
Abstract
The central nervous system has a limited capacity to regenerate, and thus, traumatic injuries or diseases often have devastating consequences. Therefore, there is a distinct need to develop alternative treatments that can achieve functional recovery without side effects currently observed with some pharmacological treatments. Combining biomaterials with pluripotent stem cells (PSCs), either embryonic or induced, has the potential to revolutionize the treatment of neurodegenerative diseases and traumatic injuries. Biomaterials can mimic the extracellular matrix and present a myriad of relevant biochemical cues through rational design or further functionalization. Biomaterials such as nanofibers and hydrogels, including self-assembling peptide (SAP) hydrogels can provide a superior cell culture environment. When these materials are then combined with PSCs, more accurate drug screening and disease modeling could be developed, and the generation of large number of cells with the appropriate phenotype can be achieved, for subsequent use in vitro. Biomaterials have also been shown to support endogenous cell growth after implantation, and, in particular, hydrogels and SAPs have effectively acted as cell delivery vehicles, increasing cell survival after transplantation. Few studies are yet to fully exploit the combination of PSCs and innovative biomaterials; however, initial studies with neural stem cells, for example, are promising, and, hence, such a combination for use in vitro and in vivo is an exciting new direction for the field of neural regeneration.
Introduction
Central nervous system damage
D
To overcome CNS injury or degeneration, a multifaceted approach is likely to be required, primarily to emulate desirable physical and biochemical characteristics of the healthy extracellular matrix (ECM). This approach can improve current cell culture environments, and improve cell transplantation outcomes in vivo by presenting a benign, supportive microenvironment for enhanced regeneration. Such requirements to provide both the physical and biochemical cues found in vivo can be fulfilled with the use of biomaterials.
The benefits of biomaterials in CNS regeneration
Many in vitro and in vivo approaches have been investigated to improve our understanding of disease and achieve neural regeneration. However, an inability to effectively replicate the biological environment has hampered their success.
In the native in vivo environment, the ECM, composed of proteoglycans, proteins, and signaling molecules, provides structural and biochemical support to residing cells. It has the ability to influence cell behavior, including differentiation, proliferation, survival, and migration [2,3]. Biomaterials possess favorable properties that can be engineered to provide the necessary mechanical and biochemical signaling to cells similar to that seen in the native cellular environment [1].
In vitro, gold standard two-dimensional (2D) cell culture systems are used to model diseases and produce large number of cells for drug screening or transplantation. Currently, however, the inherent limitation of these systems is an inability to fully mimic the natural three-dimensional (3D) environment and to thus be effective when translated in vivo. Similarly, the transplantation of exogenous cells in the CNS to promote neural regeneration has become a widely researched strategy to overcome the limitations of current treatments [4]. However, issues such as long-term graft integration and cell survival have limited the application of this strategy to achieve neural regeneration. To overcome such limitations, biomaterials such as electrospun nanofibers, hydrogels, and self-assembling peptides (SAPs) are being investigated to enhance current cell culture systems to improve the accuracy of disease modeling and drug screening and to support cell transplantation in vivo.
These materials have the ability to present ECM-like mechanical cues to cells, as hydrogels can be tailored to have a similar elastic modulus to the CNS tissue, whereas nanofibrous scaffolds and SAPs possess fibrillar structures on a length scale, representative of proteins found in the in vivo milieu. Chemical modification of these materials can also provide bioactive signals to cells, through growth factor presentation or the presentation of bioactive sequences, all of which have been shown to increase cell survival and proliferation, and, in some cases, direct cell differentiation [5 –9].
Biomaterials and stem cells
Appropriately designed biomaterials have the ability to achieve such complex structures and have the potential to significantly improve stem cell culture systems and improve cell transplantation outcomes for neural regeneration. By functionalizing these materials with niche-specific signals, there is an opportunity for cells to be produced in vitro that possess in vivo-like morphology. These cell culture systems can then be implemented in disease models and provide large number of cells for drug screening and transplantation. In addition, by designing biomaterials as carriers that can provide physical and biochemical support to cells during transplantation in vivo, it may be possible to synthetically fabricate a milieu that is conducive to neural regeneration and can promote functional recovery post-CNS injury or disease.
Three-Dimensional Biomaterials Can Provide Superior Cell Culture Environments
Limitations of current cell culture systems
Commonly used 2D cell culture conditions are limited in their ability to provide the complex physical and biochemical cues present in the in vivo environment that are essential to cell development, survival, and function [10]. The inherent heterogeneity of traditional cell culture systems and subsequent lack of consistent spatial or temporal gradients of mechanical or biochemical signals, as well as the absence of 3D cell–cell and cell–matrix interactions, can result in significant alterations in gene and protein expression, cell morphology, and function [11 –13]. These limitations can compromise the performance of these cells for further in vitro experimentation and future in vivo deployment, so it is imperative to develop systems that accurately replicate relevant in vivo environments for culture [14].
Significantly, cell culture systems that only provide insufficient 2D support to cells affect the accuracy of further in-depth investigations of cells and their behavior in vitro, including disease modeling and drug screening. As cells generated and maintained in vitro are exposed to different physical and biochemical cues than their in vivo counterparts, findings from in vitro experimentation are not always accurate representations of cellular development and behavior in their natural environment, and can, therefore, be difficult to translate into in vivo experiments [11]. Reproducibility and scalability are also limitations of cell culture systems, because of the use of human or animal tissue, including nonquantified tumor-derived materials such as Matrigel [12]. These limitations are problematic when considering the large number, and in vivo-like nature, of cells required for transplantation, disease modeling, and drug screening, and illustrate the need for a superior cell culture system that can effectively recapitulate the 3D in vivo environment for neural regeneration applications and strategies.
Electrospun nanofibers for neural differentiation and proliferation in vitro
Electrospun nanofibers have been intensely researched for applications in neural regeneration because of morphological similarities to the ECM that include similar fibril diameter, high surface area-to-volume ratio, and high porosity [1,15]. Such similarities highlight their suitability for use in a 3D cell culture system.
