
Abstract
The reprogramming of somatic cells to induced pluripotent stem cells (iPSCs) is a complex process that involves significant epigenetic alterations in the reprogrammed cells. Epigenetic modifiers such as histone deacetylase (HDAC) inhibitors have been shown to increase the efficiency of derivation of iPSCs in humans and mice. In this study, we used three HDAC inhibitors, valproic acid, sodium butyrate, and suberoylanilide hydroxamic acid, together with ascorbic acid, for derivation and long-term feeder-free culture of porcine iPS-like cells. In the absence of exogenous growth factors and/or small molecules, these inhibitors were able to maintain the expression of key pluripotency markers, including genes known to be specific for naive pluripotent state in mouse stem cells, for over 60 passages under feeder-free conditions. Surprisingly, the cells became dependent on HDAC inhibitors for the maintenance of proliferation. Moreover, despite showing successful integration into blastocysts upon injection, the cells were unable to undergo normal differentiation in vitro and in vivo in the form of teratomas. Our results suggest that HDAC inhibitors maintain pluripotency gene expression of porcine iPSC-like cells in long-term culture, but prevent lineage specification, requiring further optimization of culture conditions for porcine iPSC derivation.
Introduction
T
Due to the significant physiological, metabolic, and genetic similarities between humans and pigs, porcine iPSCs could be very useful as animal models for the development of regenerative therapies. In addition, the availability of porcine iPSCs would significantly benefit the generation of transgenic animals by exploiting the abilities of pluripotent cells to self-renew for unlimited periods of time without reaching senescence, thus allowing for multiple genetic modifications to be performed before being utilized in chimera production or in somatic cell nuclear transfer. Following the establishment of mouse and human iPSCs [9,10], several groups have reported on the derivation of porcine iPSCs using a variety of different culture conditions [11 –17]. Despite initially encouraging results, including the production of chimeras from mesenchymal stem cell-derived pig iPSCs [16], major hurdles remain that prevent application of these cells in preclinical models. The reported iPSC lines vary significantly in the expression of pluripotency markers and in their pluripotency abilities, reflecting differences in cell culture conditions used for their derivation [12,16,17]. Germ line-competent porcine iPSCs have not yet been reported [18]. Further optimization of cell culture conditions is necessary to achieve complete reprogrammed porcine cells that are equivalent to their mouse counterparts.
There have been very few reports on the use of HDAC inhibitors for derivation and culture of porcine iPSCs. In one report, supplementation with 1 mM VPA resulted in 2.7% increase in the number of alkaline phosphatase (AP)-positive clones in primary culture [11]. In another study, 1 μM VPA was used for the conversion of porcine iPSCs from primed to naive-like state [15]. In these reports, VPA was used only transiently for short periods of time together with other supplements such as growth factors and small molecule inhibitors. The effects of the HDAC inhibitors alone have not been determined in short- or long-term culture of porcine somatic cells after reprogramming. In this study, we investigated the effects of three HDAC inhibitors: NaB, VPA, and suberoylanilide hydroxamic acid (SAHA), on the proliferation potential of porcine reprogrammed cells and characterized the expression of critical pluripotency genes in putative porcine iPSCs cultured long term under feeder-free conditions.
Materials and Methods
Unless mentioned otherwise, the chemicals and kits used in this study were purchased from Sigma-Aldrich.
Establishment and culture of putative iPSCs
Porcine fetal fibroblasts (pFFs) carrying the mouse OCT4 promoter–enhanced green fluorescent protein reporter (O4-EGFP) construct have been established in our laboratory as reported previously [19]. Sleeping beauty (SB) transposon vectors for the expression of loxP-flanked porcine OCT4, SOX2, c-MYC, and KLF4 construct (SB-loxP-CAG-pOSMK-IRES-Tomato-loxP-pA) and porcine NANOG-human LIN28 (SB-loxP-Ef1a-pNANOG-IRES-hLIN28-loxP-pA) have been produced as described earlier [20]. Three pFF-Oct4-EGFP lines (each derived from a separate fetus) were cultured at early passage in a basic medium [DMEM high glucose (Lonza), 15% fetal bovine serum (FBS) (Lot 41Q2035K; Gibco),
Reverse transcription and polymerase chain reaction analysis of gene expression
Following disaggregation with trypsin, putative piPSCs were centrifuged, resuspended in TRI Reagent Solution (Ambion), and total RNA was extracted according to the manufacturer's manual. The purified total RNA was reverse transcribed with MuLV Reverse Transcriptase (Applied Biosystems). Polymerase chain reaction (PCR) was then carried out using 50 ng reverse-transcribed RNA per 25 μL reaction volume using Platinum Taq Polymerase (Invitrogen). Real-time relative quantitative analysis was performed on ABI 7500 Fast System using SYBR Green Master Mix (Applied Biosystems). The primers used for PCR and real-time PCR analysis of endogenous porcine, OCT4, SOX2, NANOG, c-MYC, KLF2, KLF4, KLF5, TERT, UTF1, EpCAM, ZFP42, CHD1, TDH, CDH1, ESRRB, and GAPDH, are shown in Supplementary Table S1. The primer sequences for KLF2, KLF5, and EpCAM were adopted from [21]. The sequences of the primers used for analysis of transgene expression have been shown earlier [20]. The data from the relative quantitative analysis were analyzed with GeneEx software (bioMCC). Statistically significant differences were determined by Student's t-test performed by the same computer program.
