
Abstract
Somatic cells may be reprogrammed into pluripotent cells by the ectopic expression of defined transcription factors. However, some of the hurdles that affect the generation of induced pluripotent stem cells include extremely low efficiency and slow reprogramming. In the present study, we examined the effects of small molecules on cellular reprogramming and found that 8-Bromoadenosine 3′,5′-cyclic monophosphate (8-Br-cAMP), an analog of cyclic adenosine monophosphate (cAMP), improves the reprogramming efficiency of reprogrammable mouse fibroblasts induced with dox in serum replacement (SR) medium. Interestingly, treatment with 8-Br-cAMP in mouse embryonic stem cell culture conditions does not affect reprogramming into the pluripotent state; however, reprogramming efficiency is significantly enhanced by inhibition of protein kinase A (PKA) in SR medium. Therefore, our results suggest that PKA signaling is unnecessary and may in fact act as a barrier to reprogramming into pluripotent stem cells.
Introduction
S
Recent reports suggest that the cyclic adenosine monophosphate (cAMP) signaling pathway supports the maintenance of ESCs and cellular reprogramming [13 –15]. Protein kinase A (PKA) and exchange protein activated by cAMP (EPAC) are the major targets of cAMP [16,17]. EPAC acts as a guanine nucleotide exchange factor, but, unlike PKA, does not phosphorylate target proteins. Therefore, an increase in intracellular cAMP levels mainly leads to the activation of PKA, EPAC, and cyclic nucleotide-gated ion channels. Subsequently, activation of PKA and/or EPAC signaling plays a critical role in metabolic regulation, gene expression, proliferation, differentiation, migration, and adhesion [18 –20]. Increasing cAMP levels by treatment with 8-Br-cAMP (cAMP analog) and forskolin (activator of adenylyl cyclase) has been shown to significantly enhance self-renewal in mouse ESCs (mESCs) and block differentiation on the removal of leukemia inhibitory factor (LIF) or treatment with retinoic acid [13]. Interestingly, 8-Br-cAMP was found to enhance the reprogramming efficiency of human fibroblasts by twofold [14]. On the other hand, 6-phenyl cAMP, which specifically activates EPAC, but not PKA, was shown to increase glucose uptake and proliferation in mESCs [15].
In this study, we attempt to elucidate the roles of cAMP signaling in the reprogramming process by culturing cells in different culture media, namely serum-containing or serum replacement (SR) media. We found that an increase in intracellular cAMP levels enhances the reprogramming efficiency of mouse fibroblasts in SR-based medium; however, this effect is less pronounced in serum-containing medium. We further investigated the importance of PKA and EPAC, both of which are targets of cAMP, for the efficient reprogramming of somatic cells.
Materials and Methods
Cell culture
We used fibroblasts prepared from reprogrammable OG2 (rOG2) mice to generate iPSCs that carry the heterozygous Oct4-GFP (ΔPE) transgene and the dox-inducible polycistronic cassette encoding the four reprogramming factors (Oct4, Sox2, Klf4, and c-Myc) and are homozygous for the transcriptional activator (M2rtTA) [21]. Animals were approved by the Institutional Animal Care and Use Committee of Konkuk University (Republic of Korea). The rOG2 fibroblasts were cultured in fibroblast medium: Dulbecco's modified Eagle's medium (DMEM; Gibco BRL) containing 10% fetal bovine serum (FBS; HyClone) and 1× penicillin/streptomycin/glutamine (Invitrogen). iPSCs were maintained on mouse embryonic fibroblast (MEF) feeder cells inactivated with 0.01 mg/mL mitomycin C in standard mESC culture medium: DMEM supplemented with 15% FBS, 1× penicillin/streptomycin/glutamine, 0.1 mM nonessential amino acids, 1 mM β-mercaptoethanol, and 1,000 U/mL LIF (ESGRO; Chemicon).
Generation of iPSCs
Fibroblasts were obtained from rOG2 mice and cultured for 2 days in fibroblast medium containing dox. From day 2, the transduced fibroblasts were cultured in mESC culture medium or SR-based medium supplemented with DMEM (Gibco BRL) containing 20% SR (Gibco BRL), 1× penicillin/streptomycin/glutamine, 0.1 mM nonessential amino acids, 1 mM β-mercaptoethanol, 1,000 U/mL LIF (ESGRO; Chemicon), with or without 8-Br-cAMP (B5386; Sigma), forskolin (F6886; Sigma), 8-pCPT-2′-O-Me-cAMP (C8988; Sigma), and H89 (371963; Millipore). Media and small molecules were changed daily. On day 20, Oct4-GFP-positive colonies were picked for expansion and maintained in mESC culture medium. For the quantification of reprogramming efficiency, we counted Oct4-GFP-positive colonies grown in serum-based or SR-based medium.
