
Abstract
The evolutionarily conserved Notch and Wnt signaling pathways have demonstrated roles in normal mammary gland development and in breast carcinogenesis. We previously reported that in human mammary gland, signaling through NOTCH3 alone regulates the commitment of the undifferentiated bipotential progenitors to the luminal cell fate, indicating that NOTCH3 may regulate the expression of unique genes apart from the other Notch receptors. In this study, we used gain of function and loss of function experiments and found that a Wnt signaling receptor, Frizzled7 (FZD7), is a unique and nonredundant target of NOTCH3 in human breast epithelial cells. Interestingly, neither the constitutively active forms of NOTCH1-2, 4 nor loss of expression of these receptors were able to alter expression of FZD7 in human breast epithelial cells. We further show that FZD7-expressing cells are found more frequently in the luminal progenitor-enriched subpopulation of cells obtained from breast reduction samples compared with the undifferentiated bipotent progenitors. Also, we show that NOTCH3-induced expression of FZD7 occurs in the absence of CSL (CBF1-Suppressor of Hairless-Lag-1). Our data suggest that noncanonical Notch signaling through NOTCH3 could modulate Wnt signaling via FZD7 and in this way, might be involved in luminal cell differentiation.
Introduction
T
In addition to the canonical signaling, the CSL-independent (noncanonical) signaling by Notch receptors has also been reported [7]. While the mammalian Notch receptors display both overlapping and distinct tissue expression patterns during development and in adult tissues [8], the current model of canonical Notch signaling suggests that these receptors play redundant functions in terms of their transactivation functions and their biological roles [6].
Notch signaling plays important roles in luminal cell fate determination in the mammary gland [9,10]. Luminal cells in the mouse and human mammary glands are continuously produced due to the proliferation and differentiation of luminal progenitors that are themselves generated from uncommitted bipotential progenitors [11 –13]. The bipotent progenitors are ultimately produced from a self-renewing population of breast stem cells [12,14]. We previously reported that NOTCH3 transcripts were highly expressed in the human luminal progenitors compared to the bipotent progenitors and the opposite expression pattern was observed for the NOTCH4 transcript levels [10].
However, NOTCH1 and NOTCH2 receptors were ubiquitously expressed in both progenitor subtypes and in their differentiated progeny. Furthermore, we showed that loss signaling through NOTCH3 alone led to decreased production of luminal cells from bipotent progenitors [10]. These findings suggest that NOTCH3 could exhibit nonredundant functions during the process of bipotent cell commitment to the luminal differentiation pathway and that NOTCH3 may regulate expression of specific genes, apart from the other Notch receptors.
In this study, we provide evidence that the Notch-induced expression of a Wnt signaling receptor, frizzled7 (FZD7), is exclusively regulated by noncanonical signaling through NOTCH3 receptor in the human mammary epithelial cells, suggesting that the Notch-mediated modulation of the Wnt signaling pathway could be regulated through the NOTCH3-FZD7 signaling network.
Materials and Methods
Nonmalignant human breast epithelial cell lines
The human mammary epithelial cell strain (HMEC) was obtained from Lonza (
Isolation of primary mammary epithelial cell subsets
The breast reduction samples were obtained through informed written consent (University of Manitoba, Research Ethics Board #H2010:292) and were dissociated into organoid-enriched fractions and subsequently turned into single-cell suspension as described [16]. Single cells were stained with antibodies raised against EpCAM (1:100 dilution StemCell Technologies, Inc.) and α6 integrin (CD49f), conjugated to Alexafluor 647 (1:100 dilution, StemCell Technologies, Inc.). A goat anti-rat Fluorescein isothiocyanate was used to detect the EpCAM protein and propidium iodide exclusion was used to identify live cells (for staining procedure see reference 9). Lineage-positive cells (CD31+ and CD45+) were removed by using biotinylated specific antibodies and streptavidin-PE Tex-Red secondary antibody. The luminal progenitors (EpCAMbrightCD49flow) or the bipotent progenitors (EpCAMlowCD49fbright) were isolated via Fluorescent-Activated Cell Sorting (FACS) as described [10].