AP staining
AP staining solution was freshly prepared before each staining by suspending 1 mg/mL Fast Red TR and 0.4 mg/mL Naphthol AS-BI phosphate disodium salt in an AP buffer [100 mM NaCl2, 100 mM Tris (pH 9.5), and 50 mM MgCl2]. The cells were fixed for 5 min with 4% formalin, washed thrice with phosphate-buffered saline (PBS), and incubated with the staining solution for 15–20 min or until the development of red precipitate on the AP-positive cells.
Immunocytochemistry
The wells with putative porcine iPSCs were fixed with 4% formalin for 15 min, washed with PBS, and incubated with primary mouse antibodies raised against SSEA-1, SSEA3, SSEA-4, TRA-1–60, or TRA-1–81 (BioLegend) suspended in PBS, supplemented with 2% Knockout Serum Replacement (Life Sciences) at 10 ng/mL for 1 h at room temperature. For staining of neuronal-like cells, a similar procedure was applied using anti-PGP 9.5 primary antibody (Daco). Following washing with PBS, the cells were incubated with peroxidase-conjugated secondary anti-mouse IgM or IgG antibody diluted 1:1,000 for 30 min. After washing thrice with PBS, the bound antibody was visualized with peroxidase substrate solution prepared using an AEC Staining Kit as instructed by the manufacturer.
Embryo contribution assay
To produce parthenogenetic blastocysts, slaughterhouse-collected oocytes were matured in vitro, activated as described previously [22], and then cultured in PZM3 for 5 days. Expanding blastocysts were injected with four to six OCT4-EGFP-positive cells from Group A at P40-43 and cultured in PZM3 or PZM3: piPSC medium (containing deacetylase inhibitors) mix at ratio 3:1. Three days postinjection, the blastocysts were observed under UV light and photographed using inverted Leica DM IRB microscope equipped with Nikon DS-Fi1c camera. In addition, some embryos were washed thrice in PBS with 0.5% FBS and 1% penicillin/streptomycin and directly fixed in 4% paraformaldehyde for 15 min at room temperature. Thereafter, nuclei were stained with NucRed™ Live 647 (R37106; Life Technologies) for 30 min at room temperature. Then, embryos were transferred on a glass slide within a droplet of PBS and examined immediately using a confocal imaging system LSM510 with an Axioplan 200 (Carl Zeiss Micro Imaging GmbH), using a 40× Plan Apochromat objective.
In vitro differentiation
To see whether the piPSCs were able to differentiate spontaneously in the absence of HDAC inhibitor supplementation, we split and cultured putative piPSCs at P15, 20, or 30 in a basic medium without HDAC inhibitors. To attempt neuronal differentiation, the piPSCs were disaggregated to single cells with trypsin, cultured in hanging drops (2,000 cells/30 μL drop) to form embryoid body (EB)-like aggregates for 7 days in iPSC medium with or without HDAC inhibitors. The viability of the cells in hanging drop culture where the medium was not supplemented with HDAC inhibitors was monitored every day by plating cells from several drops on gelatinized plastic wells with an HDAC inhibitor-supplemented piPSC medium. After 7 days, cell aggregates were cultured further on gelatin-coated plastic wells in a neuronal differentiation medium [DMEM/F12 supplemented with sodium pyruvate, amino acids, penicillin–streptomycin, and N2 supplement (Thermo)] for up to 2 weeks. To improve the survival, the differentiation medium in some wells was further supplemented with HDAC inhibitors.
Previously isolated piPSC-like cells derived in standard culture conditions [20] as well as miPSCs derived from mouse fetal fibroblasts by transfection with the SB-loxP-CAG-pOSMK-IRES-Tomato-loxP-pA and cultured on STO feeders in a medium supplemented with 1,000 U/mL ESGRO (Thermo) were differentiated under the same protocol and served as positive controls.
Teratoma assay
Before conducting the animal experiments, permission was obtained from the local animal welfare authority (LAVES).