RNA isolation and real-time quantitative reverse transcription–polymerase chain reaction
Total RNA was isolated using the RNeasy Mini Kit (Qiagen) and digested with DNase I (RNase-free DNase; Qiagen) according to the manufacturer's instructions. Total RNA (1 μg) was reverse transcribed using SuperScriptIII Reverse Transcriptase (Invitrogen) and Oligo(dt)20-primer (Invitrogen) according to the manufacturer's instructions. Polymerase chain reaction (PCR) was performed using Taq polymerase (Takara) and appropriate primers. PCR primer pairs were as follows: Oct4 (endo) sense, 5′-GATGCTGTGAGCCAAGGCAAG-3′; Oct4 (endo) antisense, 5′-GGCTCCTGATCAACAGCATCAC-3′; β-actin sense, 5′-CGCCATGATGACGATATCG-3′; β-actin antisense, 5′-CGAAGCCGGCTTTGCACATG-3′.
Flow cytometry
Cells were pelleted by centrifugation at 1,500 rpm for 5 min and resuspended in phosphate-buffered saline (PBS). Flow cytometry analysis for Oct4-GFP fluorescence was performed using FACSCalibur (Becton Dickinson). A minimum of 30,000 events were collected for each analysis.
Bisulfite DNA sequencing analysis
Genomic DNA was treated with sodium bisulfite to convert all unmethylated cytosine residues into uracil residues using the EpiTect Bisulfite Kit (Qiagen) according to the manufacturer's protocol. Briefly, purified genomic DNA (0.5–1 μg) was denatured at 99°C and then incubated at 60°C. Modified DNA (ie, after desulfonation, neutralization, and desalting) was diluted with 20 μL of distilled water. Subsequently, bisulfite PCR (BS-PCR) amplification was carried out using 1–2 μL aliquots of modified DNA for each PCR reaction. PCR amplification of the promoter regions of Oct4 and Nanog was carried out as previously described [22,23]. PCR products were subcloned into pGEM-T Easy vector (Promega). The reconstructed plasmids were purified, and individual clones were sequenced (Solgent Corporation). Clones with ≥90% cytosine conversion were accepted and all possible clonalities were excluded based on criteria from BiQ Analyzer software (Max Plank Society). Primer pairs were as followed: Oct4-first sense, 5′-TTTGTTTTTTTATTTATTTAGGGGG-3′; Oct4-first antisense, 5′-ATCCCCAATACCTCTAAACCTAATC-3′; Oct4-second sense, 5′-GGGTTAGAGGTTAAGGTTAGAGGG-3′; Oct4-second antisense, 5′-CCCCCACCTAATAAAAATAAAAAAA-3′; Nanog-first sense, 5′-TTTGTAGGTGGGATTAATTGTGAA-3′; Nanog-first antisense, 5′-AAAAAATTTTAAACAACAACCAAAAA-3′; Nanog-second sense, 5′-TTTGTAGGTTGGGATTAATTGTGAA-3′; Nanog-second antisense, 5′-AAAAAAACAAAACACCAACCAAAT-3′.
Immunocytochemistry
Cells were fixed with 4% paraformaldehyde (PFA) for 20 min at room temperature and washed with PBS. Cells were treated with PBS containing 10% normal goat serum and 0.03% Triton X-100 for 45 min at room temperature. Primary antibodies used were anti-Nanog (1:250 dilution; Chemicon), anti-Sox2 (1:1,000 dilution; Chemicon), anti-β III tubulin (Tuj1, 1:1,000 dilution; Chemicon), anti-Brachyury (1:1,000 dilution; Chemicon), and anti-HNF3β (1:200 dilution; R&D). For detection of primary antibodies, fluorescence-labeled (Alexa Fluor 568; Molecular Probes) secondary antibody was used according to the manufacturer's specifications.
In vitro differentiation
cAMP-iPSCs were transferred into a suspension culture dish after trypsinization and cultured for 3 days in mESC culture media without LIF. The embryoid bodies were plated on 0.1% gelatin-coated glass chamber slides and cultured for 12 days. Differentiated cells were stained for three germ layer markers: Tuj1 for ectoderm, Brachyury for mesoderm, and HNF3β for endoderm.