Protein expression analysis
Flow cytometry was used to examine FZD7 protein expression and the different Notch receptors. For this purpose, specific antibodies raised against FZD7 (1:100 dilution; R&D Systems clone# 151143), NOTCH1 (Cat No. A301-894A; Bethyl Laboratories), NOTCH2 (Cat No. A301-083A; Bethyl Laboratories), and NOTCH3 (Cat No. 2889; Cell Signaling) were utilized. For flow cytometry, single-cell suspensions were prepared from the breast reduction samples or 184-hTert cells or the HMECs essentially as described [16].
Protein expression was detected using a goat anti mouse antibody conjugated to PE (Jackson ImmunoResearch Laboratories, Inc.) and quantified using the FlowJo Software (TreeStar, Inc.). Expression of NOTCH4 and the CSL protein levels were examined using a polyclonal anti NOTCH4 antibody (Cat No. ARP32726; Aviva Systems biology) and polyclonal anti CSL antibody (Cat No. sc-28713; Santa Cruz Biotechnology) respectively, using conventional western blot assay and normalized to the β-actin protein expression.
Lentiviral transduction
HMECs or the 184-hTERT cells were transduced with lentivirus expressing a short hairpin RNA (shRNA) to knockdown expression of each Notch receptor separately. The lenti-shRNA constructs (pool of three separate lenti-shRNA for each Notch receptor) and a lenti construct carrying a scrambled shRNA sequence were obtained from OpenBio Systems (Thermo Scientific). The lenti-shRNA-transduced cells were selected using puromycin (2 μg/mL; Sigma Aldrich). The forced expression of human Notch receptors was achieved using lentivirus expressing the constitutively active (intracellular domain [ICD]) form of each Notch receptor.
Plasmid constructs carrying human NOTCH-ICD cDNA were obtained as follows: the pcDNA3-Notch1-ICD vector was a kind gift from Dr. Andrew Weng (Terry Fox Laboratory, Vancouver, B.C. Canada), NOTCH2-ICD and NOTCH3-ICD cDNA fragments were polymerase chain reaction (PCR) cloned (Supplementary Table 1; Supplementary Data are available online at
Empty vector control and NOTCH3-expressing 184-hTERT cells were reinfected with three separate lentivirus expressing three different short hairpin RNA (shRNA) to knockdown CSL expression or lentivirus construct carrying a scrambled shRNA sequence. The scrambled shRNA and lenti-shCSL-transduced cells were selected using puromycin (2 μg/mL; Sigma Aldrich) for 48 h.
Breast reduction samples where dissociated into single cells and cultured overnight as described and subsequently infected either with lentivirus expressing empty vector (EV) or NOTCH3-ICD expressing lentivirus for 4 h. After 72 h, cells expressing the GFP+ cells (ie, NOTCH3 overexpressing cells) were examined for their expression of FZD7 via flow cytometry.
Transcript expression analysis
RNA was isolated from the lenti-shNOTCH or lenti-NOTCHICD or lenti-shScrambled or lenti-empty vector control-transduced cells (three separate infections using pool of three different lenti-shRNA virus) using the TRIzol reagent (Invitrogen) according to the manufacturer's protocol and treated with RNase-free DNase (Promega). The differential transcript expression of genes in these RNA samples was investigated using quantitative real-time PCR (qPCR). The qPCR data were analyzed using the comparative threshold cycle method (ΔCT), and normalized against the β-actin and GAPDH transcript expression.
Chromatin immunoprecipitation
184-hTERT cells were infected with constructs expressing a FLAG-tagged version of NOTCH1 or NOTCH3 (Addgene) or a construct expressing FLAG-GFP alone. The FLAG alone construct used is a nonlenti viral vector (pTAG-1) that contained a mammalian promoter to express FLAG in the transfected cells. The transduced cells we isolated via FACS using GFP expression as the section marker. Sorted cells were dual cross-linked as described previously [18]. The dual cross-linked chromatin was sonicated and chromatin immunoprecipitation (ChIP) was performed using anti-FLAG antibody (Cat. No. F1804; Sigma Aldrich). The ChIP DNA was used as a template for qPCR. Upstream promoter sequence of FZD7 was retrieved from the Eukaryotic Promoter Database (
Results
PCR array identifies FZD7 as a potential NOTCH3-specific target gene
To identify the unique gene targets of NOTCH3 signaling, the lentviral transduction was employed to knock down NOTCH3 and NOTCH4 expression or to express a short hairpin scrambled control sequence in HMEC cells and the differential expression of 84 Notch signaling targets and associated genes were examined using the SABiosciences, Notch signaling qPCR array. The qPCR array analysis revealed that a Wnt signaling receptor, FZD7, low-density lipoprotein receptor-related protein 5 (LRP5), and delta-like 3 (DLL3) transcript levels were decreased in lenti-shNOTCH3-transduced cells and not in the lenti-shNOTCH4-transduced cells (Supplementary Fig. S1). The preliminary data obtained through the qPCR array were validated using independent primer sets and HMECs transduced with lenti-shNOTCH1-4.