Electrospinning uses a viscous polymer solution that is electrified by a high-voltage source. At a critical voltage, the electrostatic attraction between the polymer solution and collector becomes greater than the electrostatic repulsions between charges within the solution, resulting in the formation of a fiber jet from the polymer solution. This fiber experiences whipping instabilities, during which the solvent evaporates, resulting in the deposition of polymer fibers on the collector [15,16]. Sheets of nanofibers (scaffolds) can be collected after electrospinning and easily utilized in subsequent cell culture. Polymers such as poly(ɛ-caprolactone) (PCL), poly-
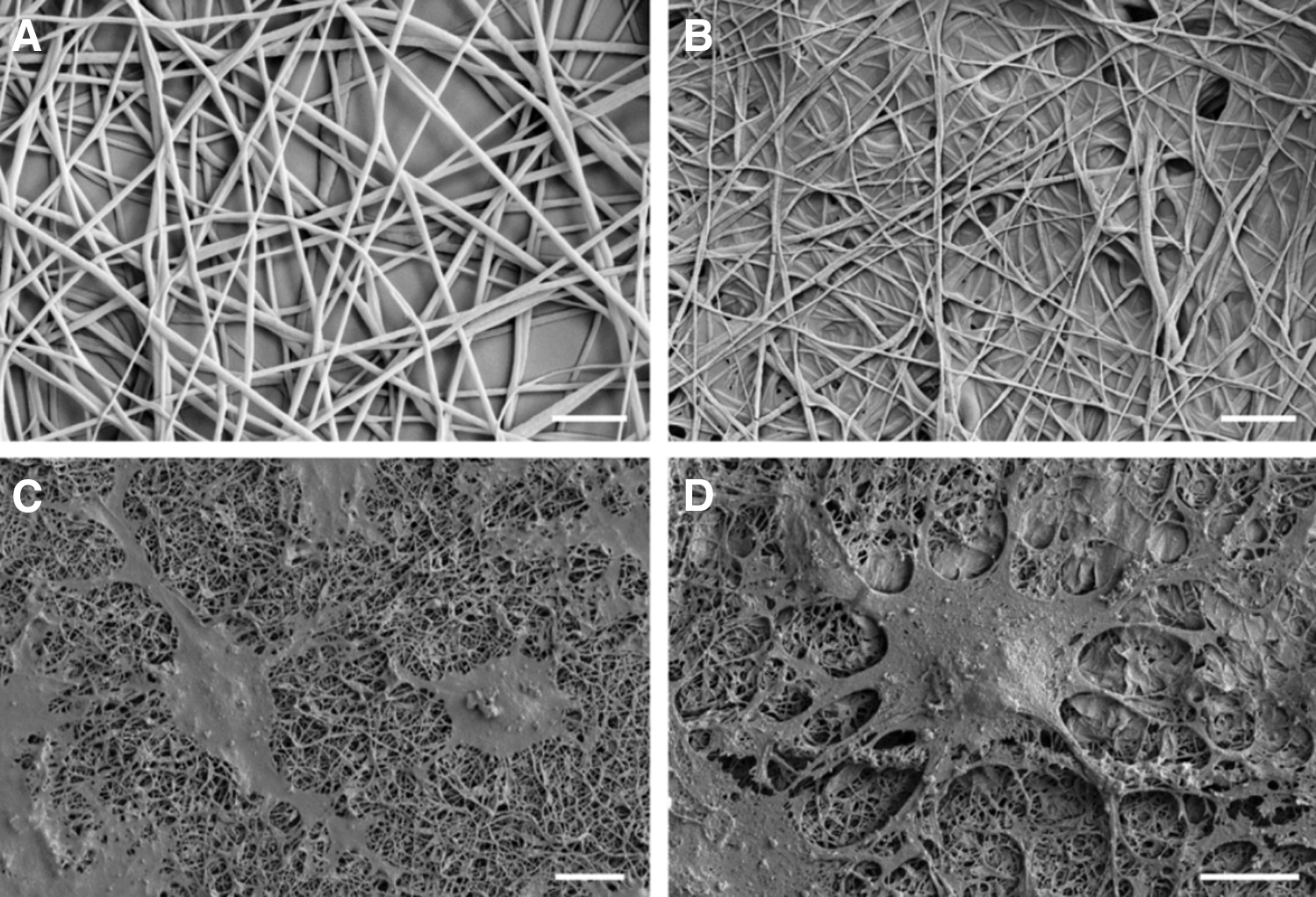
Field-emission scanning electron microscope images of
Electrospun nanofibers are an attractive biomaterial for a 3D cell culture system as the scaffolds can be tailored for different applications through fiber diameter, alignment, and surface modification [17]. Modification of nanofiber surfaces through chemical attachment of bioactive molecules allows for the presentation of biochemical cues necessary to create a cell culture system that effectively recapitulates the extracellular environment. For example, cell adhesion and spread were enhanced on PCL nanofiber scaffolds that were aminolyzed with ethylenediamine as a result of the increased hydrophilicity from the presentation of amine moieties, as shown in Fig. 2 [18]. In addition, neural stem cells (NSCs) primarily differentiated into oligodendrocytes when cultured on both control and aminolyzed PCL nanofiber scaffolds as compared to the laminin control [18], indicating that appropriately designed PCL nanofiber scaffolds have the ability to direct neural differentiation toward a specific lineage.
Cultured NSCs (labeled with nuclear marker Hoechst 3342,
Neural survival, proliferation, and differentiation can also be influenced through the presentation of proteins on the surface of electrospun scaffolds. For example, covalent attachment of brain-derived neurotrophic factor (BDNF) or glial cell-derived neurotrophic factor (GDNF) to aminolyzed PCL nanofibers significantly increased cell proliferation and survival compared to presentation of these proteins in their soluble forms [5,7,8].
The effect of ECM protein coatings on nanofibers has also been investigated. Laminin-coated polyurethane nanofibers have been shown to support complex morphology of astrocytes with long cellular extensions, representative of in vivo morphology, compared to the flat morphology observed when using the 2D laminin-coated glass cover slips (Fig. 3) [13]. The expression of intermediate filament proteins [glial fibrillary acidic protein (GFAP), vimentin, and nestin] and heat-shock protein (HSP70), a marker of cellular stress, was reduced in astrocyte cultures on laminin-coated nanofibers, compared to those cultured on laminin-coated glass slips [13]. In addition, cell cultures of mouse embryonic stem cells (ESCs) on Ultra-Web™, a commercially available polyamide-based 3D nanofibrillar porous matrix, have been shown to enhance cell proliferation and self-renewal compared to tissue culture dishes [19]. Culturing primary astrocytes on 3D PCL nanofiber scaffolds resulted in decreased GFAP expression, while increasing the expression of BDNF and excitatory amino acid transporter 2 (EAAT2) compared to those cultured on 2D PCL [20]. These biochemical changes are indicative of a phenotype that is representative of astrocytes in the healthy brain and demonstrate that the morphology of the culture environment is important in developing in vivo-like cell behavior and phenotype. Therefore, the ease in which the 3D morphology and surface chemistry of nanofibers can be altered is promising for influencing stem cell fate and thereby holds relevance for applications with regard to in vitro disease modeling, drug screening, and generation of cells suitable for in vivo clinical application.