Nude immunodeficient mice (n = 13) produced by mating of outbred NMRI strain parents were injected each with 1 × 106 putative porcine iPSCs from Group A at P45-47 subcutaneously in the right flank. For positive control, additional six mice were injected each with 1 × 106 mouse iPSCs derived from mouse fetal fibroblasts by transfection with the SB-loxP-CAG-pOSMK-IRES-Tomato-loxP-pA transposon and cultured on STO feeders with ESGRO. The mice were observed daily for 10 weeks. Having reached the critical size of the tumor, the animals were sacrificed. After taking biopsies for DNA/RNA extraction, tissues were fixed in neutral-buffered 4% paraformaldehyde not exceeding 48 h, dehydrated (Shandon Hypercenter, XP), and subsequently embedded in paraffin (TES; Medite). Sections (2–3 μm thick, microtome Reichert-Jung 2030), were deparaffinized in xylene and H&E stained according to standard protocols. Morphologic evaluation for the three germ layers (Axioskop 40; Zeiss microscope) was performed by a trained pathologist and representative microphotographs were taken (AxioCam MRc; Zeiss).
Results
Derivation and long-term culture of porcine iPSCs
All three pFF-O4-EGFP lines transfected with SB transposons and SB100× transposase formed EGFP-positive cell clusters within 8 days in culture on STO feeders, indicating the reactivation of the OCT4 gene. By day 12 of primary culture, these cells were organized in compact flat colonies with defined borders (Fig. 1A, B). After manual picking and disaggregation in trypsin, individual colonies continued to proliferate in SAHA, VPA, NaB, and AA-supplemented medium and formed clonal lines with an average 76% efficiency (n = 12 colonies picked from each transfected pFF-O4-EGFP line). Three putative porcine iPSC lines with compact iPSC-like morphology and strong EGFP fluorescence were chosen for long-term culture in different conditions as defined in the Materials and Methods section for Groups A and B.

Morphology of putative porcine iPSCs.
During prolonged culture (>15 passages), the feeder-free culture resulted in the formation of flat colonies where the cells were epithelial-like and EGFP positive (Group A; Fig. 1C, D), or assumed fibroblast-like morphology and lost their EGFP expression (Group B; Fig. 1E, F). The Group A cell lines have been maintained for over 90 passages and during this time, one cell line maintained the expression of the O4-EGFP reporter in all cells, while the other two lines lost EGFP fluorescence after P45/50. The proliferation in all cell lines was dependent on the transgenes, which were downregulated, but not completely silenced [as evidenced by weak Tomato fluorescence and confirmed by reverse transcription polymerase chain reaction (RT-PCR) analysis; results not shown]. When the transgenes were excised by expression of Cre-recombinase, the proliferation stopped completely.
After ∼40–45 passages, the putative iPSCs in Group A also became dependent on the HDAC inhibitors for proliferation, and withdrawal of the inhibitors resulted in remarkable slowing of the cell growth (EGFP-negative cell lines) and significant cell death within a few days (EGFP-positive line). By conducting differential experiments, it was determined that each inhibitor alone was able to maintain the cells in culture, although the iPSCs showed a decreased proliferation rate when cultured only with VPA compared to NaB, SAHA, or all three inhibitors together (Supplementary Fig. S1; Supplementary Data are available online at
Pluripotency gene expression
Gene expression analysis at P20, P40, and P60 showed that the feeder-free cells from Group A expressed pluripotency surface markers AP and SSEA-1, but not SSEA-3, SSEA-4, TRA-1–60, and TRA-1–81 (Supplementary Fig. S2). Furthermore, gene expression analysis by RT-PCR showed that these cells expressed endogenous OCT4, SOX2, NANOG, TERT, ZFP42, UTF1, CHD1, c-MYC, KLF4, KLF5, EpCAM, ESRRB, CDH1, and TDH (Fig. 2A). By using specific primers (shown on Supplementary Table S1), it was verified that the cells expressed the stem cell-specific isoform A of endogenous porcine OCT4 at P20 as well as P60 (Fig. 3A). Real-time PCR analysis revealed that most of the pluripotency markers were significantly upregulated compared with the pFFs, with the exception of KLF4, which did not show a statistically significant difference and c-MYC, which was downregulated in putative iPSCs relative to pFFs (Fig. 2B). KLF2 was detected at very low levels in both pFFs and putative iPSCs (real-time PCR showed Ct values bigger than 35 cycles).
