
Abstract
Mesenchymal stromal cells (MSC) are increasingly used as an investigative therapeutic product for immune disorders and degenerative disease. Typically, MSC are isolated from human tissue, expanded in culture, and cryopreserved until usage. The safety and efficacy of MSC therapy will depend on the phenotypical and functional characteristics of MSC. The freeze-thawing procedure may change these characteristics. Furthermore, the cells encounter a microenvironment after administration that may impact their properties. It has been demonstrated that the majority of MSC localize to the lungs after intravenous infusion, making this the site to study the effects of the in vivo milieu on administered MSC. In this study, we investigated the effect of freeze-thawing and the mouse lung microenvironment on human adipose tissue-derived MSC. There were effects of freeze-thawing on the whole genome expression profile of MSC, although the effects did not exceed interdonor differences. There were no major changes in the expression of hemostatic regulators on transcriptional level, but significantly increased expression of procoagulant tissue factor on the surface of thawed adipose MSC, correlating with increased procoagulant activity of thawed cells. Exposure for 2 h to the lung microenvironment had a major effect on MSC gene expression and affected several immunological pathways. This indicates that MSC undergo functional changes shortly after infusion and this may influence the efficacy of MSC to modulate inflammatory responses. The results of this study demonstrate that MSC rapidly alter in response to the local milieu and disease-specific conditions may shape MSC after administration.
Introduction
M
There are a few factors concerning MSC functionality that could explain some of the discrepant outcomes. First, culture-expanded MSC may not be fully compatible with human blood and trigger the instant blood-mediated inflammatory reaction (IBMIR) when administered intravenously [2]. Second, one of the recurring differences between preclinical and clinical studies is the use of MSC from continuous cultures in vitro and in most preclinical studies, as opposed to the use of cryopreserved MSC in the large majority of clinical studies [3]. Third, the phenotype and functionality of MSC may change upon encounter with the in vivo microenvironment after administration.
The IBMIR reaction toward MSC may on the one hand compromise therapeutic cell survival, but on the other hand also trigger their beneficial paracrine effects in vivo [2]. Triggering of IBMIR results in release of factors that can activate MSC, but may also promote priming of anti-inflammatory effector cell types (e.g., regulatory T cells, myeloid-derived suppressor cells, and alternatively activated macrophages) in response to opsonized MSC [4,5].
There are indications for significant differences between MSC that are thawed shortly before use and MSC that come straight from the culture flask. Frozen–thawed MSC have impaired immunomodulatory properties and demonstrate increased triggering of IBMIR compared to MSC from continuous culture [6]. Thawed MSC furthermore show elevated levels of heat shock proteins and impaired responsiveness to inflammatory conditions within the first 24 h after thawing [7]. It is therefore possible that cryopreserved MSC are less effective than MSC from continuous culture for certain purposes, whereas for other applications cryopreserved MSC may be particularly suitable. It is important for the advancement of MSC therapy that the effects of cryopreservation on cell functionality are mapped in detail so that optimally effective MSC can be used for therapy.
Working with living cells implies that the cells can change their phenotypical and functional properties in response to environmental stimuli. The in vivo milieu that cells encounter upon administration may influence cellular function. However, insufficient knowledge of the homing habits of administered MSC, limited cell survival, and the complexity of the in vivo environment make it difficult to analyze the changes that MSC undergo after administration [8]. Some studies have demonstrated the isolation and reculture of administered MSC from mice [9,10], but none have been able to analyze the function of administered MSC in vivo.
It has become clear that intravenously injected MSC initially accumulate in the lungs due to size restrictions of the lung microvasculature [11]. In the lungs, MSC encounter pulmonary microvascular endothelial cells and resident macrophages and may undergo reciprocal interactions with these cells. It has been demonstrated that MSC affect lung endothelial cells by restoring endothelial permeability by the secretion of hepatocyte growth factor [12].
We have previously demonstrated that the expression levels of multiple cytokines and chemokines in the lungs are modulated after infusion of MSC [13]. The cytokines and chemokines are most likely derived from lung endothelial cells and lung-resident immune cells. Lung-derived factors and intercellular cell surface molecule interactions may have an effect on administered MSC. This would suggest that the functionality of MSC can change already shortly after administration when MSC are present in the lungs and this may alter the therapeutic effect of MSC. A better understanding of the interplay between MSC and the lung tissue-resident cells may lead to optimization of current MSC therapy protocols.
In this study, we examined phenotypical differences between cryopreserved MSC and MSC from continuous culture and analyzed the effect of the lung microvasculature milieu on MSC properties.