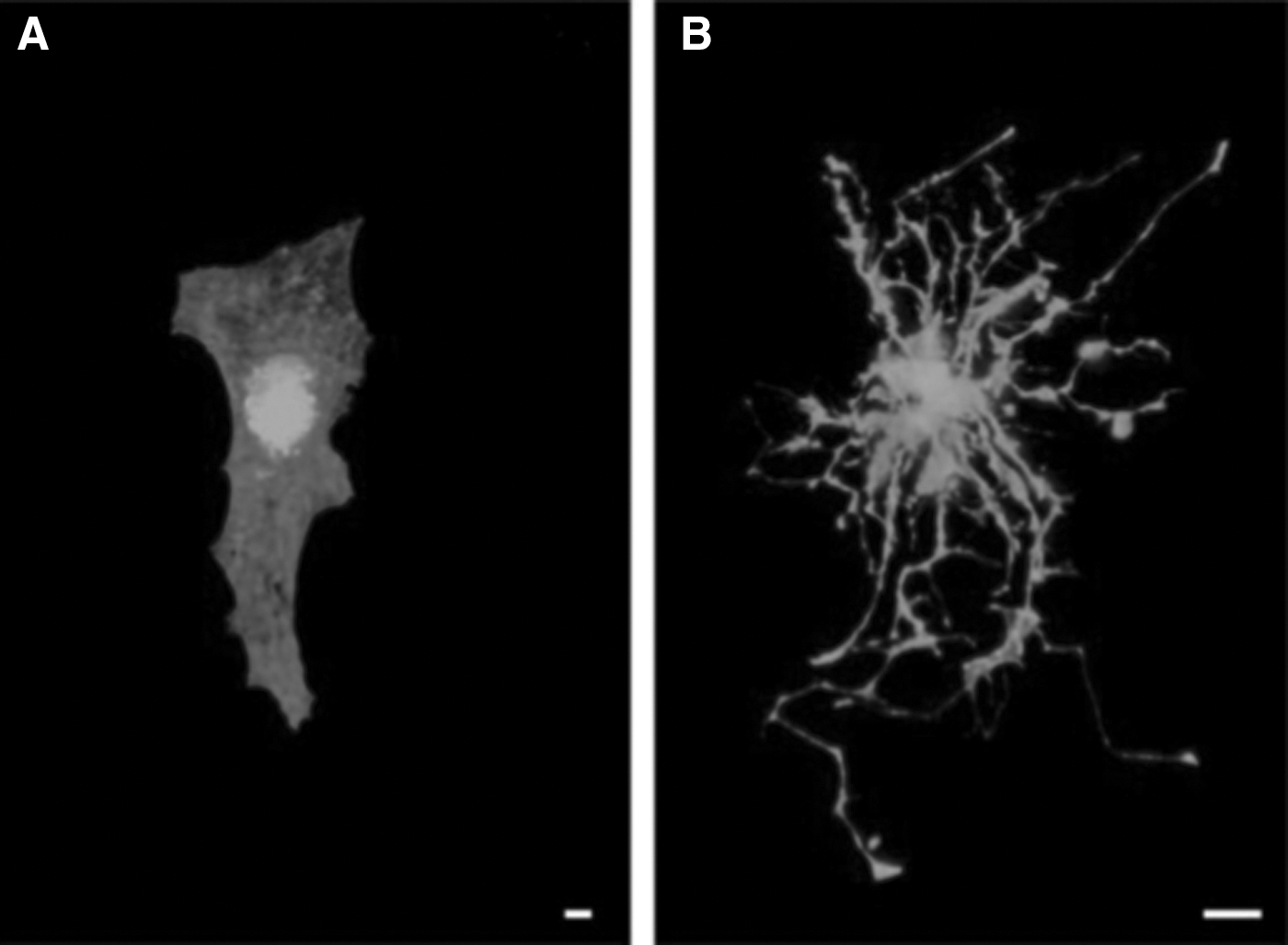
Cell morphology of enhanced green fluorescent protein (EGFP)-expressing astrocytes cultured on:
Potential of hydrogels for improved in vitro cell culture
An exciting method for translating the benefits of electrospun scaffolds to improve current cell culture systems for in vitro application is the development of hydrogels. These are highly hydrated, water-insoluble polymer networks that contain both chemical (covalent bonds) and physical (chain entanglement and secondary forces) cross-links [1,21]. They can be synthesized to possess numerous characteristics of the architecture and mechanics of the native cellular environment [22]. Many hydrogels have been investigated for neural regeneration applications and include biologically derived materials, such as hyaluronic acid (HA), collagen, xyloglucan, and Matrigel, as well as synthetically derived hydrogels such as polyacrylamide and polyethylene glycol (PEG) [22,23]. They are appealing biomaterials because of their high oxygen and nutrient permeabilities, as well as low interfacial tensions and macroporosity, facilitating cell infiltration and effective graft–cell interface for innervation of host circuitry [1,22 –24].
It is essential that mechanical properties of biomaterials for neural regeneration are similar to that of native brain tissue to ensure that cells receive the appropriate mechanical growth cues, while also avoiding modulus mismatch and a subsequent foreign body reaction if also implemented in vivo [23,25,26]. This highlights another desirable characteristic of hydrogels, as their mechanical properties can be easily manipulated to reflect that of the CNS by controlling the underlying structure (eg, the number of fibers and cross-links) [1]. Although proliferation of human ESCs (hESCs) and human-induced pluripotent stem cells (hiPSCs) is insensitive to ECM stiffness, early differentiation is regulated by ECM stiffness when substrate stiffness is similar to brain tissue. It has been shown that neural differentiation of hESCs and hiPSCs is increased on hydrogels with a stiffness of 0.1 kPa as compared to substrates with stiffness of 75 kPa [27]. A downstream increase in neurons was observed when hESCs and hiPSCs were initially exposed to soft (0.7 kPa) ECMs (for 5 days out of a total 19 days), indicating that mechanical signaling may also have temporal significance [27]. In addition, when neural stem/progenitor cells (NSPCs) were cultured on methylacrylamide chitosan hydrogel, it was found that cellular proliferation and neuronal differentiation were maximum when elasticity was <1.0 kPa. Oligodendrocyte maturation and myelination have been observed to be greater on scaffolds with E < 1.0 kPa, whereas stiffer substrates (E > 7 kPa) favored oligodendrocyte differentiation [25]. This demonstrates the varying effects that ECM stiffness can have on cell proliferation and differentiation and hence, the potential for biomaterials to create a myriad of tailored cell culture environments.
The significance of the mechanical properties of a potential scaffold highlights the suitability of hydrogels in cell culture systems, as their mechanical properties can easily be tuned through the regulation of cross-linking density [1]. However, limited biodegradation, shrinkage of hydrogels after cross-linking, and potentially toxic monomers and cross-linking agents are shortcomings that hinder the clinical application of synthetic hydrogels in neural regeneration [28]. Conventional hydrogels, although porous, lack the inherent fibrous morphology found in the ECM, which is an essential aspect in creating superior, 3D cell culture environments. As such, an alternative approach has been investigated to develop more sophisticated hydrogel structures that comprise fully degradable small molecules and possess fibrous morphology, namely SAPs.
SAPs provide tailored bioactivity in vitro
SAPs are an alternative hydrogel scaffold that can be used in 3D cell culture applications, as they have the capacity to self-assemble and form complex nanostructures from simple peptide building blocks without the limitations already described. The noncovalent self-assembly of peptide-based molecules enables the presentation of chemically functional peptide sequences at the surface of ordered nanostructures such as tubes, rods, and sheets, which can influence cell survival, proliferation, migration, and differentiation [29]. In addition, self-assembly can be controlled by pH, ionic strength, temperature, or enzymatic triggers [29]. Peptide-amphiphiles (PAs), aromatic N-terminally capped peptides, and proline-containing oligopeptides are examples of SAP systems that have been investigated for neural regeneration applications, as they mimic the ECM through the presentation of biochemical and physiochemical cues [28 –30].
Specific peptide sequences can be introduced to SAP systems to impart bioactivity and influence cell behavior. For example, the laminin-based epitope, isoleucine-lysine-valine-alanine-valine (IKVAV), known to promote neurite growth [31], has been incorporated into a variety of SAP systems, including RADA16 [32 –34]. RADA16 is a class of commercially available SAPs where self-assembly occurs due to complementary charge and hydrophobic interactions. Containing a 16 amino acid sequence with alternating charge, RADA16 self-assembles to form stable β-sheet structures [29,32]. The presentation of a bioactive laminin epitope, IKVAV, in a RADA16 scaffold has been found to promote neuronal differentiation, with an increased expression of the neuronal marker, MAP2, as well as a significantly higher percentage of neurons than astrocytes on the IKVAV-containing scaffold [9].