Teratoma formation
The properties associated with teratoma formation were evaluated in trypsin-dissociated iPSCs. We subcutaneously transplanted 1 × 106 cells suspended in DMEM into the testes of 5-week-old nude mice. After 5 weeks, the tumors were dissected and fixed in 4% PFA, processed using graded ethanol, and embedded in paraffin after sectioning with hematoxylin and eosin staining.
Chimera formation analysis
cAMP-iPSCs were aggregated with denuded postcompacted eight-cell-stage embryos to obtain an aggregate chimera. Eight-cell embryos flushed from 2.5 days postcoitum (dpc) B6D2F1 female mice were cultured in microdrops of embryo culture medium under mineral oil. After cells were trypsinized for 10 s, clumps of iPSCs (4–10 cells) were selected and transferred into microdrops containing zona-free eight-cell embryos. Morula-stage embryos aggregated with cAMP-iPSCs were cultured overnight at 37°C, with 55% CO2. The aggregated blastocysts were transferred into the uterine horns of 2.5 dpc pseudopregnant recipients.
Statistical analysis
All experiments were performed in triplicate and data represented as mean ± standard deviation. Significance of differences was assessed by an unpaired t-test at P value <0.05.
Results
Generation of iPSCs from rOG2 fibroblasts
Mouse iPSCs can be generated from rOG2 MEFs in about 10 days following doxycycline (dox) induction, with a reprogramming efficiency of about 0.01%–0.5% in serum-containing mESC medium [1,24,25]. An SR-based medium (iSF medium) containing LIF was recently utilized for the generation of mouse iPSCs with a high reprogramming rate [26]. We therefore tested the effect of serum-free medium on reprogramming efficiency in rOG2 MEFs. The dox-inducible reprogramming system using rOG2 MEFs is very useful for the estimation of reprogramming efficiency without the occurrence of bias during transduction or transfection