Materials and Methods
Isolation and culture of human MSC
MSC were isolated from abdominal subcutaneous adipose tissue of healthy individuals that became available upon kidney donation procedure after written informed consent (protocol no. MEC-2006-190 approved by the Medical Ethics Committee of the Erasmus Medical Center). After collection, the tissue was kept in minimum essential medium-α (MEM-α) (Sigma-Aldrich, St. Louis, MO) supplemented with 1% penicillin/streptomycin solution (P/S; 100 IU/mL penicillin, 100 IU/mL streptomycin; Lonza, Verviers, Belgium) at 4°C and MSC isolated within 24 h. The tissue was minced and enzymatically digested with sterile 0.5 mg/mL collagenase type IV (Life Technologies, Paisley, United Kingdom) at 37°C for 30 min under continuous shaking. The obtained cell suspension was then washed twice, resuspended in culture medium consisting of MEM-α with 1% P/S, 2 mM
Freeze–thawing procedure
For cryostorage, MSC were removed from their culture flasks by trypsinization and washed in MEM-α with 1% P/S and 15% FBS. They were then resuspended in MEM-α with 1% P/S and 15% FBS at 1 × 106 cells per mL and mixed 1:1 with MEM-α with 20% dimethyl sulfoxide (Merck, Darmstadt, Germany) and 20% FBS, aliquoted in cryovials, and placed in a freezing box (Coolcell; Biocision, San Rafael, CA) at −150°C. To thaw the cells, cryovials were placed in a 37°C water bath until nearly all the ice was melted. The cells were then washed and kept in MEM-α with 1% P/S, 2 mM
Immunophenotyping of MSC
Flow cytometric analysis was conducted on MSC labeled with monoclonal antibodies as outlined in the supporting information (Supplementary Table S1; Supplementary Data are available online at
In vitro blood clotting assay
The clotting time of human blood was recorded on a semiautomatic 10-channel ball coagulometer (MC10plus; Merlin Medical ABW Medizin und Technik GmbH, Lemgo, Germany), as reported earlier [14]. MSC directly from the culture flask or after thawing were washed twice and resuspended in a buffer containing 5% human serum albumin. Sodium citrate-anticoagulated human blood was obtained from healthy volunteers who had not received any medication for at least 10 days. The cuvette was filled with 100 μL of citrated blood diluted 1:1 in phosphate-buffered saline (PBS). Blood was then supplemented with 50 μL of buffer with or without 3,000 MSC or with 50 μL of positive control reagent. To initiate clotting, 50 μL of 40 mM Ca2+ solution was added to a final concentration of 10 mM. The final concentration of MSC was 15,000 cells/mL, corresponding to a dose of 1–2 × 106 cells/kg commonly used in clinical trials.
Administration of human MSC in mice
MSC of four healthy human donors were trypisinized, washed, put through a 40 μm cell strainer, and 1 × 106 MSC in 200 μL PBS were administered in female C57BL/6 mice (Charles River, Wilmington, MA) through tail vein injections. After 2 h, the animals were anesthetized, blood collected in EDTA tubes, and lung tissue removed and snap-frozen for RNA isolation. The animal experiments were approved by the Animal Care and Use Committee of the Erasmus Medical Center (protocol no. EMC-3004).
Human MSC gene expression analysis by RNA sequencing
MSC gene expression was analyzed immediately after removal of MSC from culture flasks, after thawing of cryopreserved MSC, and in MSC that were trapped in the lungs after intravenous administration in mice using mRNA sequencing. For the first group, RNA was isolated from MSC that were snap-frozen immediately after trypsinization (group C). For the second group, RNA was isolated from froze–thawed MSC that were snap-frozen 1 h after recovery at 37°C (group FT). For the third group, RNA was isolated from lung tissue 2 h after infusion of MSC (group I). These samples contained mouse RNA mixed with human RNA from the injected MSC. For each group, MSC of the same three donors of the same passage were used. Lung tissue of a mouse that was injected with PBS was used as a negative control (sample M).
RNA was isolated using Trizol reagent (Life Technologies). Frozen lung tissue was sectioned in 20 μm slices before RNA isolation. Quantity and quality of RNA was assessed using the RNA 6000 Nano kit on a 2100 Bio-analyzer (Agilent, Palo Alto, CA). Samples with an RNA-integrity >8.5 were used. Samples were prepped with TruSeq RNA (v2, Illumina), sequenced SR43 bp on Hiseq2500 in rapid mode, and demultiplexed with CASAVA 1.8.4. Alignment was performed with TopHat 2.0.13 (large index mode,
Fragment counts per gene were calculated from the TopHat BAM files, using a custom R (
Including the mouse chromosomes in the reference genome was not enough to prevent all cross-alignment, as for regions identical between human and mouse, reads get assigned randomly to one of them. This results in human genes having a high read count for sample M, where no alignment against human genes is expected. In samples from group I, this results in highly nonuniform coverage over the transcript for human genes with regions identical between mouse and human. These genes show high peaks in the homologous regions originating from mouse RNA. These genes show up spuriously as significantly differentially expressed. We filtered these genes out of the final list of differentially expressed genes for the comparison of group I versus group C, using a custom-built R script and the dplyr package (CRAN;
Ingenuity software (Qiagen, Venlo, Netherlands) was used for pathway analysis.
Detection of C3 activation fragment a and thrombin–antithrombin complex in murine plasma
Formation of blood activation markers thrombin–antithrombin complex (TAT) and complement component C3 activation fragment a (C3a) in murine plasma was measured with enzyme-linked immunosorbent assay (Cusabio Biotech Ltd., Wuhan, China), at 2, 8, and 24 h post MSC infusion.