The knockdown level of each Notch receptor in the transduced cells was determined through quantitative qPCR and intracellular FACS (Supplementary Fig. S2A, B). LRP5 transcript levels remained unchanged in the transduced cells, whereas the DLL3 transcript levels were decreased in lenti-shNOTCH3 and lenti-shNOTCH4 HMECs, which is contrary to the qPCR array data. This discrepancy could be due to defective primers used in the qPCR array plates to detect DLL3 and LRP5 transcripts. FZD7 transcripts, however, were decreased only in the lenti-shNOTCH3 HMEC cells as compared with scrambled control transduced HMECs (Fig. 1A).
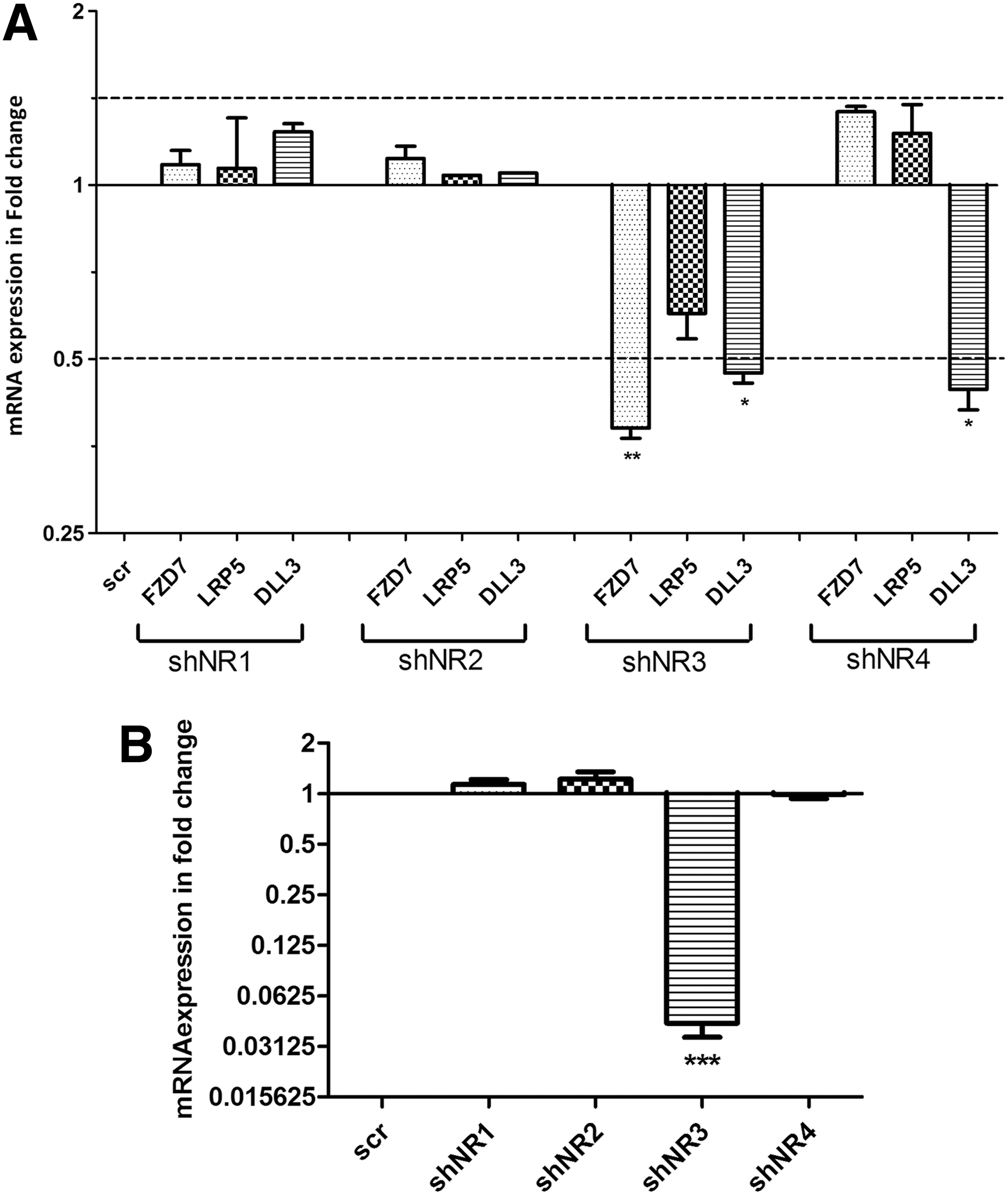
NOTCH3 regulates expression of Frizzled7 (FZD7)
To further validate this observation, another source of nonmalignant human breast epithelial cells (184-hTert cell line), were infected with different lenti-shNOTCH virus (Supplementary Fig. S3A, B) and FZD7 transcript expression was examined using qPCR. Similar to the HMEC cells, we found that FZD7 transcript levels were only decreased in the lenti-shNOTCH3-infected 184-hTert cells (Fig. 1B). Therefore this preliminary qPCR array analysis suggests that FZD7 may be a unique target of NOTCH3 signaling.