In addition, when encapsulated with NSCs, the bioactive SAPs ac-(RADA)4-GG-SKPPGTSS-CONH2, ac-(RADA)4-GG-PFSSTKT-CONH2, and ac-(RADA)4-GG-RGDS-CONH2 were found to have greater cell viability over 5 months than Matrigel and Collagen I [35]. This was attributed to the beneficial effects of the specific sequences SKPPGTSS, PFSSTKT, and RGDS on neural cell growth, migration, adhesion, and differentiation [35]. It was also found that after 3 months in culture, NSCs encapsulated in the peptide hydrogels differentiated into approximately 27% neurons, 25% astrocytes, and 26% oligodendrocytes [35]. Poor long-term cell survival on Matrigel suggests that the beneficial effect on cultured cells is predominantly because of nonquantified biological molecules present in Matrigel, and this is supported by the poor response when cultured on Collagen I. Although routinely used in neural cultures [36], there is a biological incompatibility between Collagen I and neural cells, as Collagen I is not found in brain tissue [35]. This highlights the insufficiencies of current 2D cell culture systems and presents bioactive SAPs as a potentially biocompatible, nonimmunogenic alternative that can provide a suitable environment for stem cell proliferation and differentiation in vitro.
Fluorenylmethyloxycarbonylchloride (Fmoc)–SAPs have also been investigated for biological applications [29]. These SAPs use a minimalist approach to forming complex structures through noncovalent interactions known as π−β self-assembly [37]. They are advantageous because of their ease of synthesis and ability to form a nanofibrous scaffold at physiological pH (Fig. 4) with the capacity to present bioactive epitopes at high density on the surface of the nanofiber [33]. Recently, the fibronectin cell adhesion sequence, arginine-glycine-aspartate (RGD), was incorporated into this SAP system by flanking the RGD sequence with structure-inducing aromatic residues to promote its self-assembly at physiological pH [38]. Human mammary fibroblasts were cultured on both an RGD Fmoc–SAP and a structurally and chemically analogous scrambled sequence, DGR, to assess the bioactivity of the sequence within the peptide sequence once assembled. On the DGR Fmoc–SAP, fibroblasts displayed reduced viability with apoptosis occurring until day 4 of the culture. In contrast, cell viability was maintained on the RGD Fmoc–SAP, suggesting that the cells recognized and interacted with the adhesion molecule [38]. In addition, morphological differences between cells cultured on the two different sequences were observed [38].
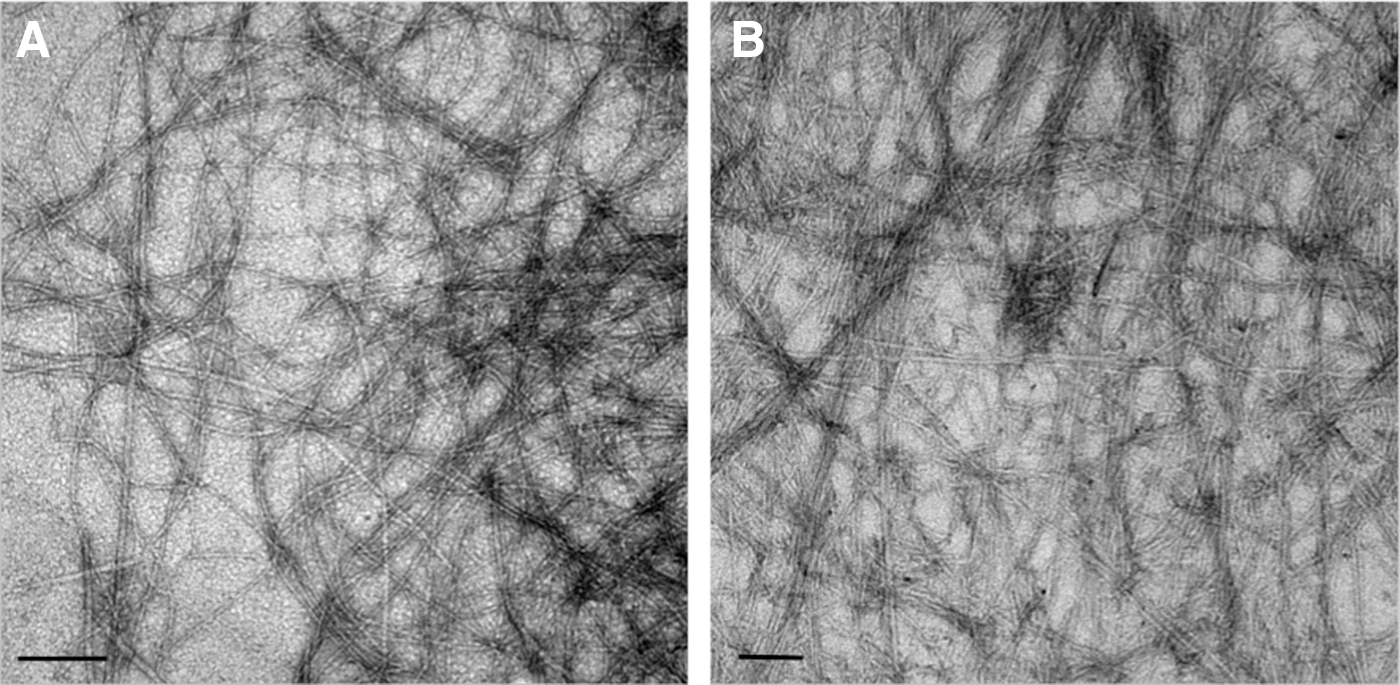
Transmission electron microscope images of nanoscale structures formed by Fmoc-based self-assembling peptides.
The mechanical and morphological properties of Fmoc–FRGDF can be tuned through the gelation mechanism, illustrating that these SAPs can be tailored for applications with particular mechanical and morphological requirements [39]. Gelation of Fmoc–FRGDF using glucono-δ-lactone resulted in the formation of nanofibrils with minimal entanglement, and hence, a weaker hydrogel, than those formed using a hydrochloric acid (HCl)-based pH-switch method. The HCl pH-switch method allowed the gel stiffness to be tuned, dependent on PBS concentration, without altering the self-assembly mechanism involving the formation of π−β sheets and antiparallel β sheet structures [39]. The influence of the gelation mechanism over the mechanical and morphological properties of SAPs diversifies the potential application of this sophisticated class of materials, holding great promise for tailoring SAP stiffness according to the desired differentiation pathway.
Therefore, through their nanofibrous structure, presentation of peptides that are similar to the ECM proteins, and tailored mechanical properties, SAPs can provide a cell culture system that effectively mimics various aspects of the in vivo environment, including relevant biochemical and topographical cues [29,35]. Combined with stem cells, this class of biomaterials could allow for the culture of cells in an in vitro environment morphologically representative of that found in vivo. In addition, the ability of SAPs to be tailored for tissue-specific applications through specific peptide sequences or stiffness could enable efficient differentiation of cells into a specific lineage, critical for the ultimate success of cell transplantation in neural regeneration, as well as necessary for accurate disease modeling and drug screening [40].
Biomaterials are ideal for developing new cell culture systems as their properties can be engineered to represent those of the native ECM. The provision of the ECM cues and a 3D environment essential to cell development can be exploited to create cell culture systems to produce high-quality cells for use in drug screening and enhance current disease models. In addition, they can facilitate directed cell differentiation, which is of use when designing disease-specific cell transplantation strategies that could also employ the use of biomaterials in vivo.
Applications of 3D biomaterial cell culture systems
Biomaterials have exhibited properties that are desirable for developing an alternative 3D cell culture system to produce large number of cells representative of those found in vivo, for a variety of applications. One shortcoming for the current in vitro cell culture and disease model systems is that cellular oxygenation, as well as nutrient and waste removal, occurs through diffusion, as the systems are not connected to the blood stream [41]. Integrating biomaterials and PSCs under dynamic cell culture conditions (such as those found in bioreactors) can provide critical mass transport to and from cells, circumventing potential mass diffusion limitations currently associated with 2D cell culture [11]. Using biomaterials in conjunction with bioreactors would improve the accuracy, and therefore efficacy, of the current in vitro disease models, as the in vivo environment would be better represented with the incorporation of physical and biochemical cues from the material and environmental control of the bioreactor.