Detection of cytokine/chemokine levels in lung tissue
To detect levels of cytokines and chemokines in lung tissue, frozen lung tissue was weighted and sliced in 10-μm-thick slices. The sliced tissue was centrifuged at 15,000 g and tissue fluid collected for cytokine/chemokine measurement by mouse cytokine/chemokine magnetic bead panel multiplex assay (Merck Millipore, Billerica, MA). The panel contained granulocyte colony-stimulating factor (G-CSF), granulocyte-macrophage colony stimulating factor (GM-CSF), interferon (IFN)γ, interleukin (IL)1β, IL6, IL7, IL10, CXCL1, CXCL5, CXCL10, CCL11, monocyte chemoattractant protein 1 (MCP1), MIP1α, and tumor necrosis factor (TNF)α. The samples were measured by Luminex 100/200 cytometer (Luminex, Austin, TX) using Xponent software.
Statistical analysis
Data were tested for significance using analysis of variance and Student's t-test. If the data did not fit a normal distribution, the Mann-Whitney test or the Wilcoxon matched-pairs test was used (two-tailed, 95% confidence intervals). Post hoc analysis was performed using Bonferroni test for multiple comparisons. P-values <0.05 were considered statistically significant.
Results
Gene expression profiling of MSC from continuous culture versus frozen–thawed MSC
To examine the effect of freeze–thawing on MSC, human adipose tissue-derived MSC of three healthy donors from continuous culture (C1, C2, C3) and of frozen-thawed MSC of the same donors (FT1, FT2, FT3) were used for transcriptome profiling by RNA sequencing. Gene expression clustering analysis demonstrated that interdonor gene expression differences were larger than the effects of cryopreservation, as frozen–thawed and cryopreserved MSC clustered per donor (Fig. 1A).
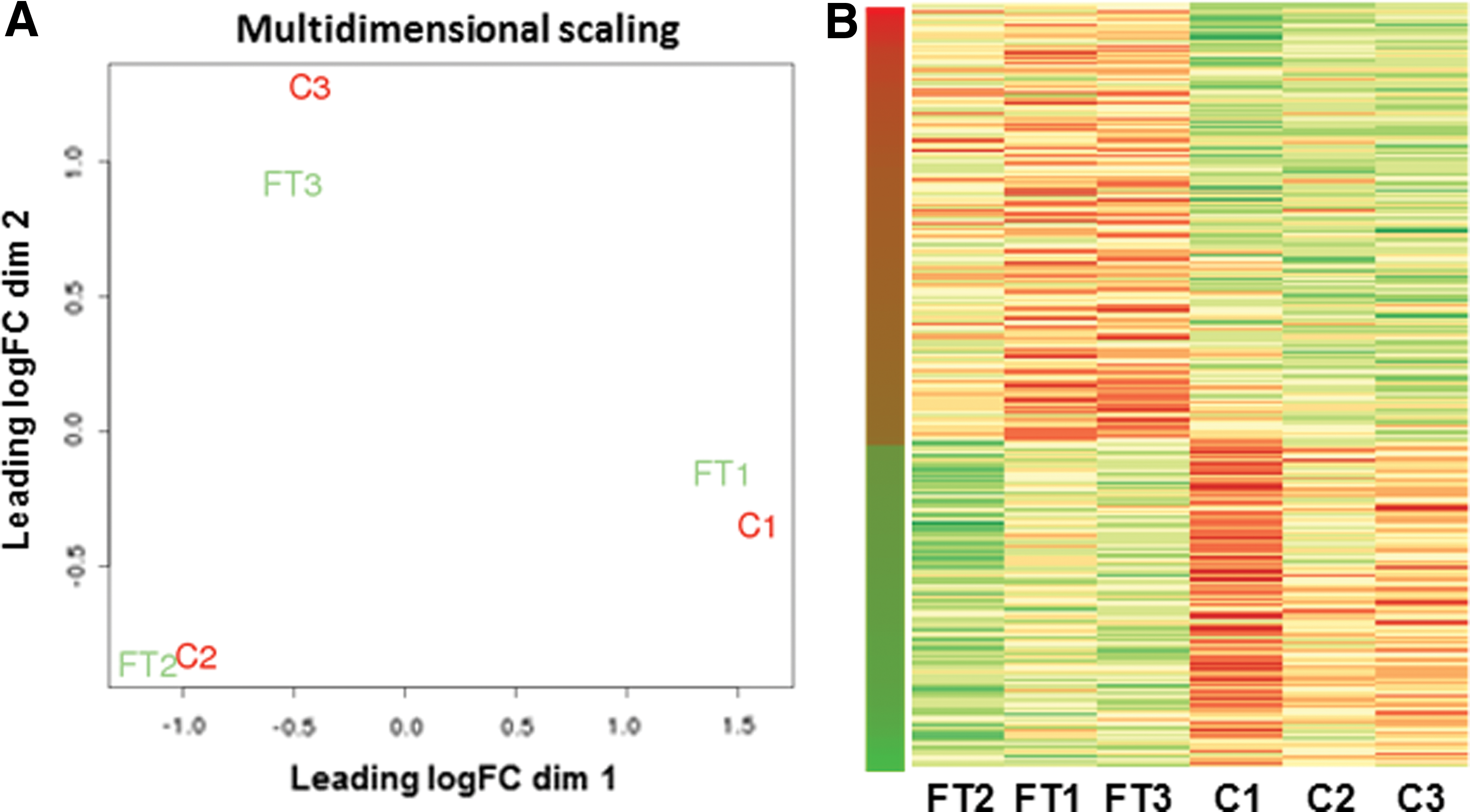
mRNA expression analysis of human MSC from continuous culture (C) and frozen–thawed MSC (FT).