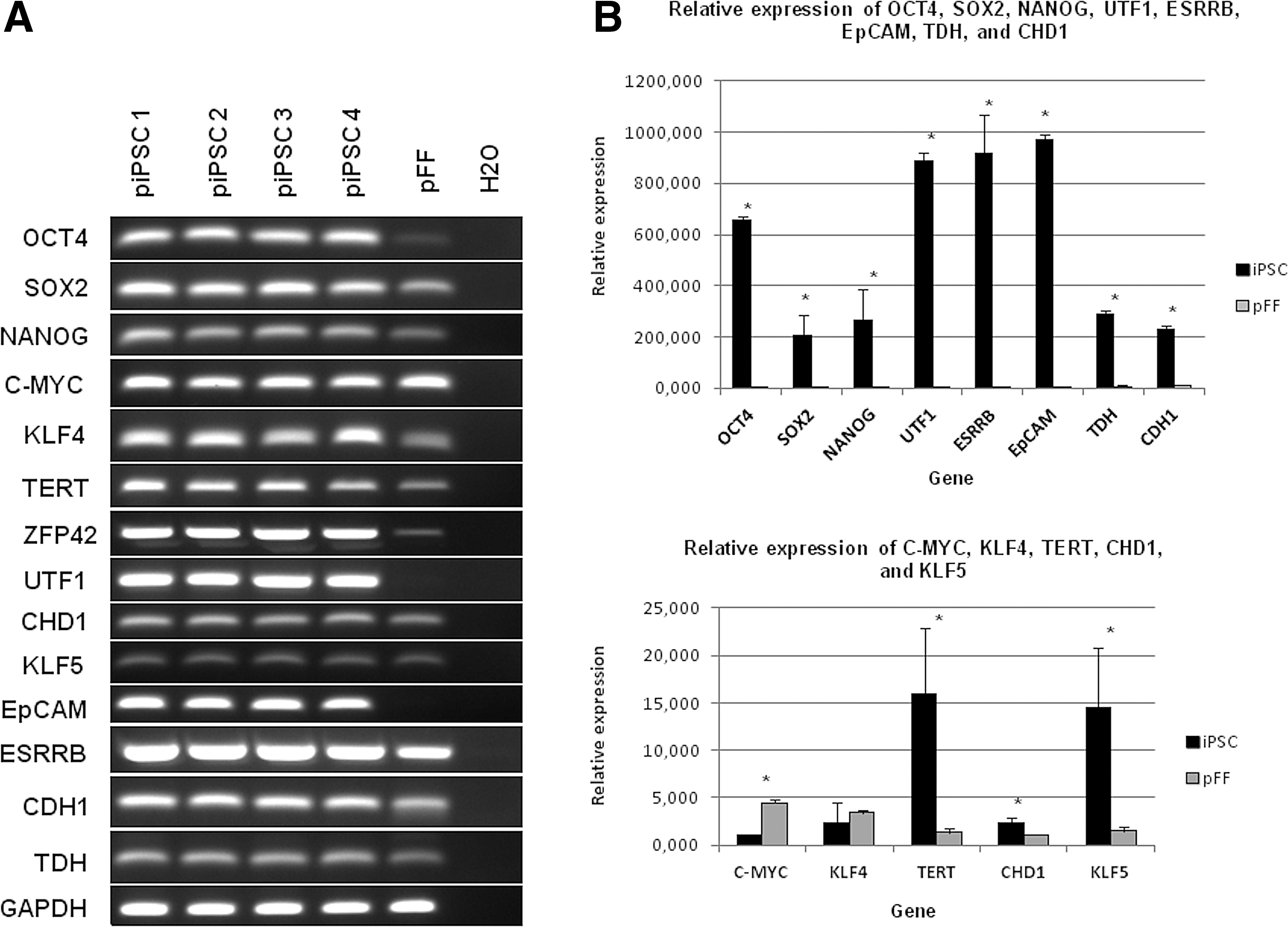
Pluripotency gene expression in putative porcine iPSCs derived with HDAC inhibitors and AA and cultured in feeder-free conditions for 20 passages.