Large scale cell generation and drug screening
2D cell culture systems have been identified as a limitation for the provision of sufficient cell numbers for applications such as cell replacement therapy (CRT) and drug screening because of their limited scalability and reproducibility [12]. Predictive toxicity assays during preclinical testing are essential for the development of new drugs [42]; however, the prediction of the pharmacokinetic bioavailability and indirect toxicity of such drugs are still limited in current culture systems [11]. As such, a large number of cells, which possess reliable disease pathology, as well as a superior culture environment, are required for improved drug screening technologies.
Biomaterials can be incorporated into culture systems to generate large cell numbers and provide a culture environment more representative of in vivo environment. Long-term culture of hESCs and hiPSCs in a thermoreversible hydrogel, poly(N-isopropylacrylamide)-co-poly(ethylene glycol) (PNIPAAm-PEG), resulted in a large cell yield, high-expansion rate, and high yield. It was also demonstrated that this 3D system supported the differentiation of hPSCs into the three germ layers as well as directed differentiation into NPCs and midbrain dopaminergic neurons. Combining such a biomaterial with a bioreactor to provide the optimal environment for cell proliferation and differentiation could yield more than 1011 cells from 5 L of hydrogel [12]. Such cell numbers can be used for applications including clinical studies employing cell transplantation, or high-throughput drug screening, potentially saving the time, money, and resources currently expended on unsuccessful clinical trials.
Disease modeling
The ability of stem cells, particularly iPSCs and ESCs, to recapitulate disease phenotype in vitro is promising for disease modeling. However, there are limitations associated with cells grown in 2D culture in accurately representing the behavior of those cells found in vivo. For example, gene expression patterns of ESCs possessing Mus musculus, Mus spretus, or Mus terricolor mitochondrial DNA haplotypes were different when cultured on electrospun 3D PCL scaffolds and 2D culture dishes. In addition, there was an 18–24-fold increase in gene expression in a 3D environment [43]. This indicates that there is a significant impact on cell behavior, dependent on the culture conditions. Therefore, it would be advantageous to implement biomaterials in future disease modeling experiments in an attempt to recapitulate the native cellular environment, and obtain meaningful results that can be used to understand disease pathology and the effect of potential therapeutic drugs.
In addition, encouraging iPSCs out of their embryonic state into populations of mature adult cells that behave reliably is a key challenge of their use in disease modeling [44]. This is of particular importance in modeling diseases such as ALS, Alzheimer's, and Parkinson's, where disease manifestation occurs in mature neurons, with synaptic properties typical of those in an adult brain. In conjunction with the development of appropriate differentiation protocols, it has been suggested that it may be necessary to develop culture conditions to allow for the full maturation of cells, as disease phenotypes may not be revealed in immature cells [44]. As such, 3D biomaterials can provide a platform on which to direct differentiation through the presentation of growth factors and cytokines and enable prolonged culturing systems not sustainable on 2D culture ware.
Biomaterials to Enhance Cell Transplantation Outcomes
Limitations of current in vivo treatment strategies
Injury to the CNS, such as trauma or stroke, can result in the formation of cystic cavities that impede cellular replacement and differentiation because of the absence of appropriate structural support [45]. In addition, current pharmacological treatment for neurodegenerative diseases does not slow disease progression, and in some cases can actually exacerbate symptoms or induce unwanted side effects [1]. Therefore, there is a need for the development of treatments to address disease progression and/or the injury response of the CNS. In addition to the limitations associated with the generation of suitable cells in 2D cell culture for use in CRT, there are also limitations associated with long-term graft–host integration. Populations of pure multipotent NSCs have previously shown poor survival in most CNS regions, as a result of the host failing to provide a sufficient environment to support the survival of NSCs [46]. Also, in the treatment of necrotic brain injuries, such as stroke, TBI, and cerebral palsy, often a void needs to be filled to facilitate regeneration. Biomaterials not only easily fill a void but also provide the physical and biochemical support required to recapitulate the stem cell niche when cells are transplanted.
Electrospun nanofibers provide physical and chemical cues in vivo
PCL nanofiber scaffolds have been shown to be biocompatible materials that can integrate with host brain circuitry, a requirement for the development of scaffolds for cell transplantation. Typically, the nanofiber scaffolds are rolled up on themselves into a “cylinder” of nanofibers, as shown in Fig. 5D, when implanted into the brain, as shown in Fig. 6. When randomly and partially aligned PCL nanofiber scaffolds were implanted into the caudate putamen of the adult rat brain, neurite infiltration was found to be dependent on fiber alignment [47]. Randomly aligned scaffolds were found to facilitate neurite infiltration, whereas neurites remained at the interface between partially aligned scaffolds and adjacent brain parenchyma (Fig. 5). This was attributed to the reduced interfiber distance of the partially aligned scaffolds inhibiting neurite infiltration. However, it was found that the partially aligned nanofibers provided a uniform perpendicular contact guidance to neurites, highlighting the potential for guided growth in vivo using partially aligned scaffolds for the development of disease-specific treatment strategies [47].

Neurite interaction with PCL electrospun scaffolds 60 days after implantation:
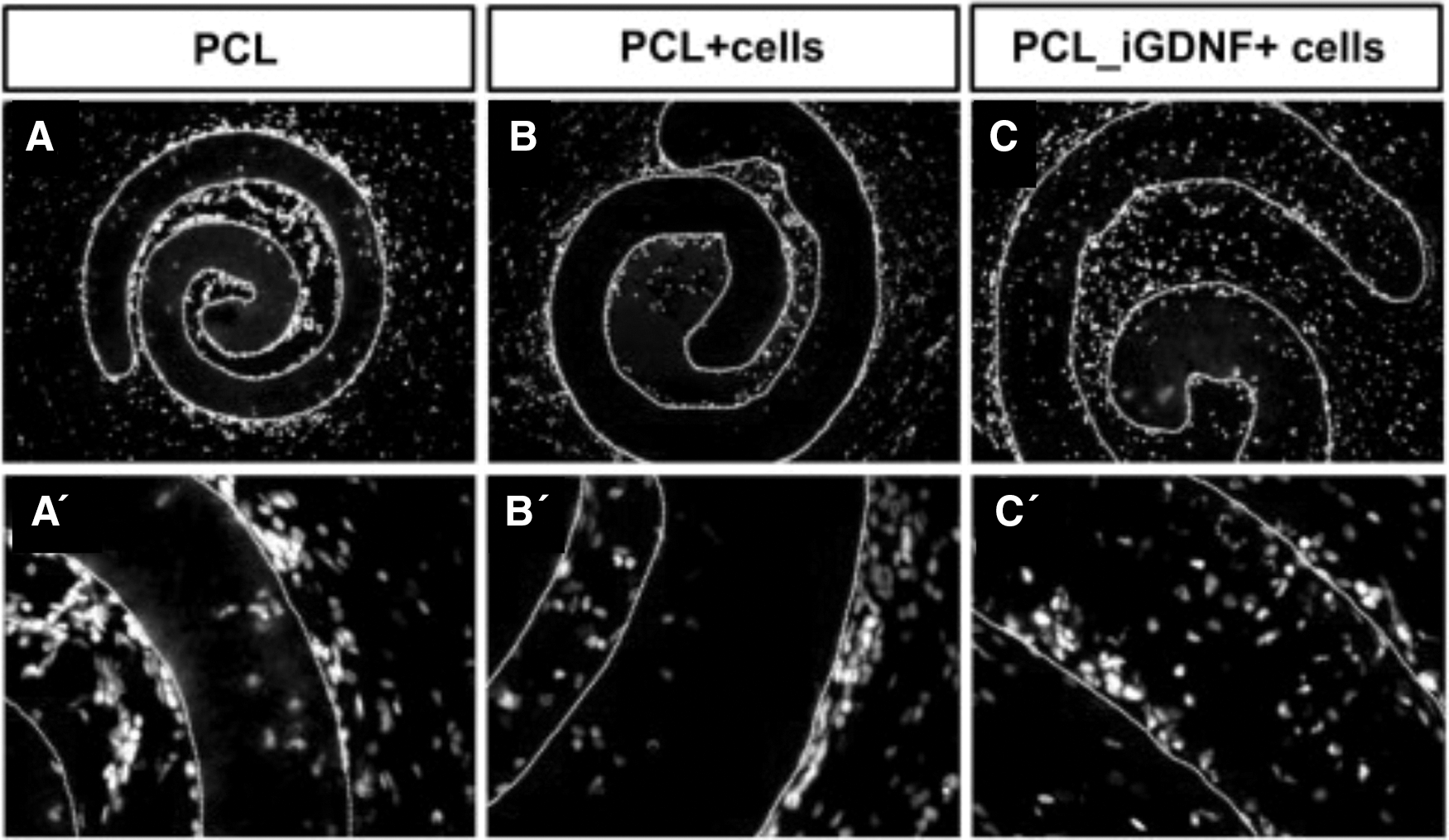
Micrographs showing the implanted nanofiber scaffold, rolled up on itself for implantation into the rat striatum, with migrated cells labeled for Hoechst.