Between the frozen–thawed MSC and continuous culture MSC groups, there were 294 genes that showed a significantly different expression. Of these genes, 167 were upregulated in the frozen–thawed MSC and 127 were downregulated (Fig. 1B). The magnitude of gene expression differences was limited to a maximum increase of 10.9-fold and a maximum decrease of 5.1-fold. A summary of the most significant gene expression changes can be found in Table 1. The full data set can be accessed at GEO accession number GSE76081.
Fifteen genes with the largest increases and 15 genes with the largest decreases are shown. All gene expression changes in the table are significant with P-values <0.05. Brief descriptions of gene functions are indicated underneath the gene names. Five pathways that showed the most significant up or downregulated activity patterns are listed with genes involved.
MSC, mesenchymal stem cells; ↑, upregulation; ↓, downregulation.
Pathway analysis of MSC from continuous culture versus frozen–thawed MSC
A number of individual genes involved in the inhibition of cellular proliferation and induction of growth arrest were upregulated, but pathway analysis did not reveal differential expression of cell survival or cell apoptosis pathways. Freeze–thawing of MSC had an effect on innate immunity pathways (Table 1). These pathways indicated an activation of the acute phase response in frozen-thawed MSC, macrophage inhibitory factor signaling, and activation of Toll-like receptor pathways through high-mobility group protein B1 signalling. A number of genes that are involved in actin rearrangement were upregulated in frozen–thawed cells, such as a number of Rho GTPases and actin-related protein, suggesting active cytoskeletal reorganization processes in MSC after recovering from cryopreservation.
Effects of freeze–thawing on MSC immunophenotype and IBMIR induction
The cell surface expression of a panel of MSC markers was unaffected by freeze–thawing (Fig. 2A). However, a small, but significant increase in the expression of the coagulation factor CD142 (tissue factor, TF) was found on thawed compared to fresh MSC (62% vs. 54% positive). Flow cytometry revealed that higher surface expression of TF on thawed cells went in hand with a small, but significant increase in the number of propidium iodide incorporating cells (84% vs. 91% viable, Fig. 2B). This indicates a small increase in membrane permeability postthawing, which may well explain the increase in TF expression, normally stored in submembrane intracellular granules [2].

Effect of freeze–thawing on MSC immunophenotype and triggering of innate immune cascade activation after whole blood exposure in vitro and in vivo.
To examine whether freeze–thawing would affect IBMIR triggering, the effect of MSC from continuous culture and frozen–thawed MSC on the clotting time of human blood was examined in vitro. The addition of MSC from continuous culture accelerated blood clotting time (Fig. 2C). Frozen–thawed MSC induced a further, small but significant, acceleration of clotting time compared to continuously cultured MSC.
To corroborate our in vitro findings with the in vivo situation, we analyzed murine EDTA-plasma for formation of complement and coagulation markers at 2, 8, and 24 h post-MSC infusion (Fig. 2D). Infusions of thawed cells lead to significantly higher formation of complement activation marker C3a and coagulation marker TAT at 8 h postinfusion (both P < 0.05).
Effect of MSC from continuous culture and frozen–thawed MSC on lung immune homeostasis
To investigate whether MSC from continuous culture and frozen–thawed MSC had a differential effect on the microenvironment after infusion, 1 × 106 human MSC were injected intravenously in C57BL/6 mice. As MSC accumulate in the lungs after intravenous administration, they may interact with endothelial cells of the lung microvasculature and innate immune cells such as macrophages and granulocytes. Two hours after injection of MSC, lungs were removed and mouse cytokine and chemokine expression analyzed by multiplex assay. Injection of MSC from continuous culture as well as frozen–thawed MSC demonstrated a clear tendency for increased levels of mouse G-CSF, CXCL1, CXCL10, MCP1, and IL6 and a decrease in CXCL5 in lung tissue (Fig. 3). MSC did not induce changes in the expression of IFNγ, CCL11, GM-CSF, IL1β, IL7, IL10, MIP1α, and TNFα (Supplementary Fig. S1). We have previously demonstrated that MSC induce a mild inflammatory response in the lungs, which may be associated with the immunomodulatory effect of MSC [13]. The present data demonstrate that there was no difference between the effects of MSC from continuous culture and frozen–thawed MSC, suggesting that the two cell preparations have a similar immunomodulatory effect in the lungs.

