
Abstract
Differentiation and development are normally unidirectional processes in which progenitor/stem cells differentiate into more mature cells. Transformation of adult cells into cancer cells is accompanied in many cases by dedifferentiation of the adult cell, while differentiation failure of progenitor cells can result in the formation of unique type of cancers called pediatric cancer. LIN28A and its paralog LIN28B are pluripotent genes that are expressed mainly in stem/progenitor cells. Since the first identification of LIN28 in mammals, numerous studies demonstrated the general oncogenic features of these genes. In this review, we emphasize the unique role of LIN28 in pediatric tumor formation. We show, based on comprehensive literature screen and analysis of published microarray data, that LIN28 expression in pediatric tumors is even more common than in adult tumors, and discuss the possibility that in the case of pediatric cancers, LIN28 acts by preventing normal development/differentiation rather than by transformation of mature cells into cancer cells. Overall, this review highlights the role of LIN28 as a bridge point between embryonic development, stem cell biology, and cancer.
P
These similarities between cancer cells and stem/progenitor cells suggest that cancer can arise also from a subpopulation of undifferentiated cells or cells that fail to complete their development process [6 –9]. Indeed, unique type of cancers called pediatric or childhood tumors are thought to be driven by the normal development mechanisms gone awry [10,11] (Fig. 1).
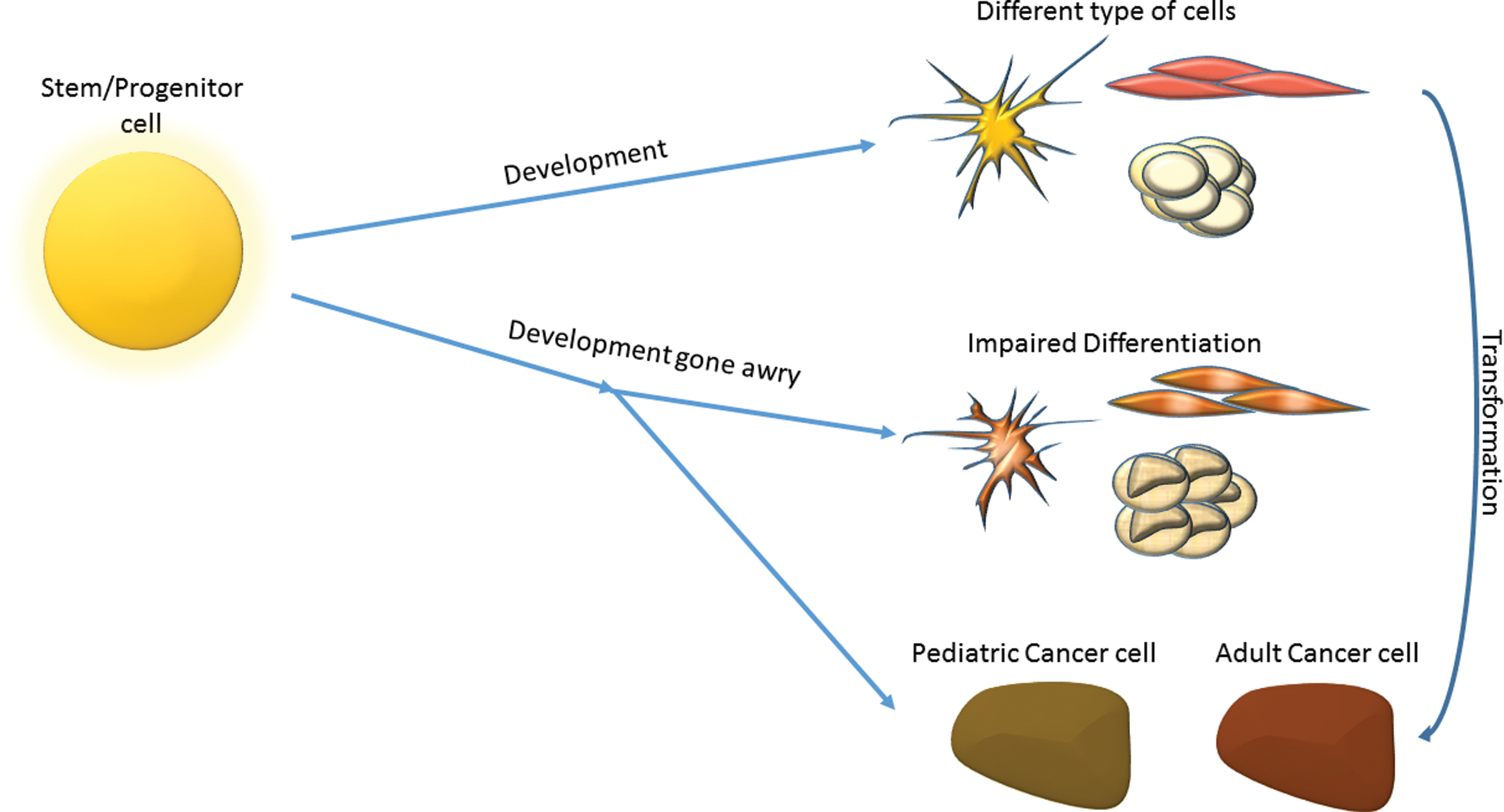
Schematic representation of pediatric cancer formation. During development stem/progenitor cells differentiate to various cell types (top right). Differentiated cells may transform to adult cancer cells. When the developmental process goes awry, it can lead to impaired differentiation of cells (middle right) or to pediatric cancer (bottom right). Color images available online at
lin28
The highly conserved RNA-binding protein, LIN28, was first identified in Caenorhabditis elegans as a heterochronic gene, a gene that alters the developmental timing or rate when dysregulated [12]. In mammals LIN28 is expressed mainly in stem and progenitor cells [13,14]. Furthermore, it was found that LIN28, along with OCT4, SOX2, and NANOG, successfully reprograms fibroblasts to induced pluripotent stem cells [15]. LIN28A, and its mammalian paralog LIN28B, regulate gene expression by two mechanisms: (1) selective prevention of pri-LET-7 microRNA maturation [16] and (2) direct targeting of a large number of mRNAs and thereby their translation modulation [17]. Utilizing these two mechanisms, LIN28 plays an important role in normal embryonic development [18,19], glucose metabolism [20], and, interestingly also, tissue repair [21].