of episomal vectors [21]. Fibroblast culture medium was replaced by mESC culture medium or iSF medium at day 2 after treatment with dox. Oct4-GFP-positive colonies were detected at day 10 in serum-containing medium only at the high dose of dox (20 μg/mL) (Fig. 1A, C). However, in iSF medium, the Oct4-GFP-positive cells appeared as early as day 5 (in 5 μg/mL of dox) (Fig. 1B, D). Interestingly, high-dose dox treatment exerted negative effects on reprogramming, whereas low-dose dox treatment was found to be effective for inducing reprogramming of rOG2 MEFs (Fig. 1B). These data indicate that SR-based medium possesses an advantage over the serum-containing medium in terms of shorter lag time for reprogramming and Oct4-GFP activation in rOG2 MEFs.
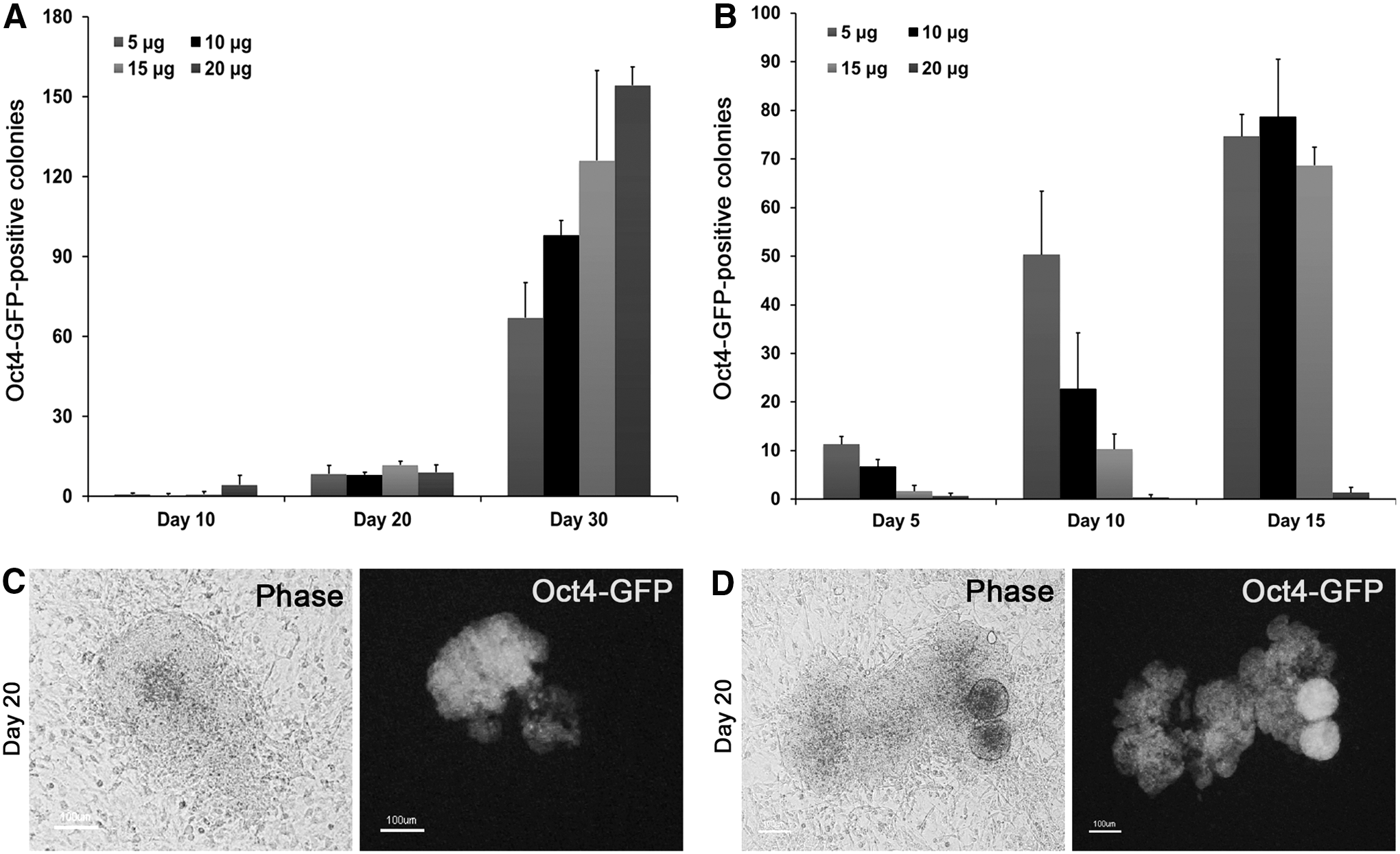
Generation of induced pluripotent stem cells (iPSCs) from reprogrammable OG2 (rOG2) fibroblasts.
Enhancement of reprogramming efficiency by activation of cAMP signaling
Recent reports suggest that reprogramming efficiency of human [14] and mouse fibroblasts [27] may be enhanced by 8-Br-cAMP treatment. In this study, we further investigated the roles of cAMP in reprogramming by treatment with the cAMP analog, 8-Br-cAMP, under two different culture conditions: serum-containing or serum-free medium or SR media. A time schedule of iPSC generation is shown in Fig. 2A. At day 2 and each subsequent day after dox induction, the medium was replaced by SR medium containing 8-Br-cAMP. Oct4-GFP+ cells were counted using FACS analysis at day 10 postinduction. FACS analysis showed that about 1.7% of cells were successfully reprogrammed into Oct4-GFP+ cells by treatment with 0.1 mM and 0.5 mM of 8-Br-cAMP at an approximately twofold higher rate than in the nontreated control group, whereas the number of GFP+ cells was <0.2% in 0.1 mM of 8-Br-cAMP and serum-containing medium (Fig. 2B).