Notch-induced FZD7 expression is regulated by NOTCH3 only
Essential to the initiation of Notch signaling is the release of the active NICD, which upon nucleation governs transcriptional regulation of Notch-target genes. Therefore, we forced the expression of a constitutively activated form of each Notch receptor (NOTCH1-4) in the 184-hTERT cells (Supplementary Fig. S4A, B) and examined changes in the FZD7 transcript. Interestingly, only increased signaling through NOTCH3 significantly increased the expression of FZD7 transcripts (2.98 ± 0.198-folds) (Fig. 2A). Using flow cytometry we observed that FZD7 protein expression level also changes only when signaling through NOTCH3 is activated (Fig. 2B,D).

FZD7 is a unique target of NOTCH3 184-hTert cells and was infected with lentivirus expressing the constitutively active form of each Notch receptor (NR1-4) or empty vector (EV) control and the transcript expression of FZD7 was examined via qPCR
Previous studies have shown that NOTCH1 could effect the expression of NOTCH3 and NOTCH4 [19,20]. Therefore, it is likewise possible that changes in NOTCH3 expression could alter the expression of other Notch receptors. Using qPCR, we found that loss of NOTCH3 in 184-hTert cells led to small (<2-fold) decrease in the transcript expression of NOTCH1 and NOTCH2 while the transcript expression of NOTCH4 remained unperturbed (Supplementary Fig. S5B). Also, overexpression of active form of NOTCH3 led to a 2-fold increase in NOTCH1 and NOTCH2 expression without altering the expression of NOTCH4 (Supplementary Fig. S5C). However, we found that activation of signaling though NOTCH1, 2, or 4 has no effect on FZD7 expression (Fig. 2A–D). Since only activated NOTCH3 signaling enhances FZD7 expression suggests that preferential activation of NOTCH3 may not be required for specific transcriptional activation of FZD7.
Luminal progenitors exhibit high expression of NOTCH3 and FZD7
FZD7 is a Wnt signaling receptor that is involved in the proliferation and differentiation of progenitors and stem cells [21 –24]. We therefore examined whether FZD7 is differentially expressed among the undifferentiated human breast epithelial cells (ie, stem and progenitor cells). Previously, we showed that stem cells/bipotential progenitors and luminal progenitors could be obtained from the breast reduction samples based on their expression of EpCAM and CD49f [25]. We observed that overall, only 1%–1.5% of the uncultured normal human breast cells express FZD7 (data not shown) and that bipotent and the luminal progenitors express the FZD7 protein. Interestingly, however, luminal progenitors contained more FZD7 + cells and expressed FZD7 transcripts and protein at a higher level compared with bipotent progenitors. (Fig. 3A–D).