The microRNA LET-7 family regulates the expression of several oncogenes such as MYCN, KRAS, and HMGA2. Therefore, LET-7 suppression by LIN28 may result in overexpression of these oncogenes and can eventually lead to tumor formation or to maintenance/advancement of the cancer to a more severe stage. Indeed, Viswanathan et al. were the first to report LIN28 abundance in advanced human malignancies [22]. The number of reports about LIN28-expressing tumors (through LET-7 dependent as well as through independent mechanisms [23]) is consistently growing and the subject is summarized by several excellent reviews [24,25]. Aside from LIN28 overexpression, single-nucleotide polymorphisms in LIN28 gene are considered to be a risk factor for multiple cancers [26 –29].
In this review we will focus on the unique role of LIN28 in pediatric cancer. For this purpose, we extend the list of reports on tumors overexpressing LIN28 (Table 1). This table reflects the recently published studies about LIN28-expressing tumors (both adult and pediatric) and allowed us to compare the frequency of LIN28 expression between adult and pediatric cancers.
AT/RT, atypical teratoid–rhabdoid tumor; HCC, hepatocellular carcinoma; N/A, not available; N/E, not expressed.
LIN28 Involvement in Pediatric Cancer
Pediatric cancers are defined as cancers that occur during childhood (age 0–15 years). These rare types of cancers arise in many cases as a result of impaired cellular development/maturation in the normal process of tissue development [30]. These cancers are divided between blood cancers (leukemias and lymphomas), that account for nearly half of the cases, and solid tumors [31]. Pediatric solid tumors represent a wide range of neoplasms, including tumors of brain, central nervous system, neuroblastoma, Wilms' tumor, bone tumors, soft tissue sarcomas, retinoblastoma, hepatoblastoma, germ cell tumors (GCTs), and other rare pediatric solid tumors [10].
An interesting observation we made from our literature survey (Table 1) is that about 25% of the cancers reported to overexpress LIN28 are pediatric cancers. This frequency is very high considering pediatric cancers represent less than 1% of all newly diagnosed cancers [31]. Moreover, we analyzed several published microarray studies (Barretina Cell Line, Wooster Cell Line,

LIN28 distribution among pediatric and adult cancers.