Effect of infusion of human MSC from continuous culture (C) and frozen–thawed MSC (FT) on mouse cytokine and chemokine expression in the mouse lung. 1 × 106 MSC were infused through the tail vein and 2 h later, lungs removed for analysis. N = 4 MSC donors. Bar graphs depict mean ± standard deviation. *P < 0.05. G-CSF, granulocyte colony-stimulating factor; MCP1, monocyte chemoattractant protein 1.
Gene expression profiling of MSC after in vivo administration
To examine whether MSC are stabile after in vivo administration or whether they undergo changes under influence of the in vivo environment, we carried out transcriptome analysis on administered MSC. Human MSC from continuous culture (1 × 106) were infused in the tail vein of C57BL/6 mice. The lungs, containing entrapped MSC, were removed 2 h after infusion and total RNA isolated (I1, I2, I3). RNA of all samples was sequenced and mapped against the combined human/mouse genome, thus reflecting human MSC gene expression in the mouse lung. Lung tissue of a mouse that did not receive human MSC was used as a negative control (M). In sample M, positive expression of several human genes was detected, reflecting cross-alignment of mouse RNA on the human genome. These particular genes, which represented 30% of the originally upregulated and 60% of the downregulated genes, were disregarded in samples I1-3. Genes in which <50% of exons showed positive reads in samples I1-3, also reflecting alignment of mouse RNA on the human genome, were also disregarded (20% of the originally differentially expressed genes).
The gene expression profile of the administered MSC differed considerably from both noninjected MSC groups, as can be seen in Figure 4A, where samples C1-3 and FT1-3 now appear to overlap, whereas I1-3 stand out. Negative control sample M stands out from all the other samples (Fig. 4A, B).
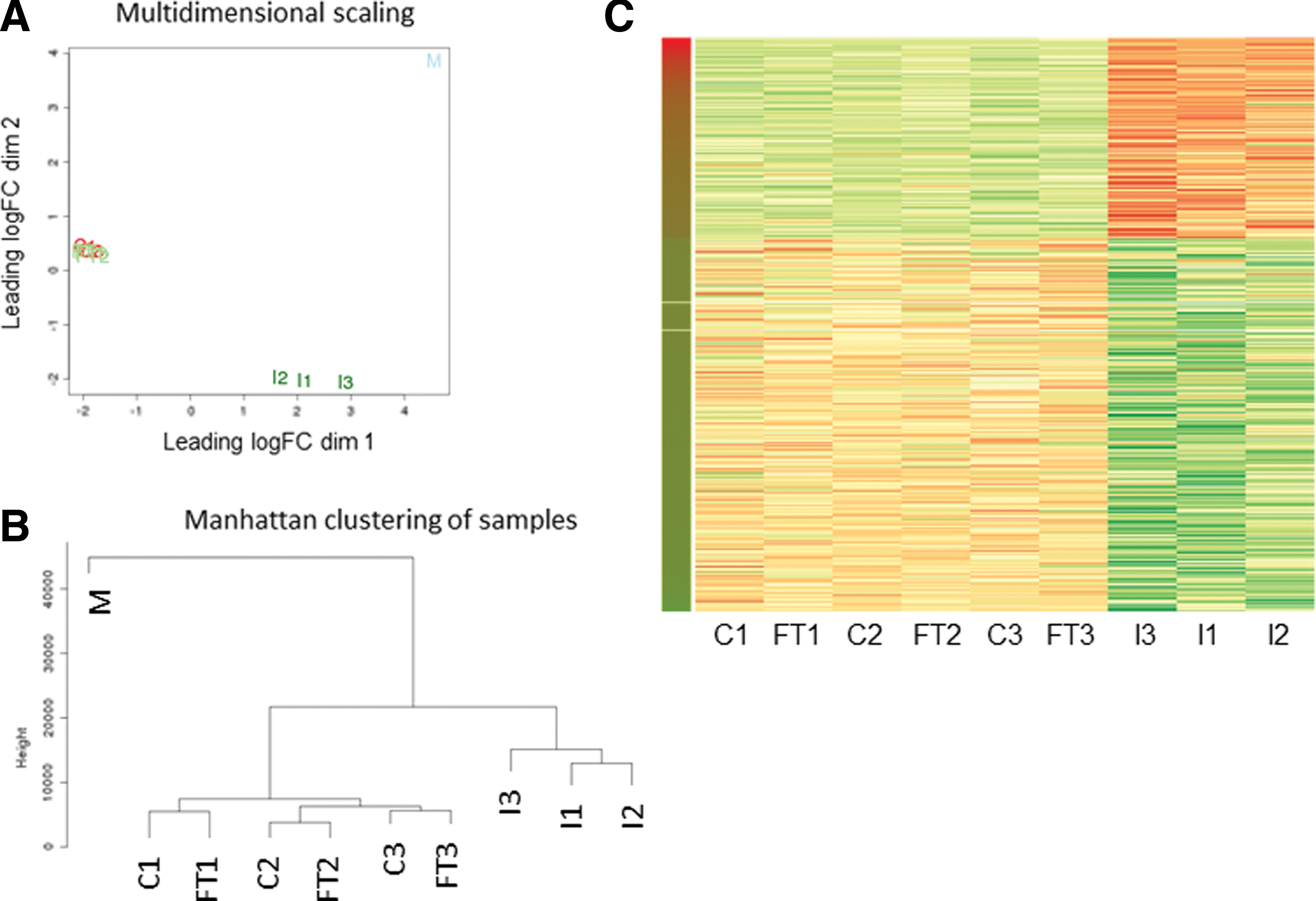
mRNA expression analysis of human MSC from continuous culture (C), frozen–thawed MSC (FT), and injected MSC (I).