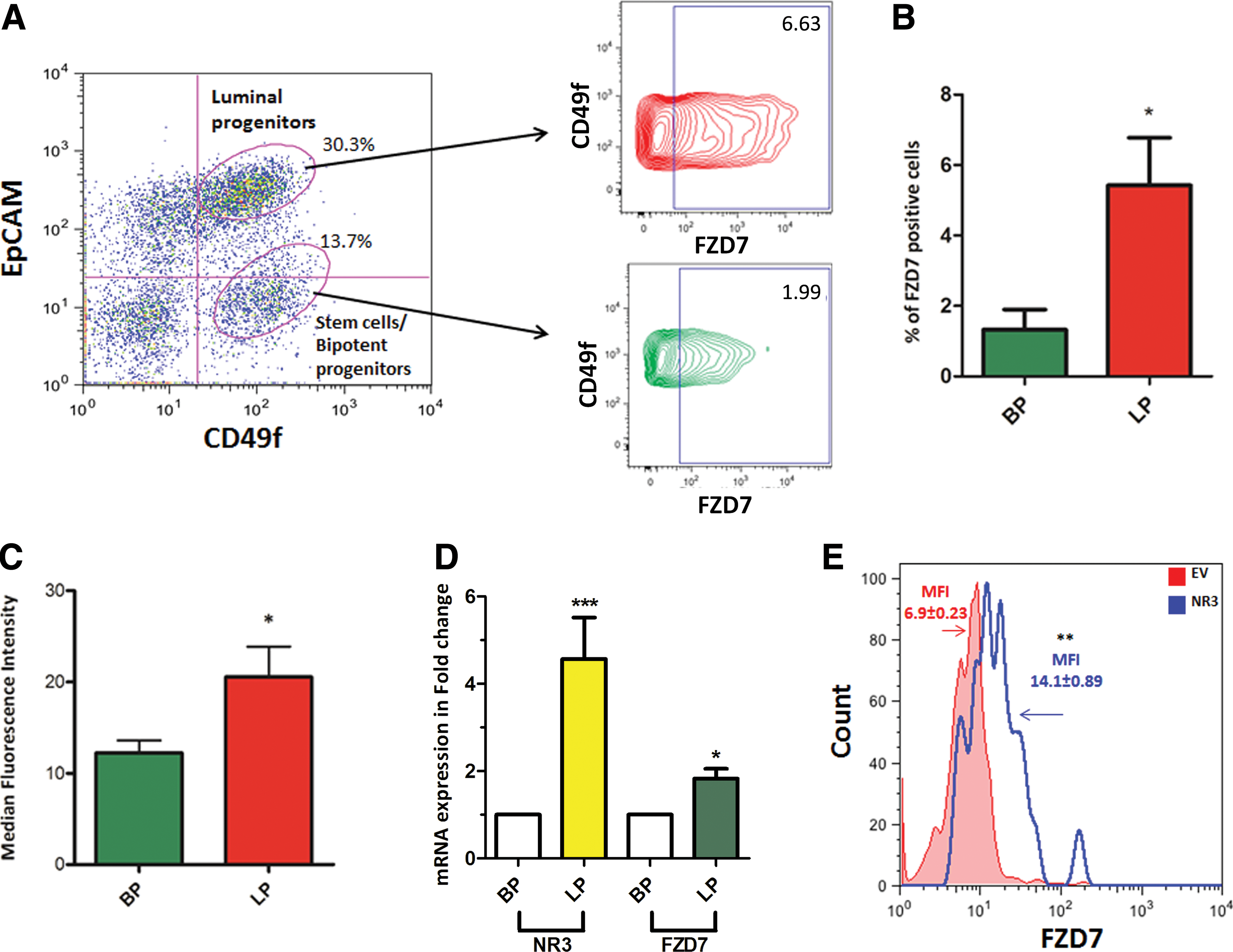
Luminal progenitors show high expression of FZD7 and NOTCH3
In addition, increased signaling through NOTCH3 significantly increased FZD7 expression in the primary human breast epithelia cells obtained from breast reduction samples. These observations are significant because we had previously shown that the luminal progenitors express NOTCH3 at a much higher level compared with the stem/bipotent progenitors (Fig. 3D and [10]). Moreover, it was recently shown that Wnt signaling could restrict luminal/alveolar differentiation and decrease NOTCH3 expression by modulating the Pygo2 expression [26], suggesting that a strong crosstalk between NOTCH3-Wnt exists. In addition, FZD7 has also been reported to regulate progenitor cell functions in endoderm cells [24]. Therefore, it is inviting to hypothesize that NOTCH3 could regulate luminal cell differentiation through NOTCH3-FZD7 signaling axis.