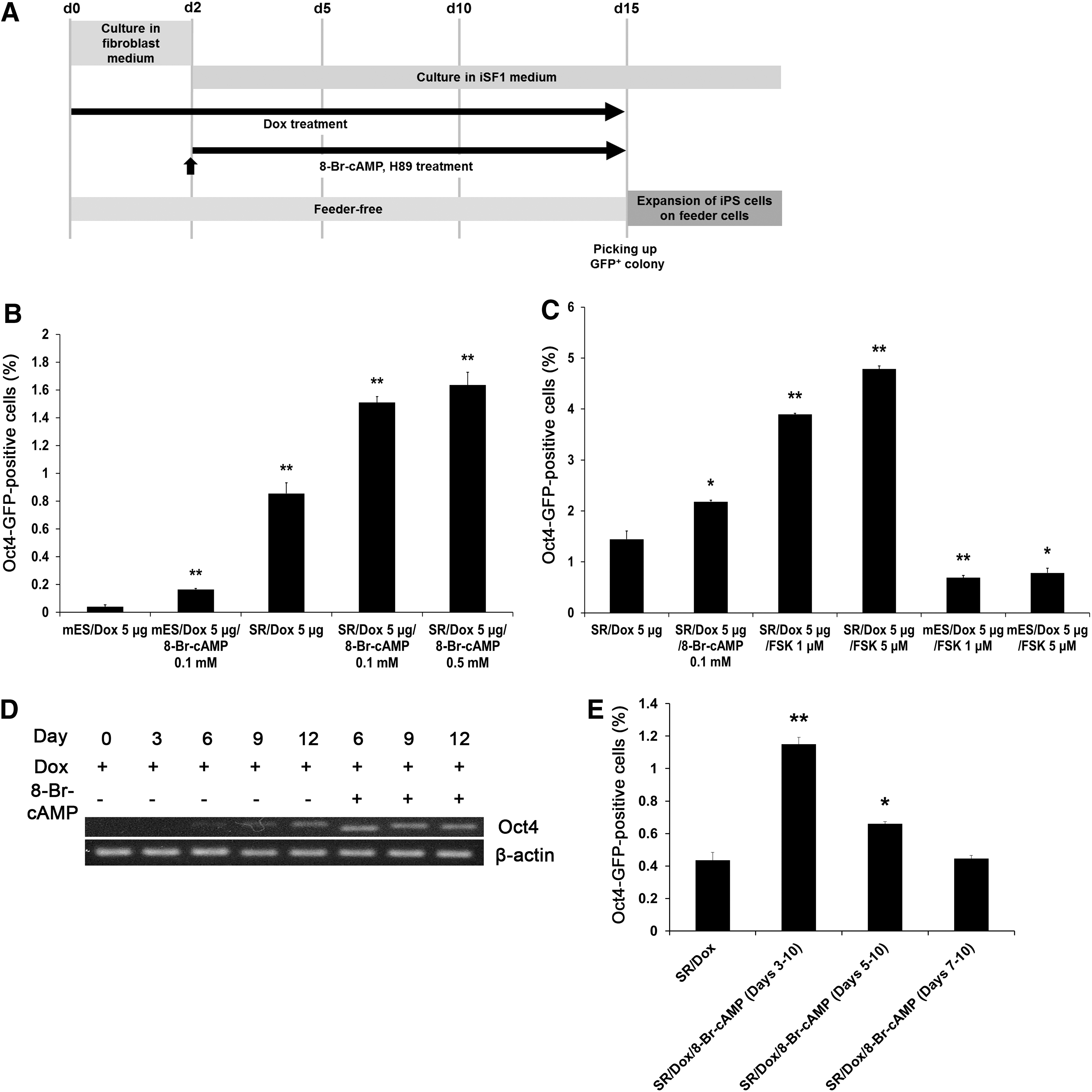
Cyclic adenosine monophosphate (cAMP) signaling enhances the reprogramming efficiency of rOG2 fibroblasts.
We next examined the effect of forskolin, an activator of adenylyl cyclase, on reprogramming efficiency. Treatment with 5 μM forskolin enhanced the number of Oct4-GFP-positive cells by approximately threefold (Fig. 2C). Notably, forskolin treatment yielded a higher number (approximately twofold) of Oct4-GFP+ cells than 8-Br-cAMP treatment (Fig. 2C). Next, we investigated the expression of endogenous Oct4. As shown in Fig. 2D, endogenous Oct4 was highly expressed on day 6 in the 8-Br-cAMP-treated group, whereas Oct4 expression was detected on day 9 in the nontreated control group (Fig. 2D). To further investigate whether cAMP signaling is required during the early stages of reprogramming, groups were treated with 8-Br-cAMP at various time points: days 3, 5, and 7 after dox induction (Fig. 2E). The later the time point at which cells were treated with 8-Br-cAMP, the lower the number of Oct4-GFP+ cells observed, indicating that cAMP signaling is crucial in the early stages of reprogramming (Fig. 2E).