Comparison of gene expression profiles between MSC from continuous culture before infusion (C1–3) and after infusion (I1–3) revealed differential expression of 2,060 genes. Of these genes, 720 were upregulated in the infused MSC and 1,340 were downregulated (Fig. 4C). The maximum increase in gene expression in the injected MSC was 1,607-fold. The decreases in gene expression were smaller, with 1,326 of the downregulated genes showing a fold change of <4. The maximum decrease was −10.3-fold. A summary of the most significant gene expression changes can be found in Table 2. The full data set can be accessed at GEO accession number GSE76081.
Fifteen genes with the largest increases and 15 genes with the largest decreases are shown. All gene expression changes in the table are significant with P-values <0.05. Brief descriptions of gene functions are indicated underneath the gene names. Five pathways that showed the most significant up or downregulated activity patterns are listed with genes involved.
↑, upregulation; ↓, downregulation.
Pathway analysis of MSC after in vivo administration
Pathway analysis 2 h after administration demonstrated that the lung microenvironment affected pathways with an immunological function in MSC. MSC showed strongly increased expression of various human leukocyte antigen class II molecules, which are upregulated in response to inflammatory cytokines. Furthermore, there was a strong upregulation of IFNγ-induced protein 10 (or CXCL10), of the common gamma chain of the receptor for IL2, IL4, IL7, IL9, IL15, and IL21, of nuclear factor of activated T cells, which modulates gene expression during immune activation, and of IFN-regulatory factor 4.
The four pathways that showed the most significantly altered patterns of activation had an immunological function. Immune signaling through the OX40 Tumor Necrosis Factor family pathway and the cytosolic pattern recognition receptor pathway showed reduced activity, whereas the TNF receptor 2 pathway and the phospho-kinase C signaling pathway showed enhanced activity (Table 2), indicating a modulation of the immunomodulatory activity of MSC upon administration.
Even though it is known that MSC have a short survival after intravenous infusion, cell death pathways were not activated. The apoptosis signaling pathway showed no pattern of activation, while the HIV-induced apoptosis pathway showed reduced activity.
Interestingly, multiple genes involved in tyrosine metabolism, such as tyrosinase and tyrosinase-related proteins 1 and 2, were among the most highly upregulated genes.
Production of soluble factors by injected MSC
As the therapeutic effect of MSC is thought to be partly dependent on secreted factors, the expression of genes encoding proteins with an extracellular function was analyzed. Eighteen soluble factors showed upregulated expression of at least 10-fold in injected MSC (Table 3). These factors included factors with an immune function, such as CXCL10, IL11, and IL33, and growth factors and factors that stimulate regeneration by progenitor cells, such as wingless-type MMTV integration site family member (WNTs) and bone morphogenetic protein (BMP)2.
All gene expression changes listed are highly significant (P < 10−5, or smaller).
Discussion
The efficiency of MSC therapy will depend for a great part on the phenotype of MSC preparations. In contrast to conventional molecular drugs that, apart from being metabolized by the recipient, do not change, cells can undergo dramatic alterations in response to changes in their microenvironment. Freeze–thawing has been indicated to affect the in vitro immunomodulatory properties of MSC [6,7]. However, MSC functionality is completely recovered after a 24-h culture period. In this study, we found limited gene expression changes in MSC 1 h after thawing. There were changes in genes involved in innate immunity pathways and cytoskeletal rearrangement. The cryopreserved cells were kept in suspension for 1 h after thawing and it is possible that these conditions induced the cells to upregulate cytoskeletal protein expression.
MSC from continuous culture and frozen–thawed MSC induced comparable immunological responses in the mouse lung, the major site of MSC embolization upon intravenous infusion. Indeed, a study by Cruz et al. demonstrated only limited differences between the effects of cryopreserved and continuously cultured MSC in ameliorating airway inflammation, supporting the concept that both MSC preparations are equally effective in vivo [15]. It is likely that the type of model used is determinative for detecting differences between frozen–thawed and continuously cultured MSC or not, as freeze–thawing may affect particular properties of MSC that are employed in particular models, but not in others.