Current methods of growth factor delivery such as direct injection are suboptimal, because despite achieving targeted delivery, a steep growth factor concentration gradient is created upon injection, which can alter or destroy tissue around the injection site [48]. In addition, as growth factors are readily degraded in their natural physiological environment, when they are delivered in vivo, degradation and diffusion to nontarget tissue limit their therapeutic value [7]. A more stable method to deliver growth factors is to use slow-releasing microspheres. Microspheres made of poly-lactic-co-glycolic acid have been found to release GDNF upon degradation at the site of implantation, which can last for several months [49]. However, this delivery method is still uncontrolled, with a nonconstant rate of growth factor release over the first few weeks. Therefore, it is desirable to control the behavior of growth factors by their stable presentation within multifunctional nanofibrous biomaterials. The influence of prolonged, stable protein presentation in vivo has been investigated by implanting NPCs together with functionalized (GDNF-tethered) PCL scaffolds into rat brains [8]. Cell viability significantly increased in the presence of immobilized GDNF compared to implantation of cells alone, demonstrating the ability of GDNF to maintain its trophic effects following tethering and implantation. Furthermore, there was a significant increase in immature neurons within the grafts exposed to immobilized GDNF as compared to the control cell graft and unfunctionalized PCL. This suggests that tethered GDNF is capable of continually supporting immature neurons within the graft, accounting for the increase in grafted NPCs [8].
The implantation of PCL with tethered GDNF was also found to improve stem cell graft outcomes in the intact and injured brain. Functionalized scaffolds enhanced the yield of neurons and oligodendrocytes within the graft, decreased reactive astrocytes, and increased the penetration of neuronal processes into the scaffold [8]. This demonstrates that the immobilization of GDNF on PCL scaffolds provides functional trophic cues capable of influencing cellular proliferation, differentiation, and neurite growth in vivo [8]. Therefore, neurotrophic factors are of great importance to improve transplanted cell integration, and the ability of electrospun nanofibrous scaffolds to be readily functionalized to control cell behavior is also highlighted.
The modification of the conventional electrospinning setup can also allow the fabrication of cylindrical nanofiber scaffolds that replicate the spinal cord. The implantation of PLGA nanofiber cylindrical grafts, fabricated using water vortex electrospinning, into a transected spinal cord rat model resulted in better locomotive and sensory scores than the control group that did not receive scaffold implantation [50]. This can be attributed to the physical support provided by the nanofiber graft for axonal growth at the interface of the spinal cord and scaffold, as well along the scaffold nanofibers [50]. This scaffold could be enhanced by further chemical functionalization, as previously discussed, to potentially improve the functional recovery of injured spinal cords.
Electrospun nanofibrous scaffolds have the ability to provide physical and chemical cues in vivo to promote tissue repair, and cylindrical nanofiber grafts have great potential for the treatment of SCIs with further development of material choice and scaffold functionalization. However, geometric limitations imposed by the fabrication mechanism make them unable to fill a void effectively, an important requirement of treatment strategies in certain acquired brain injuries. In this regard, treatment of neural injuries, neurodegenerative or traumatic, should be targeting the use of void filling nanofibrous hydrogels.
Hydrogels to support transplanted cell growth in vivo
Hydrogels are a promising biomaterial for implementation in neural regeneration strategies as they have clearly demonstrated their suitability to provide physical support to transplanted cells and the ability to be functionalized with multiple growth factors. Incorporating stem cells with functionalized hydrogels provides the potential for preferential differentiation in vivo and the development of application-specific cell/biomaterial systems.
The physical and chemical diversities that are possible with hydrogels make them desirable candidates for cell transplantation applications. Implantation of xyloglucan, a thermally gelling hydrogel, functionalized with poly-
In addition, when seeded with mescenchymal stem cells and implanted into hemisected rat spinal cords, hydroxypropylmethacrylate hydrogels, either polymerized in the presence of a solid porogen (HPMA-SP), with the fibronectin sequence RGD attached (HPMA-SP-RGD), or prepared by heterophase separation (HPMA-HS-RGD), as well as the HEMA-MOETACl (hydroxy ethyl methacrylate [2-(methacryloyloxy)ethyl] trimethylammonium chloride) hydrogel, all promoted blood vessel and axon growth inside the scaffold, dependent on their physical and chemical properties [53]. It was found that blood vessel growth was promoted when RGD was presented in the hydrogel, whereas axon growth inside the scaffold was encouraged when it possessed the web-like architecture of HPMA-SP(-RGD) [53]. These systems have illustrated that they are biocompatible in vivo, and demonstrate that altering the surface chemistry and physical architecture of hydrogels can influence cell infiltration and growth.