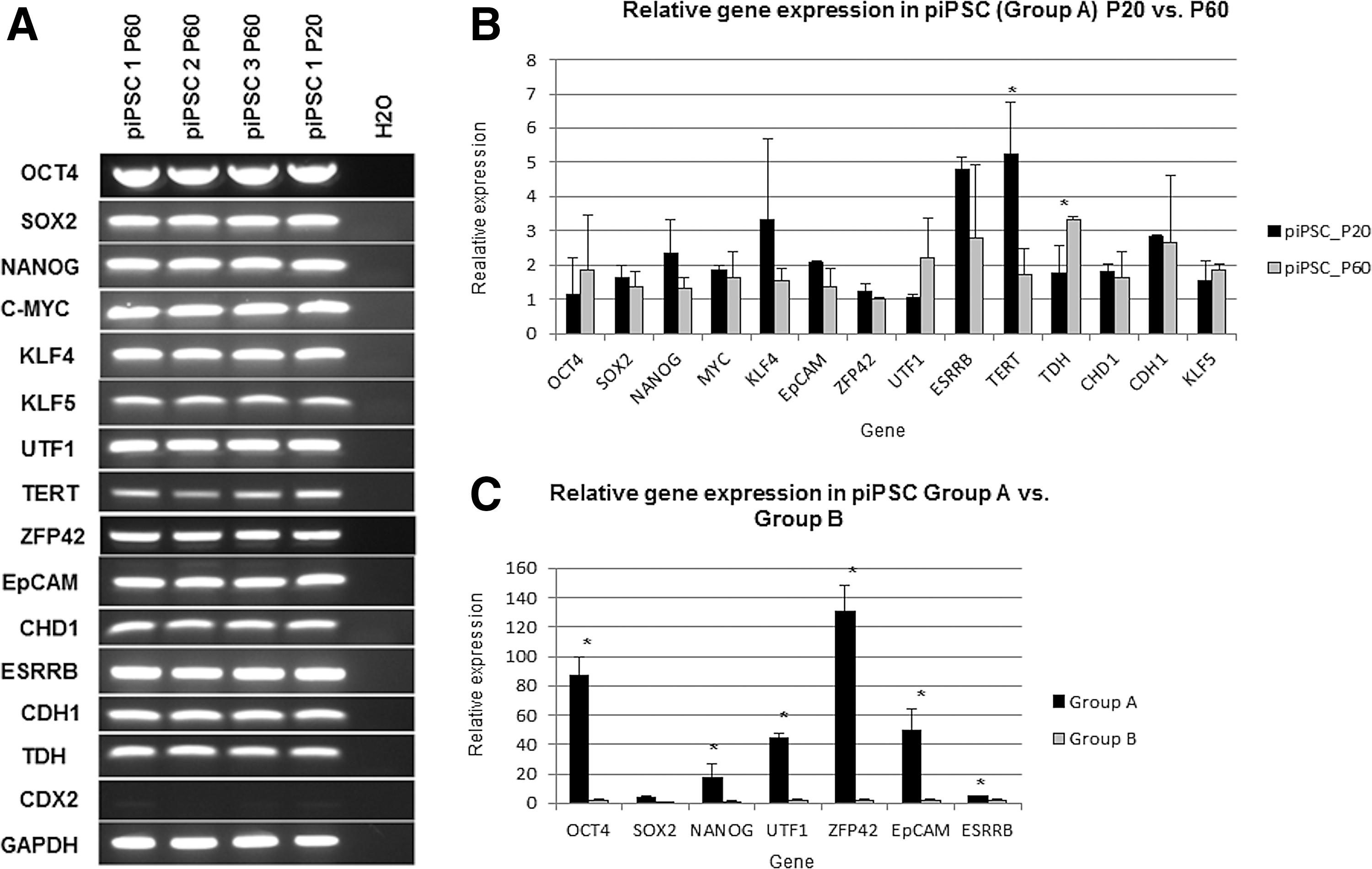
Gene expression of putative porcine iPSCs after 60 passages in feeder-free conditions with HDAC inhibitors and AA.
The ability of the HDAC inhibitors to maintain expression of pluripotency genes during long-term feeder-free culture was investigated by comparing pluripotency gene expression at P20 and P60 using RT-PCR (Fig. 3A) and real-time relative quantitation analysis (Fig. 3B). Most genes did not show significant differences, with the exception of TERT, which was significantly downregulated at P60, while at the same time, TDH was significantly upregulated (Fig. 3B). It was noted that despite loss of O4-EGFP reporter fluorescence over time, the affected cell lines continued to express the endogenous isoform A (Fig. 3A, piPSC lines 2 and 3) and there was even a small insignificant overall increase in the expression of this gene at P60 relative to P20 (Fig. 3B).
After feeder-free culture for 55 passages, the relative expression levels of key pluripotency markers OCT4, NANOG, UTF1, ZFP42, EpCAM, and ESRRB were significantly reduced in Group B compared to Group A (Fig. 3C; see also RT-PCR gel image in Supplementary Fig. S3), suggesting that ESGRO and AA alone were not sufficient to maintain the pluripotency characteristics of the cells in the absence of feeders.
At passage 60, three cell lines from Group A were also compared with piPSC-like cells established and maintained in conventional conditions [20]. Relative quantitative real-time RT-PCR revealed no significant difference in the expression levels of OCT4, SOX2, NANOG, KLF4, TERT, UTF1, TDH, CHD1, CDH1, and ESRRB (all P values >0.05). On the other hand, c-MYC was expressed significantly higher in the conventional group (P < 0.002), while EpCAM was expressed at a higher level in Group A (P < 0.05) (Supplementary Fig. S4).
