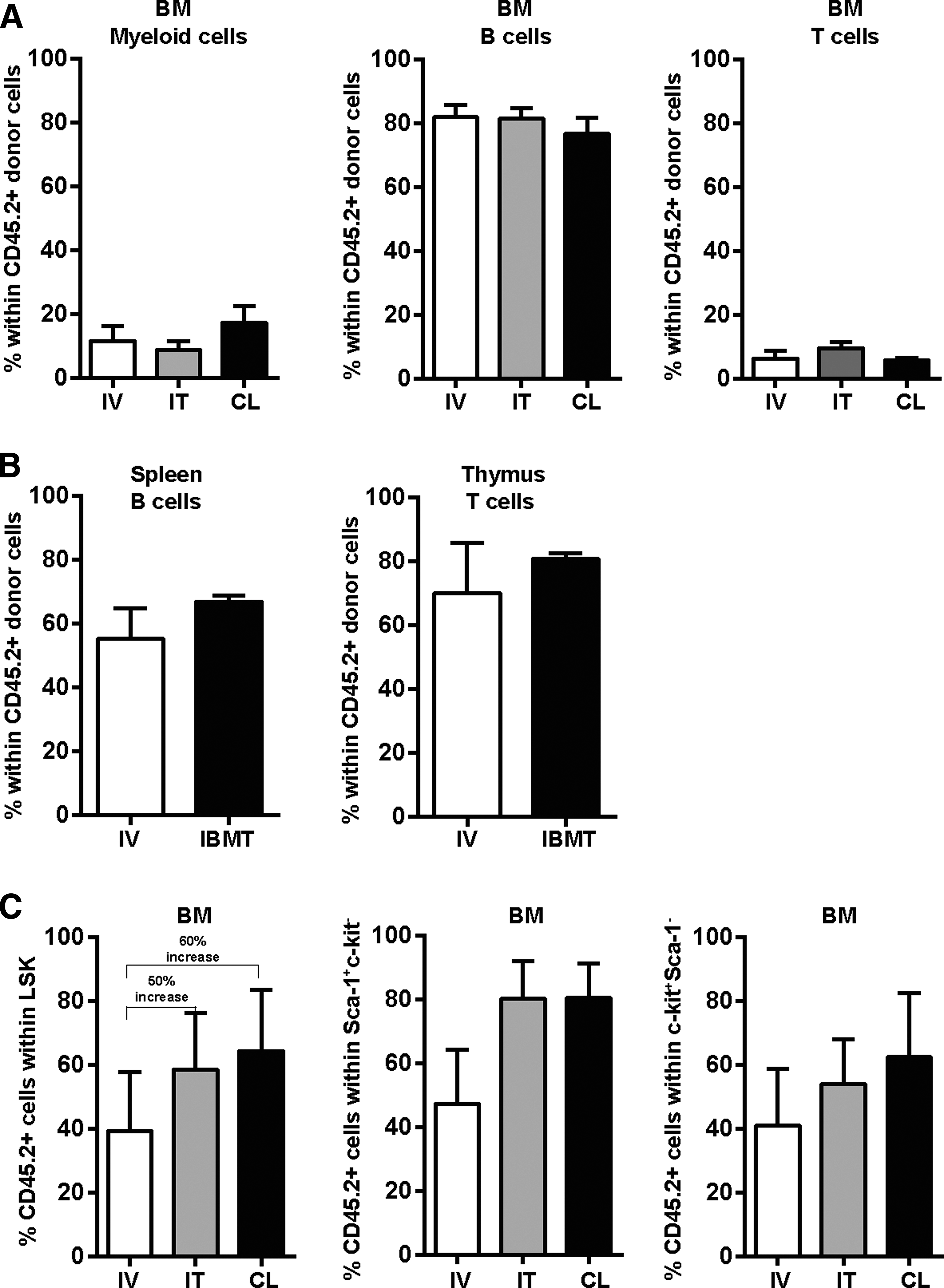
Multilineage contribution pattern in different hematopoietic tissues is not skewed by the administration route.
IBMT confers higher hematopoietic reconstitution in secondary recipients and in limiting dilution experiments
To prove bona fide E11.5 AGM-derived HSC self-renewal and multilineage differentiation, primary IV and IBMT grafts (6 million unfractionated BM cells from 18-week primary recipients) were serially IV-transplanted into secondary recipients [21] (Fig. 3A). Hematopoietic reconstitution was assessed in the PB of secondary recipients at different time points after transplantation. Primary IBMT grafts resulted in consistently higher PB chimerism levels in secondary recipients compared with primary IV grafts (65%–90% vs. 38%–52%, P ≤ 0.05, Fig. 3B). Similarly, a higher multilineage donor chimerism was found after 20 weeks in PB, BM, spleen, and thymus of secondary recipients transplanted with primary IBMT-engrafted cells compared with primary IV grafts (∼90% vs. ∼60%. P ≤ 0.05 for pooled tissues, Fig. 3C). The percentages of donor cells within the immature hematopoietic compartment (LSK), more primitive hematopoietic progenitors (Sca-1+c-kit−) or progenitor cells (c-kit+Sca-1−), was higher in IBMT over IV secondary recipients (Fig. 3D). We then assessed whether IBMT administration facilitates the engraftment of lower doses of AGM cells (Fig. 3E). IBMT-transplanted 1 ee and 0.5 ee conferred 40% and 23%, respectively, multilineage hematopoietic engraftment-assisted 20-week survival. In contrast, equivalent AGM doses IV- transplanted conferred 25% and 7% engraftment, respectively, after 20 weeks (Fig. 3F). These results confirm that IBMT administration facilitates the readout of hematopoietic reconstitution of embryonic tissues with very low HSC content.