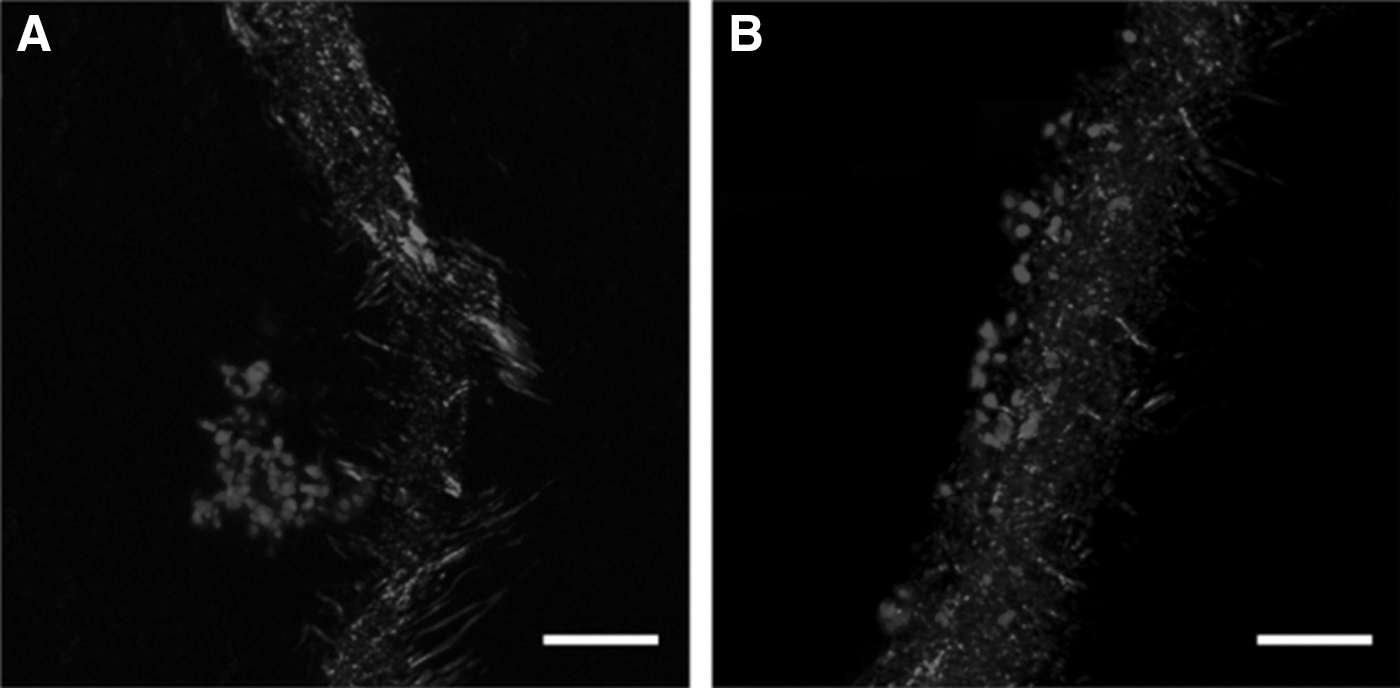
Hyaluronan-methyl cellulose (HAMC) is of relevance for neural regeneration applications as it gels at physiological temperatures and has been used as a drug delivery vehicle in the CNS. Encapsulating HAMC with epidermal growth factor (EGF) or poly(ethylene glycol)-modified EGF (PEG-EGF) was found to enhance the proliferation of NSPCs in the subventricular zone, in both uninjured and stroke injured brains [54]. In addition, when rat NSPCs mixed in platelet-derived growth factor-A immobilized to HAMC (HAMC-PDGF-A) were transplanted into a rat SCI model, the differentiation of NSPCs to oligodendrocytes was enhanced as compared to NSPCs transplanted in culture media [6]. Greater cell survival was observed in rats receiving NSPC/HAMC-PDGF-A, which was attributed to the presence of HA, as it is a key component of the ECM, and its interaction with cells through CD44 and RHAMM receptors influences cell survival and migration, which could account for the observed in vivo NSPC survival enhancement [6]. In addition, HA and MC have antioxidant properties that can reduce the flux of free radicals at the cell surface, reducing cell damage and enhancing NSPC survival [6]. Rats injected with NSPC/HAMC-PDGF-A exhibited more neurons alongside the lesion site than those that received NSPC/media; with 33% more spared neurons in the NSPC/HAMC-PDGF-A transplants (Fig. 7).
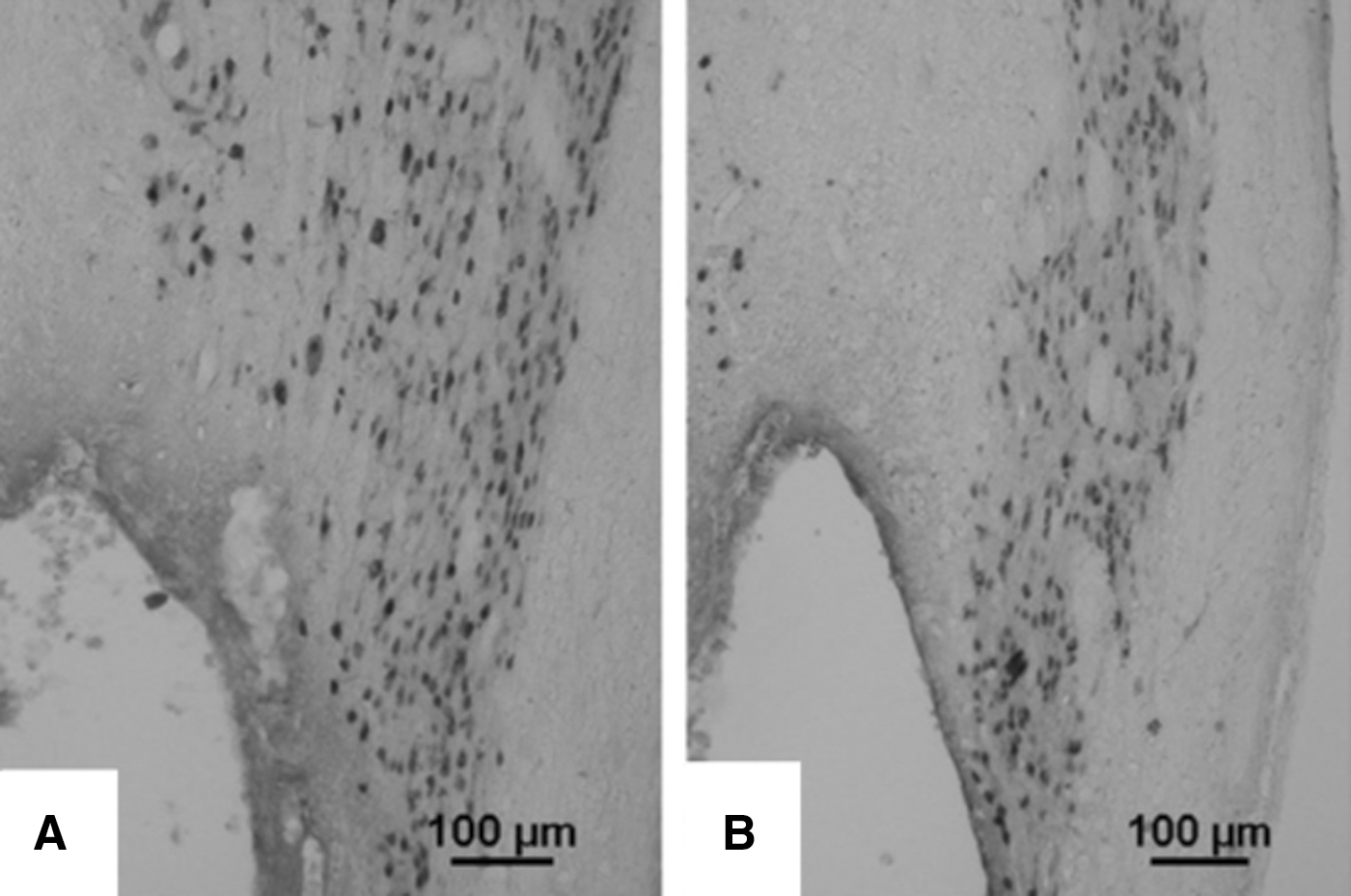
Photomicrographs illustrating NeuN staining of transplanted neural stem/progenitor cell in the presence of
NSPC/HAMC-PDGF-A transplants also resulted in an increase of 23% more host oligodendrocytes, demonstrating the benefit to host tissue achieved using NSPC/HAMC-PDGF-A transplants. This was attributed to the sustained presence of PDGF-A, as well as the presence of the HA component that has been shown to block NMDA-induced neuronal cell death. These studies demonstrate that HAMC is a promising hydrogel for both growth factor delivery and cell transplantation in the CNS. Further investigation of this material and its regenerative capacity when delivering various growth factors in combination with PSCs to support and direct differentiation, as well as to encourage regeneration, is an exciting prospect.