Contribution to embryo development
Injection of four to six EGFP-positive cells at P40-43 into day 5 parthenogenetic blastocysts resulted in the integration of the cells into the trophectoderm (Fig. 4A, B) as well as in the inner cell mass (ICM) (Fig. 4E). While only 3.7% of the injected blastocysts cultured in PZM3 had surviving EGFP-positive cells by day 8, 53% of the embryos cultured in the PZM3: piPSC medium mix showed incorporated cells. Few blastocysts that were accidentally injected with 10 or more cells showed integration of larger number of cells; however, these embryos lagged behind the rest in their growth and did not expand significantly after day 6 (Fig. 4C, D). When the blastocysts containing putative piPSCs were cultured on feeders, they attached and formed outgrowths of EGFP-positive colonies with iPSC-like morphology within a week, confirming that the cells were viable.
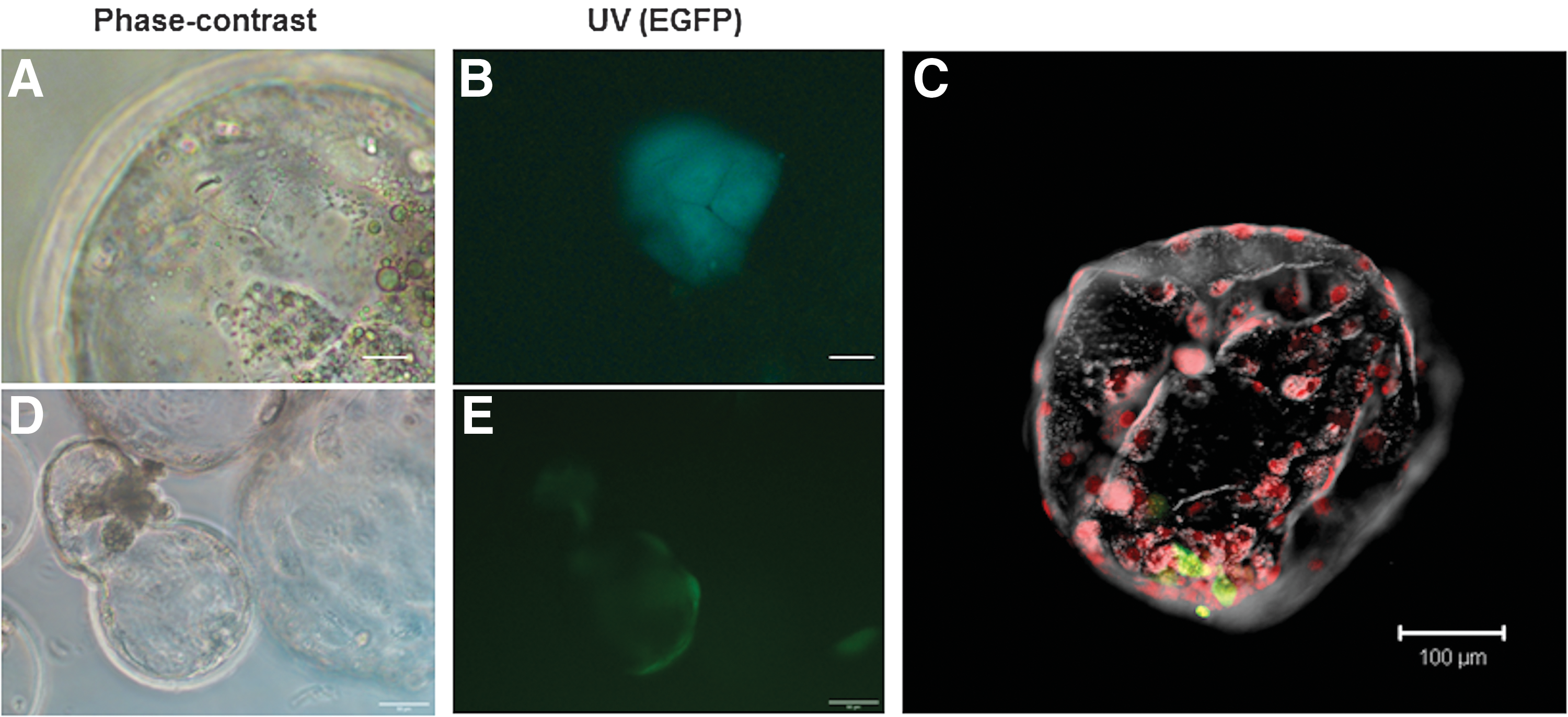
Contribution of putative porcine iPSCs to parthenogenetic blastocysts, day 3 postinjection.