It has previously been demonstrated that islets of Langerhans [16], but also culture expanded MSC, can induce IBMIR [2]. The induction of IBMIR was weakly augmented with frozen-thawed MSC [6,17]. A strong induction of IBMIR would reduce the survival time of MSC after administration and could potentially lead to adverse effects, providing a possible explanation for the limited engraftment of therapeutic MSC [18] and adverse transfusion reactions at higher doses [14]. We found a strong induction of IBMIR for the adipose MSC used in this study, which is in agreement with other reports attributing a strong procoagulant activity to adipose tissue-derived MSC [19 –21]. We measured a weak, but significantly augmented triggering of IBMIR with frozen–thawed cells. The physiological significance of this difference between continuously cultured and frozen–thawed MSC is unclear. However, most importantly, both cell types elicited strong responses. Thus, in vivo persistence will be rather limited with both continuously- and frozen–thawed adipose-derived MSC alike, not necessarily compromising their bioactivity.
In contrast to the minimal effects of freeze–thawing, in vivo administration had a major effect on the transcriptional phenotype of MSC. The observed changes induced in MSC could derive from a number of factors, among them the effects of sheer-stress induced by transport through the bloodstream, factors induced by triggering of IBMIR or the effects of blood cells, and proinflammatory mediators on MSC. MSC accumulate in the lungs upon the first passage [11] and thus within minutes of administration, the majority of MSC are exposed to the microenvironment of the lung microvasculature, where lung endothelial cells and macrophages may undergo interactions with MSC. The gene expression changes in MSC after administration indicate a response to inflammatory signals, eliciting immunological cross talk between entrapped MSC and tissue-resident immune cells. This suggests that cells in the lung microvasculature become activated upon encounter of MSC and respond with inflammatory signals. We have demonstrated previously that lung tissue shows an inflammatory gene expression profile within hours after infusion of MSC [13] and in this study, we demonstrated that levels of the mouse inflammatory factors IL6, CXCL1, CXCL10, and MCP1 were increased in lung tissue upon administration of human MSC.
There is a possibility that the gene expression changes observed in human MSC after infusion in mice are caused by incompatibility of human cells with mouse microenvironment. However, in our earlier study, we found upregulation of immune parameters in mice infused with syngeneic mouse MSC [13]. Interestingly, in this study, we found that human MSC are well capable of responding to mouse cytokines and chemokines. Therefore, we assume that we are not merely looking at a xenoresponse. The nature of the analysis used does, unfortunately, not allow administration of syngeneic cells to test this assumption.
The main question of this study is how the phenotypical changes in MSC affect their function. MSC are known to enhance their immunomodulatory function in response to inflammation [22,23], and we found evidence for increased expression of IL11 and IL33, cytokines of the IL6 and IL1 families, respectively, and for the activation of immune signaling pathways (Fig. 5). There were also nonimmunological pathways that were upregulated in MSC in response to the in vivo environment. There was for instance a dramatic increase in apolipoprotein expression, which is a family of soluble proteins involved in lipid transport. Apolipoproteins may, however, also have anti-inflammatory functions [24]. Furthermore, multiple genes involved in tyrosine metabolism, including tyrosinase, tyrosinase-related protein 1, and dopachrome tautomerase, were strongly upregulated. The significance of these gene expression changes is unknown. MSC showed large increases in WNT7a and WNT9a gene expression. WNT signaling pathways have been identified to play important roles in the control of MSC proliferation and differentiation [25]. Through WNT secretion, MSC may thus target proliferation and differentiation of resident progenitor cells. These gene expression changes indicate that the microenvironment of the lung affects immunological and metabolic activity of MSC. As MSC have a short survival time after infusion, intrinsic changes in, for instance, MSC proliferation rate or differentiation potential are not relevant for the therapeutic effect of MSC. However, changes in cytokine and growth factor secretion by MSC may affect the function of resident cell types, even after MSC have disappeared. In this study, we have not examined the effect of the gene expression changes in MSC on tissues and have not determined whether concentrations of secreted factors reach effective levels. However, we can conclude from the gene expression changes that administered MSC respond rapidly to their new microenvironment and change their immunological and metabolic function.