Fibrin-based hydrogels have also been utilized to deliver stem cells and growth factors in a dorsal hemisection subacute SCI in rats [55]. Embryonic neural stem/progenitor cells (ENSPCs) were encapsulated in fibrin hydrogels that were either unmodified, mixed with neurotrophic factor-3 (NT-3) and PDGF, or covalently attached to a heparin binding delivery system (HBDS), which then bound to the heparin binding growth factors NT-3 and PDGF. Poor cell survival was reported for ENSPCs transplanted without a fibrin scaffold, emphasizing the importance of physical support for transplanted cells. Cell numbers showed a 10-fold increase at 2 weeks when ENPSCs were transplanted in the fibrin hydrogel with the growth factors (with or without HBDS), whereas only a 2-fold increase was observed for the cells encapsulated in unmodified fibrin or transplanted without fibrin. In addition, there was an increased number of ENSPC-derived NeuN+ mature neurons when the ENSPCs were encapsulated in the fibrin/HBDS/NT-3/PDGF scaffold. This could be attributed to the HBDS limiting diffusion of the growth factors, allowing NT-3 and PDGF to influence cell behavior for a longer period of time.
Self-assembling peptides
SAPs hold great promise for in vivo applications, since their assembly is triggered by physiological changes in pH and temperature, allowing them to be injected directly into an injury site in a minimally invasive manner [56]. Importantly, the structure of SAPs can influence cell behavior, as was demonstrated when the IKVAV peptide and the IKVAV PA (self-assembles in vivo, Fig. 8) were injected into a clip compression model of SCI in mice. Functional recovery was not promoted with the injection of the IKVAV peptide alone, demonstrating that behavioral improvements are attributable to not only the IKVAV sequence but also the nanostructure of the self-assembled peptide [57].

At 9 weeks, the IKVAV PA-injected group had significant functional recovery compared to the control groups, indicating its ability to facilitate long-term functional recovery. At 5 and 11 weeks after SCI, the progression of astrogliosis at the lesion site in the IKVAV PA-injected group was suppressed, whereas process extension in astrocytes or the initial hypertrophy that may be essential for repairing the blood–brain barrier and restoring homeostasis was not altered [57]. It was also observed that IKVAV PA injection increased the oligodendroglial cell numbers while concurrently reducing apoptotic cell death and reduction of glial scar formation. This is significant as the IKVAV PA allows the initial functions of astrogliosis that are beneficial to CNS regeneration, while limiting the progression of gliosis that restricts axon outgrowth [57]. In addition, IKVAV PA was found to have degraded after 4 weeks, so the degradation time of IKVAV PA and other SAPs could be advantageous to provide temporal chemical cues to cells in vivo.
The physical support provided by SAPs is also an important feature to consider when developing scaffolds for CRT. RADA16 or RADA16–IKVAV with encapsulated NSCs (RADA16/NSCs and RADA16–IKVAV/NSCs, respectively) self-assembled into a 3D nanofibrous hydrogel immediately after in situ injection into a cavity in a rat brain [30]. As compared to NSCs suspended in the cavity without the peptide hydrogel, cell survival was enhanced when NSCs were transplanted within the peptide hydrogel. NPCs and neurons were present 3 weeks after the transplantation of RADA16–IKVAV/NSCs, whereas few NPCs and neurons were present when RADA16–IKVAV was transplanted without NSCs. This suggests that NSCs are integral to the repair and regeneration of tissue. In addition, reactive astrocytes were significantly reduced around the boundary of the implanted RADA16–IKVAV/NSCs, as compared to the RADA16/NSCs, demonstrating that the functionalization of RADA16–IKVAV could reduce glial scarring through the suppression of astrocytic differentiation [45].
The peptide K2(QL)6K2 (QL6) is another SAP that has shown potential for the functional recovery of rat SCI. Injection of the QL6 SAP 24 h after clip compression resulted in a reduction of astrogliosis and apoptosis, and improved Basso, Beattie, and Bresnahan (BBB) locomotor scores as compared to the control group that did not receive any SAP. The biocompatible QL6 provided the physical and chemical support necessary for the suppression of an inflammatory response, as well as axon survival and sprouting. However, it was noted that there was a lack of neurons in the grafted SAP [58], so using SAPs to support cell grafts would be promising for these materials. NPCs delivered using Fmoc–SAPs such as Fmoc–FRGDF, Fmoc–DYISGRF, and Fmoc–DIKVAV resulted in a limited foreign body response and innervated the host brain tissue after 28 days. This study also highlights the potential for tissue-specific SAP development, as YIGSR and IKVAV are laminin epitopes, whereas RGD is derived from fibronectin. Therefore, these SAP sequences can be tailored for the desired tissue application, which could potentially influence the differentiation of stem cells in vivo. A possible extension of this treatment strategy would be to combine it with PSCs such as ESCs or iPSCs to enhance the cell growth in the scaffold, and hence, across the lesion site. However, although SAPs have been thoroughly characterized for their material properties, the biological characterization of SAPs is limited and warrants further investigation.
Future Perspectives
Hydrogels, SAPs, and electrospun nanofibrous scaffolds hold great promise for the development of 3D culture systems and in vivo applications in the quest to improve the development of new therapies in the treatment of brain injuries. Here we have described their benefits of improved in vitro culturing for the purpose of disease modeling, drug development, and cell transplantation. These systems illustrate that the provision of a microenvironment with structural composition and elasticity of the native tissue, as well as high water content, improves implanted cell survival in vivo, and thus, are promising scaffolds. The ability of bioengineered scaffolds to provide an environment representative of the ECM, allow for cellular infiltration, and control of cellular behavior through functionalization suggests that they are ideal biomaterials to combine with cells to further the field of neural regeneration, both in vitro and in vivo. In addition to NSCs and NSPCs, PSCs such as ESCs and iPSCs are promising cell sources because of their unrestricted capacity for differentiation and proliferation. However, minimal research has been conducted to investigate the performance of these biomaterials as scaffolds to facilitate ESC or iPSC differentiation and proliferation in vitro, or neural regeneration using ESC or iPSC grafts in vivo. Therefore, to fully understand and exploit the behavior of stem cells for neural regeneration, it is suggested that biomaterials including hydrogels, SAPs, and electrospun nanofibers should be combined with such cell sources to significantly enhance current cell culture systems and in vivo neural regeneration strategies.
Footnotes
Acknowledgments
F.L.M. and A.L.R. were supported by an Australian Postgraduate Award; C.L.P. was supported by an NHMRC Career Development Award, and subsequently Senior Medical Research Fellowship provided by the Viertel Charitable Foundation, Australia; R.J.W. was funded by an Alfred Deakin Research Fellowship; and D.R.N. was supported by an NHMRC Career Development Fellowship (APP1050684). Access to the facilities of the Centre for Advanced Microscopy (CAM) with funding through the Australian Microscopy and Microanalysis Research Facility (AMMRF) is gratefully acknowledged.
Author Disclosure Statement
No competing financial interests exist.