Characterization of cAMP-iPSCs
Oct4-GFP+ colonies were picked from an 8-Br-cAMP-treated dish at day 10 after dox induction and expanded in serum-containing mESC culture medium. These iPSCs were designated as cAMP-iPSCs. To determine whether cAMP-iPSCs were completely reprogrammed into the pluripotent state, we examined the expression and DNA methylation status of pluripotency markers, as well as differentiation potential, in vitro and in vivo. Oct4-GFP-positive cAMP-iPSCs were stained positive for Nanog and Sox2 (Fig. 3A). Bisulfite genomic sequencing analysis revealed that Oct4 and Nanog promoter regions of cAMP-iPSCs were completely demethylated (Fig. 3B). cAMP-iPSCs differentiated into the ectoderm (Tuj1), mesoderm (Brachyury), and endoderm (HNF3β) in vitro (Fig. 3C). To examine the ability to form teratomas in vivo, we transplanted the cAMP-iPSCs into the testis capsules of immunocompromised mice. Five weeks after transplantation, the cAMP-iPSCs formed typical teratomas containing derivatives of all three germ layers (Fig. 3D). To confirm the developmental potential of cAMP-iPSCs, we performed chimera formation analysis by aggregation of iPSCs with morula embryos, followed by transfer into the uteri of pseudopregnant mice. At 13.5 dpc, somatic contribution of the cAMP-iPSCs was confirmed by genotyping for the GFP transgene in separate tissues, and germline contribution was confirmed by detection of Oct4-GFP-positive cells in the gonad (Fig. 3E). These results indicate that 8-Br-cAMP enhances reprogramming efficiency under serum-free conditions and that the properties of the resulting cAMP-iPSCs are similar to those of mESCs.
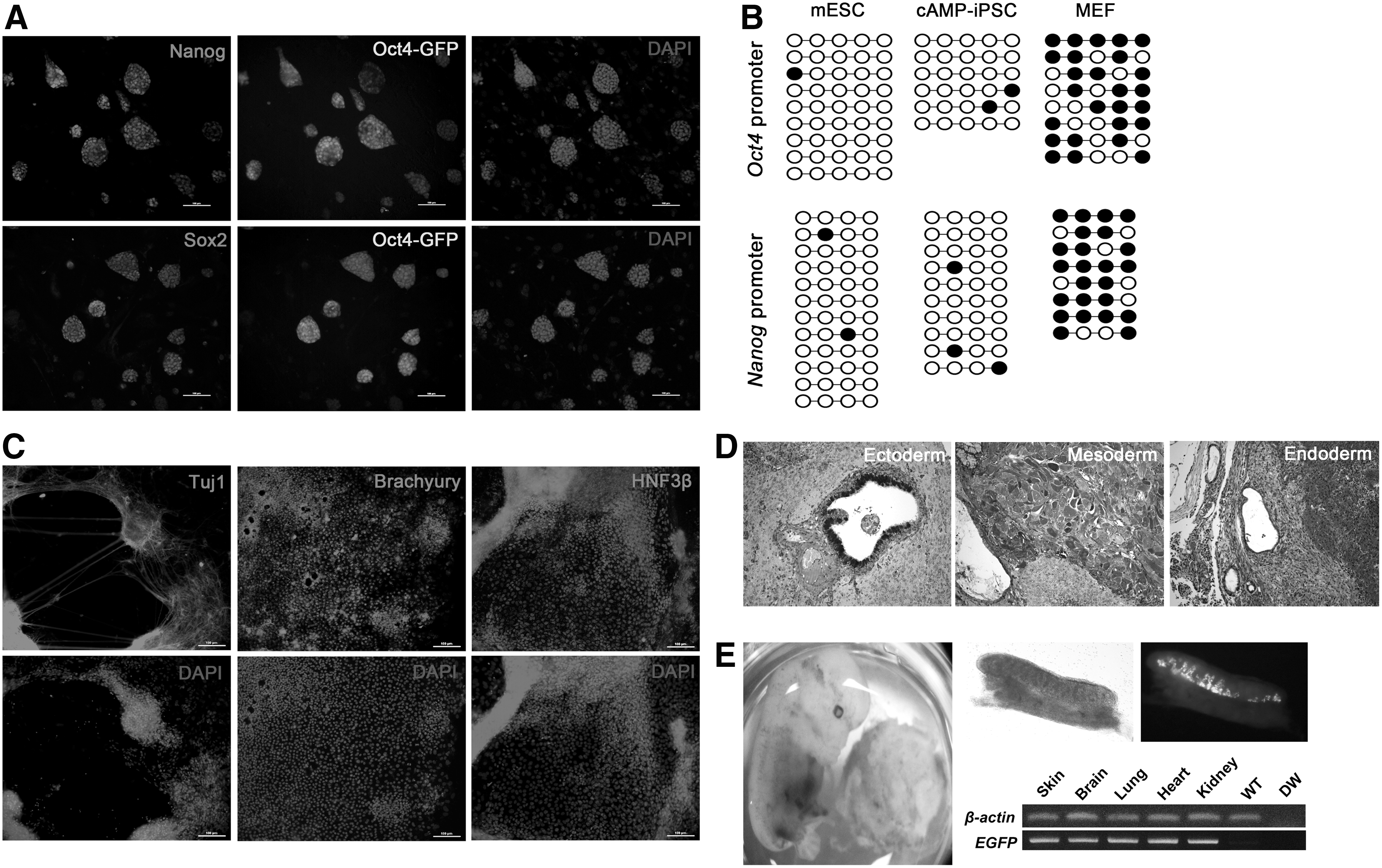
Characterization of cAMP-iPSCs.