Notch3-regulated expression of FZD7 is CLS-independent
To study whether NOTCH3 regulates FZD7 expression through canonical signaling we used ChIP to quantify NOTCH3 enrichment on the FZD7 promoter. 184-hTERT cells were transduced with FLAG, NOTCH1-FLAG, and NOTCH3-FLAG expressing constructs and ChIP was performed using anti-FLAG antibody. Compared to IgG isotype control, no significant enrichment for NOTCH1 or NOTCH3 could be observed on the two putative CSL binding sites in the FZD7 enhancer/promoter region. HES1 promoter, however, showed significant enrichment for both NOTCH1 and NOTCH3 (Fig. 4A and Supplementary Fig. S6A, B).
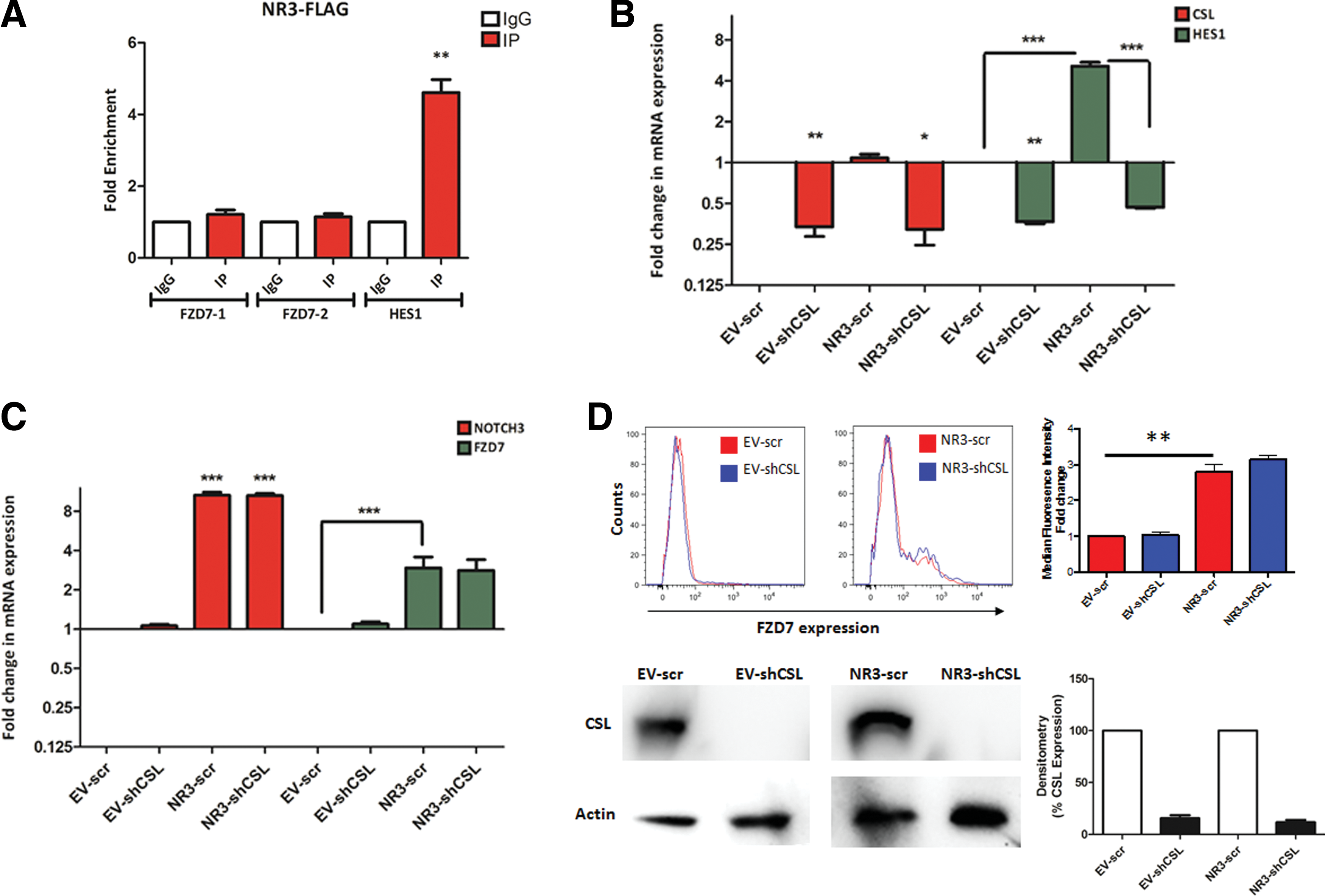
NOTCH3 regulates FZD7 expression in a CBF1-Suppressor of Hairless-Lag-1-independent manner
These observations suggest NOTCH3 could regulate FZD7 expression in a noncanonical manner. To investigate this possibility, NOTCH3-expressing 184-hTERT cells were transduced with lenti-scrambled or lenti-shCSL and expression of FZD7 was determined via qPCR and FACS (Fig. 4C, D). Interestingly, in the absence of CSL, FZD7 expression showed significant increase in NOTCH3-expressing cells, suggesting that the NOTCH3-induced FZD7 expression does not require CSL (Fig. 4C). As expected, NOTCH3 regulation of HES1 transcript expression, however, required CSL expression (Fig. 4B).