We suggest that the common expression of LIN28 in pediatric cancers points to a unique role of this gene in the formation of these cancers. We hypothesize that in the case of pediatric cancer formation, the dominant effect of the heterochronic gene LIN28 is by preventing normal development/differentiation, as opposed to its regular oncogenic role in adult cancers.
In this study, we summarize the main findings from selected articles shown in Table 1 as well as other articles that support the abovementioned hypothesis.
Wilms' Tumor
Wilms' tumor is the most common pediatric kidney cancer that originates from undifferentiated embryonic nephrogenic cells [32]. Renal-specific LIN28 overexpression during mouse embryonic kidney development leads to the formation of a tumor similar to human Wilms' tumor in a LET-7-dependent manner. The pathophysiological process that leads to the tumor formation in this mouse model consists of sustaining cap mesenchymal (CM) cells, which normally exist only until postnatal day 2 [33]. Importantly, induction of LIN28 overexpression in mature renal tissue when the CM cells have already differentiated, fails to form tumor.
In accordance with the mouse data, in humans, LIN28B was shown to be overexpressed in the most undifferentiated cells of Wilms' tumor in around 20% of the cases [32]. Collectively, these observations may imply that abnormal activation of the LIN28-LET-7 pathway prevents normal kidney development and might be one of the factors that leads to Wilms' tumor formation in human.
Neuroblastoma
The pediatric tumor neuroblastoma forms from the neural crest-derived cells in which the maturation process was arrested at different stages of differentiation [10,34]. The tumor appears mainly in one of the adrenal glands, but can also originate in other tissues of the nervous system [34]. A strong linkage between LIN28B and neuroblastoma was found by a genome-wide association study [27]. Based on this observation, the effect of LIN28B knockdown on the growth of neuroblastoma cell lines was examined and a significant decrease in growth rate was detected [27]. In agreement, a strong association between high LIN28B expression levels and lower overall survival was found. While these results indicate that LIN28B plays a role in the progression of neuroblastoma, they do not necessarily point to a role of LIN28 in the formation of the tumor.
To explore the contribution of LIN28B to the formation of the tumor, Molenaar et al. [35] overexpressed LIN28B in cells of the sympathoadrenergic lineage of the neural crest during mouse embryonic development. This led to neuroblastoma formation in 25% of the cases and demonstrated the ability of LIN28B to initiate neuroblstoma. The authors further searched for downstream targets of LIN28B and concluded that LIN28B blocks the differentiation of normal neuroblasts and neuroblastoma cells through LET-7-MYCN pathway. Interestingly however, the interplay between LIN28B and MYCN in neuroblastoma is more complicated as Beckers et al. showed recently that MYCN and LIN28B reciprocally regulate each other [36]. Indeed, in a computational modeling study, it has been shown that high levels of C-MYC can maintain high levels of LIN28 [37]. In addition, neural progenitor cell proliferation and differentiation was shown to be affected by LIN28 [23], further linking its involvement in neuroblastoma.
Finally, a recently published article by Rohrer and colleagues suggest that, by contrast to the conclusions of previous studies, in the case of LIN28B-derived neuroblastoma, LIN28B acts in a LET-7-independent manner and the tumor founder cells are not embryonic neuroblasts [38]. Thus, the specific cell type affected by LIN28B overexpression in the case of neuroblastoma is still yet to be determined.
Pediatric Hepatoblastoma and Hepatocellular Carcinoma
Pediatric hepatoblastoma is a rare liver solid tumor in children. The molecular characteristics of pediatric hepatoblastoma are similar to those of hepatocellular carcinoma (HCC) [39]. However, the causes of these tumors are different. While HCC usually arises as a result of extrinsic interference to the liver tissue, including injury, hepatitis B, hepatitis C, etc. [40], hepatoblastoma arises as a result of intrinsic factors such as mutation in the APC gene or faulty imprinting of the IGF2-H19 locus [41].
In a recently developed mouse model it was shown that overexpression of LIN28B during embryogenesis causes hepatoblastoma and HCC [9]. In this model, the expression of fetal liver markers, such as α-fetoprotein and insulin-like growth factor 2, as well as widespread overexpression of imprinted genes, indicate the fetal stage of the tumor cells. One plausible explanation for this phenomenon is that LIN28B prevents normal differentiation of hepatocyte cells. However, in this case, the other possibility of dedifferentiation by LIN28B cannot be ruled out [9]. It was further shown that LIN28 is important also for the maintenance of the tumor and that in the mouse model for liver cancer, LIN28 acts through LET-7-dependent and LET-7-independent mechanisms [9].