Enhancement of reprogramming efficiency through PKA inhibition
Although the activation of adenylyl cyclase and cAMP signaling enhanced the efficiency of reprogramming, the downstream mechanisms of these effects are unclear. It is well known that activated cAMP induces the activation of PKA, which plays a role in numerous physiological processes such as gene expression, cell growth, cell division, differentiation, and metabolism. To determine whether the PKA pathway, as a cAMP target signal, plays a major role in enhancing reprogramming, we inhibited the PKA pathway using the PKA-specific inhibitor, H89. Addition of H89 (5 μM) to the reprogramming mixture in SR medium (dox and 8-Br-cAMP) considerably increased the number of Oct4-GFP-positive cells than in the nontreated control (Fig. 4A). Enhanced reprogramming due to treatment with the PKA inhibitor, H89, was also observed without 8-Br-cAMP treatment (Fig. 4A). These results suggest that PKA signaling does not play a positive role in reprogramming, but instead impedes the process.
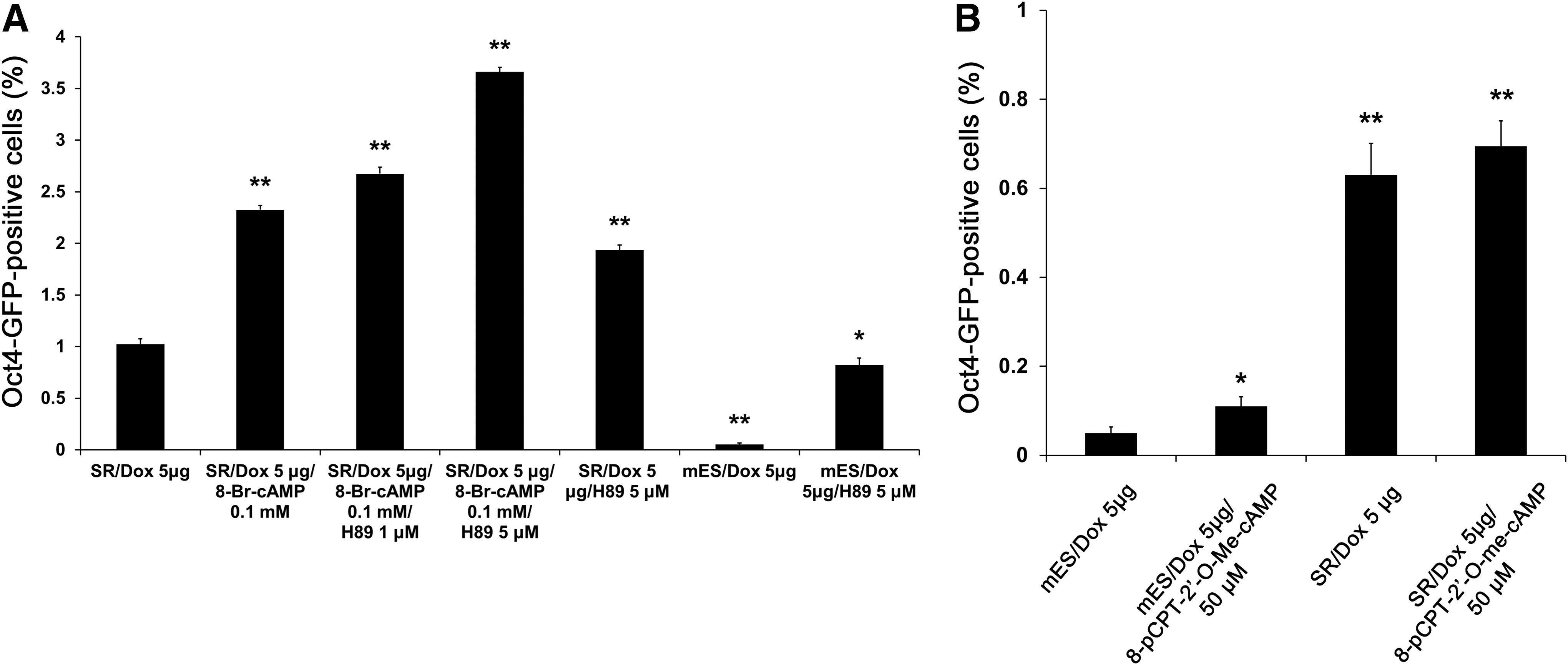
Inhibition of the protein kinase A (PKA) pathway can enhance the reprogramming efficiency.