Discussion
Previously, role of Notch signaling and, in particular, specific requirement of NOTCH3 in regulating luminal cell differentiation had been described. These studies suggested that NOTCH3 regulates expression of a unique set of target genes apart from other Notch receptors. In this study, we utilized gain of function and loss of function studies and found that Notch-induced FZD7 expression is only regulated by NOTCH3 in a CSL-independent manner in human breast epithelial cells. Furthermore, we observed that the luminal progenitor subpopulation of human breast epithelial cells exhibits a stronger expression of FZD7 along with NOTCH3 compared with the stem/bipotent progenitor subpopulation.
The role of FZD7 in regulating mouse mammary gland development was recently described by Chakrabarti et al. [27] where the authors showed that the cooperation between ΔNp63 and fzd7 could regulate mouse mammary stem cell functions. Similarly, we observed FZD7 expression in the stem/bipotential progenitor subpopulation of human breast epithelial cells suggesting that FZD7 may also play a role in regulating the human stem/bipotent progenitor cell functions. The link between the Notch-Wnt signaling crosstalk and luminal/alveolar differentiation was recently demonstrated by Gu et al. [26]. Using the Pygopus 2 (Pygo2)-null mouse breast cells, the authors demonstrated that the canonical Wnt signaling antagonizes Notch3 expression in a Pygo2-dependent manner and diminishes luminal/alveolar differentiation potential of breast cells.
In this study, we found that FZD7 expression was higher in the committed luminal progenitor that also express high levels of NOTCH3 compared to the uncommitted bipotent progenitors that express very little NOTCH3. We previously showed that that in human mammary gland NOTCH3 signaling is essential the commitment of undifferentiated bipotent progenitors to the luminal cell fate [10]. The potential crosstalk between the Notch1 and Fzd6 & 10 but not Fzd7 were observed during dendritic cell differentiation in mice, indicating that the Notch-Wnt crosstalk could regulate important cellular processes [28]. The data presented in the report also suggest that the Notch-dependent expression of FZD7 gene is regulated by NOTCH3 in a noncanonical manner.
The noncanonical regulation of Notch target genes has been previously demonstrated before where it was shown that NOTCH1 forms an activating complex with the YY1 transcription factor and enhances c-Myc expression in a CSL-independent manner [29]. It is therefore possible that through this or similar mechanisms, NOTCH3 specifically regulates expression of its unique target genes such as FZD7.
The Notch and the Wnt signaling pathways represent two of the highly conserved signaling pathways that regulate many aspects of embryonic development and cell turnover and tissue regeneration in adult tissues. Therefore, understanding how these two signaling pathways interact would expand our understanding of their contributions to tissue development but also how alterations to nature of crosstalk between these signaling pathways may contribute to disease processes.
The potential role of FZD7 in breast cancer development, particularly in the context of triple-negative breast tumors, has been described previously [30 –32]. Lastly, while in this report we showed NOTCH3 uniquely regulates FZD7 expression, it is conceivable that other Notch receptors, likewise, activate expression of unique set of genes. Therefore, application of unbiased transcriptome profiling approaches to identify unique targets of each Notch receptor and their independent contribution to mammary gland development and maturation would be highly desirable.
Footnotes
Acknowledgments
The authors thank the plastic surgery team at the University of Manitoba and the Manitoba breast tumor bank for their collaboration in obtaining breast reduction samples. This work was supported through operating grants from the Canadian Health Research Institute and the Regional Partnership Program (RFN109757), start up funds from the University of Manitoba and the CancerCare Foundation to A.R. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of this article.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.