With respect to humans, HCC is a well-studied cancer and it has been reported to express LIN28A [42,43] as well as LIN28B [44]. In contrast, to the best of our knowledge, there are no published reports on LIN28 expression levels in patients with hepatoblastoma. Although, when we analyzed microarray data from Oncomine (Barretina Cell Line), we found that LIN28A, as well as LIN28B, are expressed in this type of tumor.
Germ Cell Tumors
GCTs consist of heterogeneous neoplasms that can occur across the body. This distribution is seemingly due to the migration of primordial germ cells during embryogenesis [45]. Type II GCTs are considered to be developmental tumors since they are derived from primordial germ cells that fail to complete their development into mature germ cells [45]. Germ cell formation was one of the first developmental processes where the importance of LIN28A was experimentally demonstrated [46]. Both LIN28A and LIN28B contribute to germ cell differentiation through LET-7 repression in vitro, however, in vivo assays show that LIN28A is a more dominant player in this event [46]. Pediatric type II GCTs overexpress LIN28A and LIN28B [47,48]. Collectively, the role of LIN28 in germ cell development and the expression of LIN28 in GCT suggest that abnormal overexpression of LIN28 might dysregulate the growth or differentiation/development of germ cells [46].
In contrast to type II GCT, spermatocytic seminoma (a common type III GCT in adults) and teratomas (well differentiated type I GCT) were negative for LIN28 expression [47]. Knockdown of LIN28 revealed that it is essential for the maintenance of the undifferentiated state of the GCTs [47]. As expected, ectopic expression of LET-7 sufficiently reduced the LIN28-induced proliferation in GCT (through MYCN repression, similar to neuroblastoma) [48].
Rhabdoid Tumor
Rhabdoid tumor of the kidney and atypical teratoid–rhabdoid tumor (AT/RT) of the brain are among the most fatal pediatric cancers [49 –51]. The solely genetic aberration found in these tumors is a homozygous mutation in SNF5, a chromatin-remodeling gene, thus these tumors are considered to be “epigenetic tumors.” In more than 75% of primary AT/RTs, LIN28A or LIN28B or both are overexpressed [52]. We also found the expression of LIN28A and LIN28B in rhabdoid tumor of the kidney based on the published gene expression data in Oncomine and GEO (Barretina Cell Line, Wooster Cell Line, Neale Multi-cancer, GSE11151).
LIN28A contributes to growth and proliferation of the tumor by reducing apoptosis through LET-7–KRAS–phosphoERK pathway [52]. Injection of LIN28A knockdown AT/RT cells into brain of immunodeficient mice resulted in median survival extension compared to injection of LIN28A-expressing AT/RT cells (115 days compared to 48 days, respectively). Nevertheless, although the mice injected with the LIN28A knockdown cells lived longer, there were no differences between the expression levels of LIN28A in both tumors and the tumors were of equal size at the end of the experiment. This indicates the aggressiveness of cells that escaped from LIN28A knockdown and formed the tumor [52].
Taken together, these results point to the role of LIN28 in the maintenance and aggressiveness of the tumor. Yet, we suggest that the expression of LIN28 in these pediatric tumors indicates that this protein may prevent normal development of the brain and the kidney (such as the case in Wilms' tumor) resulting in rhabdoid tumor formation. It will be interesting to study what is the exact interplay between LIN28 overexpression and SNF5 loss of function in these tumors.
LIN28A Versus LIN28B in Cancers
The main focus of this review was to demonstrate and discuss the abundance and involvement of LIN28 in pediatric cancer formation. In doing so, our comprehensive published microarray data analysis raised an intriguing difference between LIN28A and LIN28B. In most of the published articles (Table 1), only one of the LIN28 paralogs was tested, either LIN28A or LIN28B, and there was no data about the other paralog. Only a few studies tested both paralogs. Out of these, both LIN28A and LIN28B were found to be expressed in AT/RT [52], HCC [53], colon cancer [54], esophagus cancer [55], oral squamous cell carcinoma [56], ovarian cancer [57,58], and GCT [48]. By contrast, adrenocortical cancer [59] expresses only LIN28A and Wilms' tumor expresses only LIN28B [33] (Table 1).