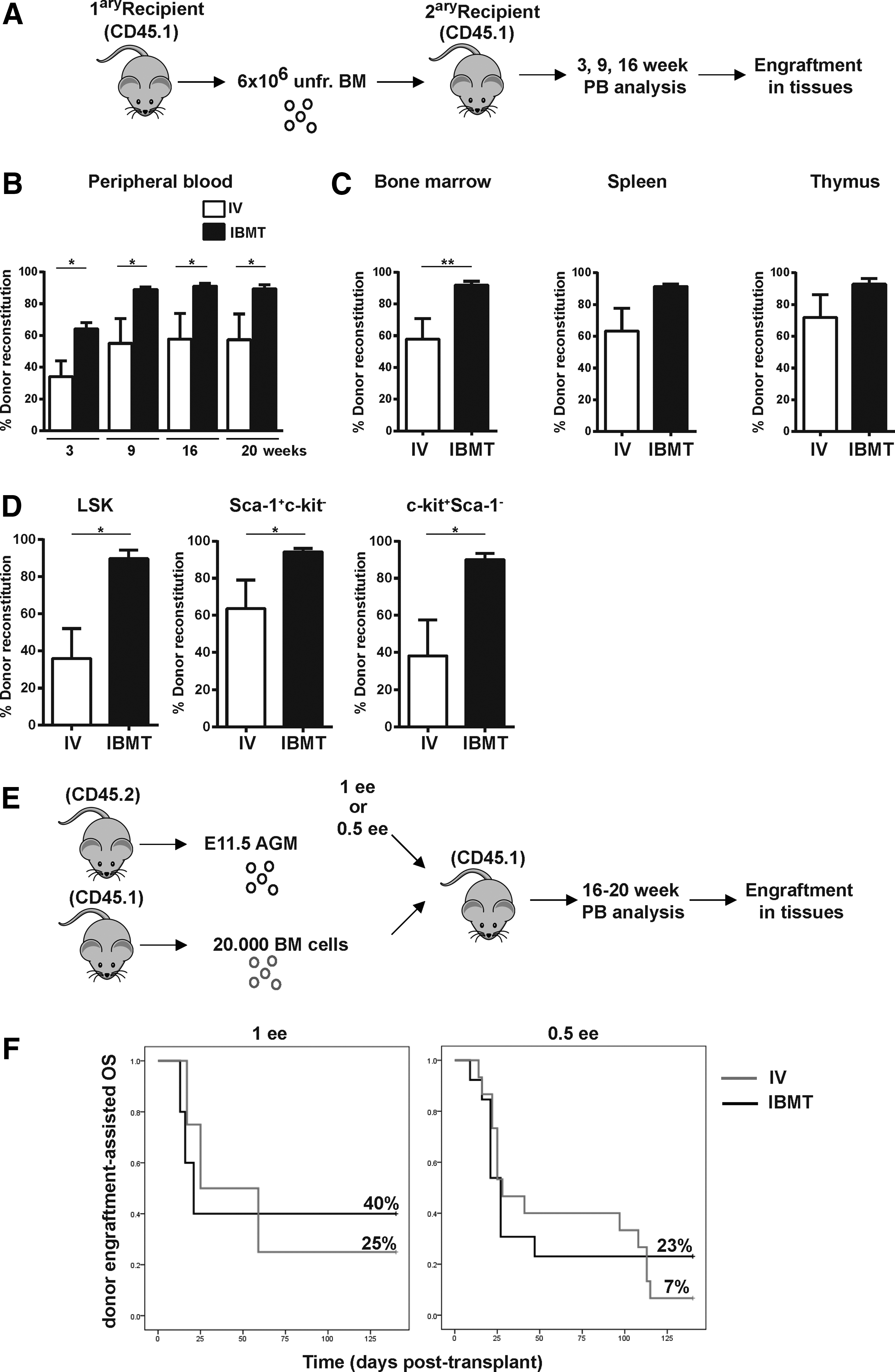
IBMT confers higher donor-derived reconstitution in secondary recipients and in limiting dilution experiments.