In vitro differentiation
Regardless of the culture conditions used, the putative piPSCs were not able to differentiate into other cell types. When cultured directly in a differentiation medium without HDAC inhibitors, mass death was observed within 4 days. Hanging drop culture without HDAC inhibitors resulted in cell death after day 3–4 and the cells failed to form aggregates. When the medium was supplemented with HDAC inhibitors, the putative piPSCs formed small clumps (Supplementary Fig. S5A), unlike piPSC-like cells derived in standard conditions that were able to form EB-like aggregates (Supplementary Fig. S5B). Further culture of the cell clumps in the N2 medium without HDAC inhibitors lead to successful attachment, but the cells did not expand and differentiate (Supplementary Fig. S5C), while culture with these inhibitors improved the survival, but did not allow for differentiation into neurons (Supplementary Fig. S5D). Embryoid bodies from positive control miPSCs and piPSCs derived with feeders and ESGRO successfully differentiated into neuronal-like cells that reacted positively with anti-PGP 9.5 antibody (Supplementary Fig. S5E, F).
Teratoma assay
One EGFP-positive/HDAC-dependent and one EGFP-negative/HDAC-independent line was tested for teratoma formation. While the EGFP-positive line did not form any tumor outgrowths in nude mice, the EGFP-negative cells formed an undifferentiated tumor composed of only one cell type and multiple blood vessels (supposedly originating from the host) (Fig. 5) with 33% efficiency. Analysis of pluripotency gene expression by RT-PCR showed that all endogenous pluripotency genes as well as the transgenes were still expressed (results not shown). The miPSCs injected as positive controls formed well-differentiated teratomas with 100% efficiency (results not shown), confirming that the injection procedure was effective.

Histology of nondifferentiated tumor generated by injection of putative piPSCs maintained with HDAC inhibitors into immunodefficient mice.
Discussion
Despite years of extensive research, the optimal culture conditions for derivation and long-term maintenance of true pluripotent porcine iPSCs could not be yet determined. Reports on the needs of porcine iPSCs for specific growth factors are conflicting as some groups have shown that some piPSC lines have functional JAK/STAT3 signaling pathway and may develop leukemia inhibitory factor (LIF)-dependency after prolonged culture [23,24], while others have isolated porcine iPSCs without using this growth factor [11,16,17,21,25,26]. Evidence of chimera contribution was shown using bFGF, but not LIF supplementation [16]. Unfortunately, these results have not been reproduced to date, and some studies have shown that despite meeting most pluripotency criteria, porcine iPSCs failed to produce live-born chimeras [14,27]. These results demonstrate that to date it remains unclear how to derive germ line-competent iPSCs equivalent to mouse or rat iPSCs.
In the majority of the published studies, piPSCs have been maintained over unspecified or relatively short periods of time in culture conditions typically used for mouse or human iPSCs/ESCs. These conditions have so far proven inadequate for the derivation of fully pluripotent piPSCs or ESCs, particularly in feeder-free conditions; therefore, the current situation necessitates the testing of novel methods and culture conditions for the isolation of piPSCs that have not been yet used for human or mouse pluripotent cells. In our trial-and-error experiments for further culture optimization, we investigated whether HDAC inhibitors would be beneficial for maintaining the pluripotency of porcine iPSCs. To our knowledge, the effects of the culture conditions used in our experiments on the pluripotency characteristics of iPSCs have not been yet reported in other species. Surprisingly, we found that SAHA, NaB, and VPA were able to maintain relatively high levels of expression of key pluripotency markers, including genes that are considered markers of naive pluripotent state in mouse ESCs/iPSCs, without need for feeder cells and further supplementation with growth factors and/or small molecule inhibitors. This effect could not be reproduced with ESGRO and AA in the absence of feeders. Compared with previously derived porcine iPS-like cell lines cultured in the presence of LIF and feeders [20], the cells cultured with HDAC inhibitors in feeder-free conditions showed no significant differences in the expression levels of the key pluripotency markers. On the other hand, the new conditions did not improve the differentiation competence of the putative piPSCs, suggesting that the use of HDAC inhibitors does not provide any advantage over the conventional culture conditions other than allowing the cells to be cultured in the absence of feeders and growth factors, which significantly reduces the costs and the amount of work required for their maintenance.