Overall, we did not notice any dominance between LIN28A or LIN28B among the different studies. Nevertheless, when we compared adult versus pediatric cancers in the microarray published studies, the distribution of LIN28A and LIN28B differed. We found that LIN28B is much more common in cancers than LIN28A (Fig. 2A). This tendency was even more pronounced in pediatric tumors where we could not find even a single tumor that expressed only LIN28A. To verify this finding, we examined additional microarray expression data focusing on pediatric cancers only (Neale Multi-cancer,
These differences between LIN28B and LIN28A can be explained either at the functional level (the oncogenic capacity of LIN28B is stronger than that of LIN28A) or at the regulational level (LIN28A and LIN28B have the same potential to cause tumors, but ectopic expression of LIN28B is much more common than that of LIN28A). While the overall domain structure of these two proteins is very similar [60], there is still some diversity between these proteins that may result in functional differences. For example, only LIN28B contains NLS sequence [60].
Furthermore, the LET-7-independent pathways of these proteins are still largely unknown and it is plausible that they have dissimilar mRNA targets. On the other hand, at least in cases where the direct target of LIN28 is LET-7, one should expect that the consequences of LIN28A expression would be similar to those of LIN28B expression. In these cases, both of the genes have the potential to cause tumor formation and the abundance of LIN28B compared to LIN28A can be explained at the regulation levels—for some reason LIN28B misregulation (and overexpression) is more common than LIN28A misregulation. Wilms' tumor is a fitting example, as both paralogs are sufficient to form the tumor in a mouse model, but only LIN28B is expressed in primary tumors in humans [33]. An example of a specific regulator of LIN28B, but not LIN28A, is macroH2A1 histone variant. In a recent article [61], Park et al. identified the macroH2A1 histone variant as a LIN28B upstream regulator in bladder cancer. Loss of macroH2A1 expression significantly correlated with the elevated levels of LIN28B expression and subsequently inhibited the mature LET-7 microRNA expression.
From a clinical perspective, it is probably sufficient to verify in each specific tumor, which one of the two paralogs is expressed. However, if one wishes to study the molecular mechanism underlying the formation of LIN28 tumors, a critical question is whether only one of the two genes has the potential to generate the tumor.
To Conclude
The role of LIN28 as an oncogene has been extensively demonstrated and discussed elsewhere [22,24,25]. It has been shown in many types of cancers that LIN28 is expressed in the more aggressive, less differentiated cells of the tumor, and in some cases in the cancer stem cells [25]. In the current review, we emphasized the function of LIN28 in solid pediatric cancer. We showed that LIN28 (especially LIN28B) overexpression is very common in solid pediatric cancer. This observation, along with the fact that LIN28 is a heterochronic gene that is normally expressed in stem cells, suggest that in the case of pediatric cancer LIN28 functions by blocking normal development/differentiation of progenitor cells.
While we focused in this review on solid cancers, the effect of LIN28 overexpression on abnormal development and tumor formation might be expanded also to hematopoietic cancers. It has been shown that overexpression of LIN28B in hematopoietic cells of transgenic mice results in peripheral T-cell lymphoma (PTCL), and that before the onset of the PTCL, the otherwise healthy mice showed evidence of aberrant T-cell development [62]. At least in some cases, aberrant hematopoietic development preceded malignant transformation [63]; thus, it is tempting to assume that in this case also, LIN28B overexpression leads to tumor formation by preventing normal development.
The observation that LIN28 overexpression can lead to the existence of undifferentiated cancer cells through blocking of differentiation (mainly in pediatric cancers) or by dedifferentiation of adult tissue (in adult cancers), highlights the fundamental differences between pediatric and adult cancers from a genetic perspective. In the future, it would be interesting to explore to what extent this observation is true for other stem cell factors as well. In terms of clinical applications, understanding the exact molecular and pathophysiological mechanisms by which LIN28 promotes tumor formation/tumor maintenance in each specific tumor type may open new venues for therapeutic research and development.
Footnotes
Author Disclosure Statement
No competing financial interests exist.