We next investigated whether the other cAMP effector, EPAC, acts as positive effector in reprogramming. Addition of the EPAC-specific cAMP analog 8-pCPT-2′-O-Me-cAMP to the reprogramming mixture increased the number of Oct4-GFP-positive cells in mESC culture medium; however, a smaller increase in the number of Oct4-GFP-positive cells in the SR medium was observed (Fig. 4B). Taken together, these results highlight the importance of the activation of the cAMP pathway and the inhibition of PKA in enhancing reprogramming into pluripotent cells.
Discussion
Since the discovery of reprogramming using selected factors, numerous studies have focused on the development of methods and the identification of small-molecule combinations for iPSC establishment. Recent studies of the effects of the cAMP signaling pathway during reprogramming report that treatment with 8-Br-cAMP, the c-AMP analog, enhances the reprogramming efficiency of human fibroblasts by approximately twofold [14]. Additionally, it has been shown that treatment with forskolin, the activator of adenylate cyclase, transforms human ESCs to naïve pluripotency, similar to the mESC state [28,29]. Fritz et al. showed that the EPAC/Rap1 signal pathway can replace Oct4 during reprogramming under mESC culture condition [27]. However, we observed that the reprogramming efficiency following the induction of the cAMP pathway was affected by culture conditions. The reprogramming process and efficiency were significantly enhanced by the activation of the cAMP pathway in SR medium compared with serum-based medium (Fig. 2B, C). SR medium contains high amounts of vitamin C, which is suggested to improve reprogramming efficiency when added to induction media [6,30,31]. However, PKA, a major downstream target of cAMP signaling, acted as an impediment to the reprogramming process. On the other hand, treatment with the EPAC-specific cAMP analog increased reprogramming efficiency. These results suggest that the enhancement of reprogramming by cAMP treatment is attributable to the EPAC pathway rather than the PAK pathway. However, the effects of EPAC-specific cAMP treatment were not as dramatic as those achieved by treatment with 8-Br-cAMP or forskolin. This indicates that other pathways, in addition to the EPAC pathway, are involved in the enhancement of reprogramming efficiency by cAMP treatment. Moreover, these multiple downstream signaling mechanisms appear to exert their effects only during the early stages of induction of reprogramming (at about day 3 post-transduction).
We also showed that the enhancement of reprogramming by the cAMP pathway requires suitable medium conditions. Previous studies reported the role of the cAMP/PKA signaling in mESC self-renewal, proliferation, and differentiation [13,15]. Treatment with cell-permeable 8-Br-cAMP (1 and 5 mM) decreased the expression of Oct4 in the presence of LIF, but increased its expression in the absence of LIF. Forskolin had no effect on Oct4 mRNA levels during maintenance, but increased the expression of Oct4 during differentiation induced by treatment with retinoic acid. Interestingly, H89, a known PKA inhibitor, also reduced the expression of Oct4 in the presence of LIF, but induced the expression of this gene in the absence of LIF. Despite H89 being frequently used as a PKA inhibitor, it is also known to affect other protein kinases [32]. We showed that the increase in reprogramming efficiency in response to cAMP treatment differed according to the culture medium; a much greater increase in reprogramming efficiency was observed in cells cultured in serum-free medium than those in serum-based medium. The differing effects of cAMP treatment may be due to the various factors present in serum, which may interfere with the effects of 8-Br-cAMP treatment.
These observations are expected to contribute to the elucidation of the early events that occur during reprogramming and provide insights into the effects of signaling pathways for inducing pluripotency in differentiated cells, thereby helping to overcome the barrier of cellular potency.
Footnotes
Acknowledgments
This research was supported by the Basic Science Research Program of the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT and Future Planning (grant nos. 2015R1A2A2A01003604 and 20110019489) and the NRF funded by the Ministry of Education (grant nos. 2013R1A1A2011394 and 2011-355-C00114).
Author Disclosure Statement
No competing financial interests exist.