Discussion
Early in embryogenesis, hematopoietic stem and progenitor cells (HSPCs), such as those derived from yolk sac, AGM, or embryonic stem cells (ESCs), are very poorly represented due to either their scarcity or low developmental potential [17,22]. Definitive HSCs emerge from the AGM, and AGM-derived HSCs are the first cells reconstituting hematopoiesis in an irradiated adult animal [23]. Assessing the hematopoietic potential and plasticity of early AGM-derived cells relies on robust and sensitive in vivo long-term functional assays, and transplantation into irradiated mice is considered the in vivo gold standard test of HSC functionality. Although HSPC transplantation assays are commonly performed by IV administration, direct injection of cells into the BM is an alternative modality that has been used successfully to engraft human HSCs [5 –7]. IBMT-based HSC delivery is, however, less common in the mouse syngeneic setting.
In this study, we compared the ability of IBMT and IV administration of AGM-derived cells to reconstitute the hematopoietic system of myeloablated recipients. In contrast to the IV route, IBMT administration resulted in significantly higher levels of AGM HSC long-term multilineage engraftment in all analyzed hematopoietic tissues of primary and secondary recipients. Importantly, the multilineage contribution pattern was not skewed in any hematopoietic organ by the IBMT administration route, suggesting that IBMT does not impair in vivo HSPC differentiation and migratory properties. However, IBMT conferred higher LSK long-term engraftment, in line with the superior IBMT reconstitution. Importantly, multilineage reconstitution was barely observed after 20 weeks when 0.5 ee was IV transplanted, but was detected in 23% of the IBMT-transplanted mice receiving 0.5 ee. Our study indicates that IBMT represents a superior administration route to detect HSC activity for many applications, including when a limited number of HSCs are available from developmentally early sources such as the AGM, and when studying BM niche-HSC interactions [24]. We envision that the potential advantage of IBMT over IV might be as follows: (1) it bypasses the requirement for homing to the BM niche; (2) it facilitates survival within the niche; and (3) it prevents transplanted cells from being taken up by organs in vivo as occurs with intravenously injected cells.
Recently, Daley and coworkers' demonstrated that recipient age influences the engraftment of embryonic HSCs [16] by showing that IV injection of AGM cells into neonatal recipients yielded a more robust engraftment than in adult recipients. Another study demonstrated that intrahepatic transplantation of human CB-CD34+ cells into newborn immunodeficient mice provided enhanced hematopoietic engraftment without the requirement of accessory cells [25]. Our data suggest that IBMT of AGM cells in adult recipients may be used as an alternative modality to overcome the logistics of matching recipient's age with the nature of donor cells. The group of Ikehara has also compared IBMT and IV transplantation in mouse models. They showed that IBMT constitutes a powerful strategy in several scenarios, including treatment for autoimmune diseases [26]. Similarly, Li et al. analyzed the kinetics of hematopoietic reconstitution very early after IBMT or IV transplantation and found more donor-derived cells in IBMT-transplanted animals [14]. They also analyzed the long-term maintenance of donor-derived hematopoiesis after IBMT or IV using serial transplantation and found that the frequency of donor-derived HSPCs was higher in IBMT- than in IV-transplanted tertiary recipients [13]. In addition, Baba et al. used serial transplantation assays to demonstrate how IBMT accelerates donor cell engraftment and proliferation and also facilitates reconstitution of dendritic cells [27]. In contrast to our results, de Haan's group showed that IBMT in a murine syngeneic setting does not provide enhanced engraftment in comparison with IV administration [15]. A likely explanation for this discrepancy may be the different ontogeny stages of donor cells, since adult BM cells with a considerably higher HSC potential were used in the de Haan study.
Whether IBMT is superior to IV in terms of engraftment for other embryonic tissues, such as yolk sac, placenta, FL, or ESC-derived cells, remains unknown. Likewise, the engraftment potential of IBMT-transplanted ex vivo expanded AGM cells needs to be investigated in future studies. In the clinical setting, several trials have assessed the benefit of allogeneic transplantation using IV or IBMT administration [28]. One study observed reduced risk and severity of acute graft-versus-host disease in the IBMT cohort. Moreover, a 20-year follow-up trial concluded that intraosseous infusion of BM cells was a safe and efficient procedure [29]. Thus, we envision that IBMT will become the administration route of choice in experimental and clinical hematology.
Footnotes
Acknowledgments
This work was supported by the European Research Council CoG to P.M., the Spanish Ministry of Economy and Competitiveness (ISCIII/FEDER PI14/01119 to C.B. and SAF2013-43065R to P.M.), the Spanish Association Against Cancer to P.M. and C.B., the Fundación Inocente Inocente to P.M., a Marie Curie Career Integration Grant to A.S.-P. (FP7-PEOPLE-2013-CIG-631171), and the José Carreras Leukämie-Stiftung to P.M. and A.S.P. C.B. is supported by a Miguel Servet II contract (CPII13/00011). C.P. is supported by PFIS scholarship (FI12/00468). P.M. also acknowledges the financial support from the Obra Social La Caixa-Fundació Josep Carreras and the Generalitat de Catalunya (SGR330).
Author Disclosure Statement
The authors have no conflict of interest to disclose. All authors have read and approved the article in its present form.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.