When HDAC inhibitors were applied in long-term feeder-free culture, pluripotency genes were continuously expressed for over 60 passages without significant changes of the expression levels of key pluripotency genes such as OCT4, NANOG, ZFP42, UTF1, EpCAM, and ESRRB. By comparing global expression profiles of mouse, human, and porcine iPSCs, it was shown that EpCAM and ZFP42 correlate with pluripotency and were suggested as suitable markers to evaluate reprogramming in porcine iPSCs [21]. On the other hand, upregulation of genes from the KLF group in our cells was incomplete. Only KLF5 showed a small, but significant increase, while endogenous KLF4 was expressed at a lower level compared with pFFs, although the difference was statistically not significant. In addition, KLF2 expression was detected only at negligible levels. These results suggest that in future work, activation of endogenous KLF2/4/5 expression might be a worthwhile direction to pursue.
Similar to the vast majority of previous reports on porcine iPSC derivation [16,17,25,28], the exogenous reprogramming factors in putative porcine iPSCs were not completely silenced and the cells were not able to proliferate or maintain an undifferentiated profile in the absence of exogenous transcription factors. This could indicate incomplete reprogramming, as suggested by Du et al. [27], likely due to the fact that the culture conditions used are still not sufficiently optimal for porcine iPSCs. Similar results were reported recently regarding mouse iPSCs produced by using elevated levels of nonsilenced, transposon-delivered, reprogramming transcription factors [29]. The iPSCs derived in this study were characterized by aberrant morphology and dependency on sustained expression of transgenes for their maintenance. Moreover, despite demonstrating stable endogenous pluripotency marker expressions, these cells failed to produce chimeras and were classified as intermediate state (or F-class state) iPSCs (29). The results from these experiments imply the existence of a spectrum of distinct pluripotency-like states. Based on the data presented in the published literature, it can be suggested that the majority of the reported pig iPSC lines can be included in this category.
An unexpected outcome of our experiments was that the cell lines became dependent at various degrees on HDAC inhibitors for their proliferation. This was particularly well pronounced in the iPSC-like line that has retained the O4-EGFP expression for over 90 passages until today. This problem may have prevented the formation of teratomas in nude mice by this cell line since the cells need to survive and expand to form tumors. During our embryo integration experiments, mixing the embryo culture medium with porcine iPSC medium had positive effects on the cell survival and integration, which suggests that at least some of the HDAC inhibitors could help with the initial survival of the piPSCs during the early steps of in vivo differentiation. Nevertheless, providing these supplements to the cells inside the experimental animals would be a very difficult task, as demonstrated by the rapid uptake and metabolism of NaB, resulting in very low blood concentration [30].
Another more serious problem that was encountered was the inability of the iPS-like cells to undergo differentiation in vitro and in vivo. All three tested cell lines did not survive in the neuronal differentiation medium without HDAC inhibitors, while in the presence of these compounds there was no change in their differentiation status. This problem, together with the inability of the cells to form differentiated teratoma, could be due to the prolonged exposure to HDAC inhibitors, resulting in the accumulation of unfavorable histone acetylation patterns that stabilize the cells in pluripotency-like state. Therefore, in future work it would be necessary to optimize culture conditions where the cells can be adapted to proliferate without HDAC inhibitors following derivation or at certain time points before differentiation.
In conclusion, we successfully established and maintained putative piPSCs in growth factor- and feeder-free culture in medium supplemented only with three HDAC inhibitors and AA. We demonstrated that these inhibitors could maintain the expression of key pluripotency genes, including some markers for naive pluripotent state, in long-term culture without the need for supplementation with growth factors and/or small molecule inhibitors. Similar to other reports, the porcine iPSC-like cells were dependent on continuous expression of the transgenes for their maintenance and despite the expression of key stem cell markers could not fulfill all the criteria for pluripotency. Thus, the cells were similar to recently reported mouse iPSCs which showed an alternative state of pluripotency, the F-state. Despite the unexpected findings stemming from the long-term use of HDAC inhibitors, the insight that these inhibitors can be applied to maintain pluripotency marker expression in the absence of feeders and growth factors, at least for a certain time period during reprogramming of porcine cells to iPSCs, will help to further improve the conditions for the derivation of porcine iPSCs.
Footnotes
Acknowledgments
We thank Dr. Wilfried Kues (Friedrich-Loeffler-Institute, Mariensee, Germany) for the injection of the immunodeficient mice used for the teratoma assay, Dr. Zoltan Ivics (Paul-Ehrlich Institute, Langen, Germany) for providing the SB transposon vector backbone and the SB transposase expression vector, and Dr. Poul Hyttel (University of Copenhagen, Denmark) for allowing us to use and modify the transposon vectors constructed in his laboratory. In addition, we are grateful to Dr. Sabine Klein [Institute for Farm Animal Genetics (FLI), Neustadt, Germany] for the technical help with confocal microscope imaging. This study was supported by the German Research Foundation (Grant Ni 256/32–1).
Author Disclosure Statement
No competing financial interest exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.