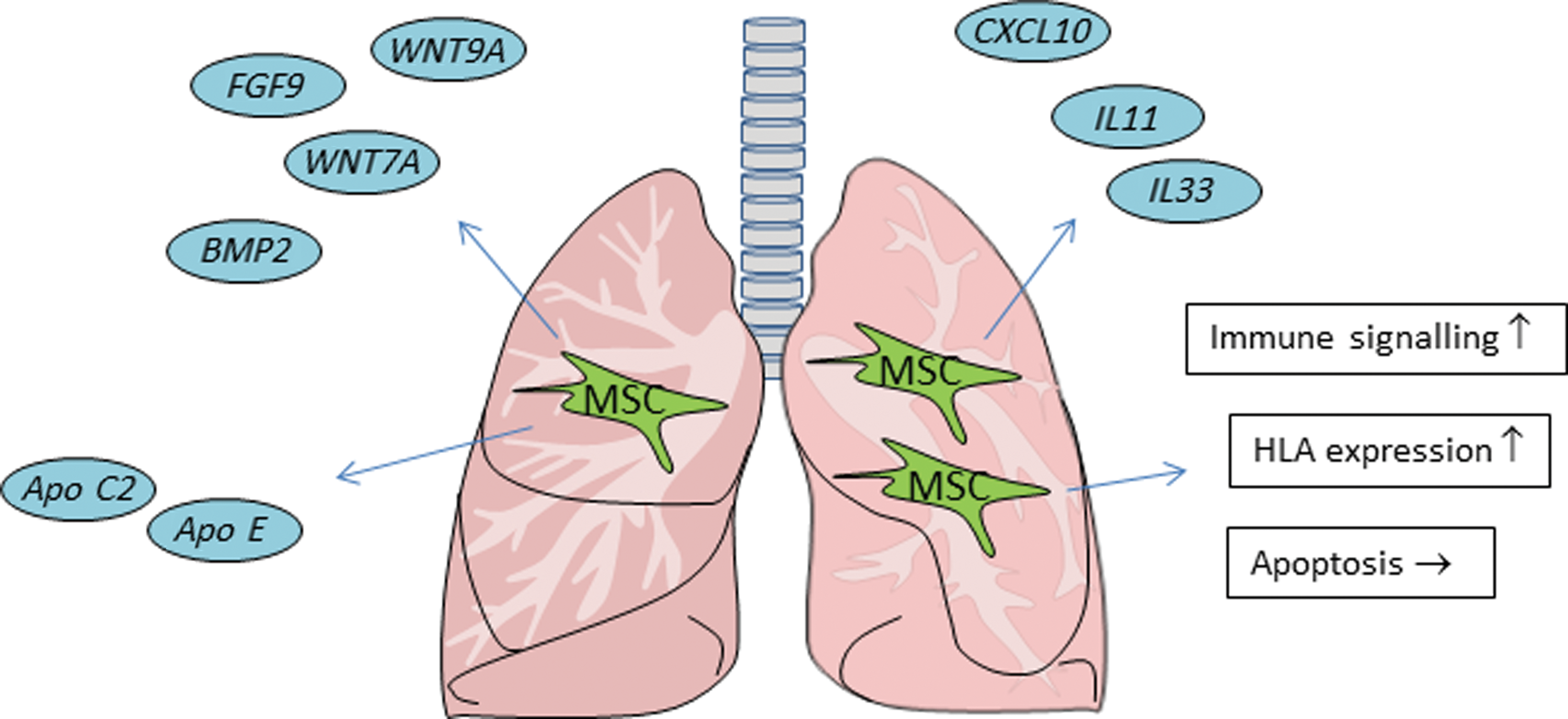
Schematic overview of the changes in MSC 2 h after intravenous administration. Infused MSC end up in the lungs and at 2 h, increase the expression of a range of immunomodulatory and growth factors and change the activity of immune and metabolic pathways. Apo C2, apolipoprotein C2; Apo E, apolipoprotein E; BMP2, bone morphogenic protein 2; FGF9, fibroblast growth factor 9; Wnt7A, wingless-type MMTV integration site family member 7A; Wnt9A, wingless-type MMTV integration site family, member 9A. Color images available online at
Earlier studies have shown that a large majority of intravenously infused MSC disappear within 24 h after administration [9]. We therefore expected to see an upregulation of apoptosis pathways in MSC upon intravenous administration. This was not the case, which would suggest that the disappearance of MSC is independent of the induction of apoptosis. Our data can however not completely prove this. First, apoptosis may be induced independently of gene expression changes. Second, the filter that we used to avoid cross-reactivity of RNA of the mouse lung tissue with the human genome may have filtered out apoptosis genes that are conserved between mouse and human [26]. Therefore, we cannot rule out the possibility that infused MSC undergo apoptosis in the lungs.
Over the last two decades, it has been shown that MSC therapy is safe [27], and the current challenge is to develop efficient therapy. Effort is put in identifying MSC subsets that possess superior immunomodulatory properties and in the development of culture protocols that generate MSC with optimized function. Efficacy testing should allow discrimination between more- and less-potent MSC batches. The findings of this study indicate that MSC show high responsiveness and plasticity upon systemic infusion. Properties that are present in vitro are not necessarily maintained after administration, and vice versa. This is a fact to be taken into account in the development of efficient MSC therapy.
In summary, this study demonstrates that freeze–thawing procedures have little impact on MSC gene expression, but tend to sensitize the cells for stronger recognition by the IBMIR. However, MSC from continuous culture and frozen–thawed MSC had a similar impact on immunological parameters in the lungs. Upon intravenous injection, MSC underwent major gene expression changes, reflecting a response to inflammatory activation. This study describes for the first time that MSC change phenotype and potentially function upon systemic administration, which is important for understanding MSC therapy and improving its efficiency.
Footnotes
Acknowledgment
Guido Moll's contributions were made possible by DFG funding through the Berlin-Brandenburg School for Regenerative Therapies (BSRT, GSC203).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.