
Abstract
The origin of the wide spread node and duct system described by Rai et al. remains a mystery. The explanation came when another study on yolk sack hemopoiesis was compared with the “primo vascular system”. It came out that the yolk sack hematogenic tissue, introduced to the embryo by the vitelline veins, does not disappear in adults, but continues to exist in form of nods and ducts along blood vessels and different organs.
F
Part of my commentary [1] is on origin of hemopoietic system. The splanchnic mesoderm under the influence of differentiation factors (morphogens) differentiates into hematogenic tissue, whose cells are now hemangioblasts (Fig. 1). The capillaries fuse together forming a network on the yolk sac surface and finally are collected in the vitelline veins. Surrounded by the hematogenic tissue, the united vitelline veins enter the embryo. Here they continue as ductus venosus, which is enclosed by two tube-like liver diverticula. Endodermal cords of these liver diverticula proliferate on all sides of ductus venosus.
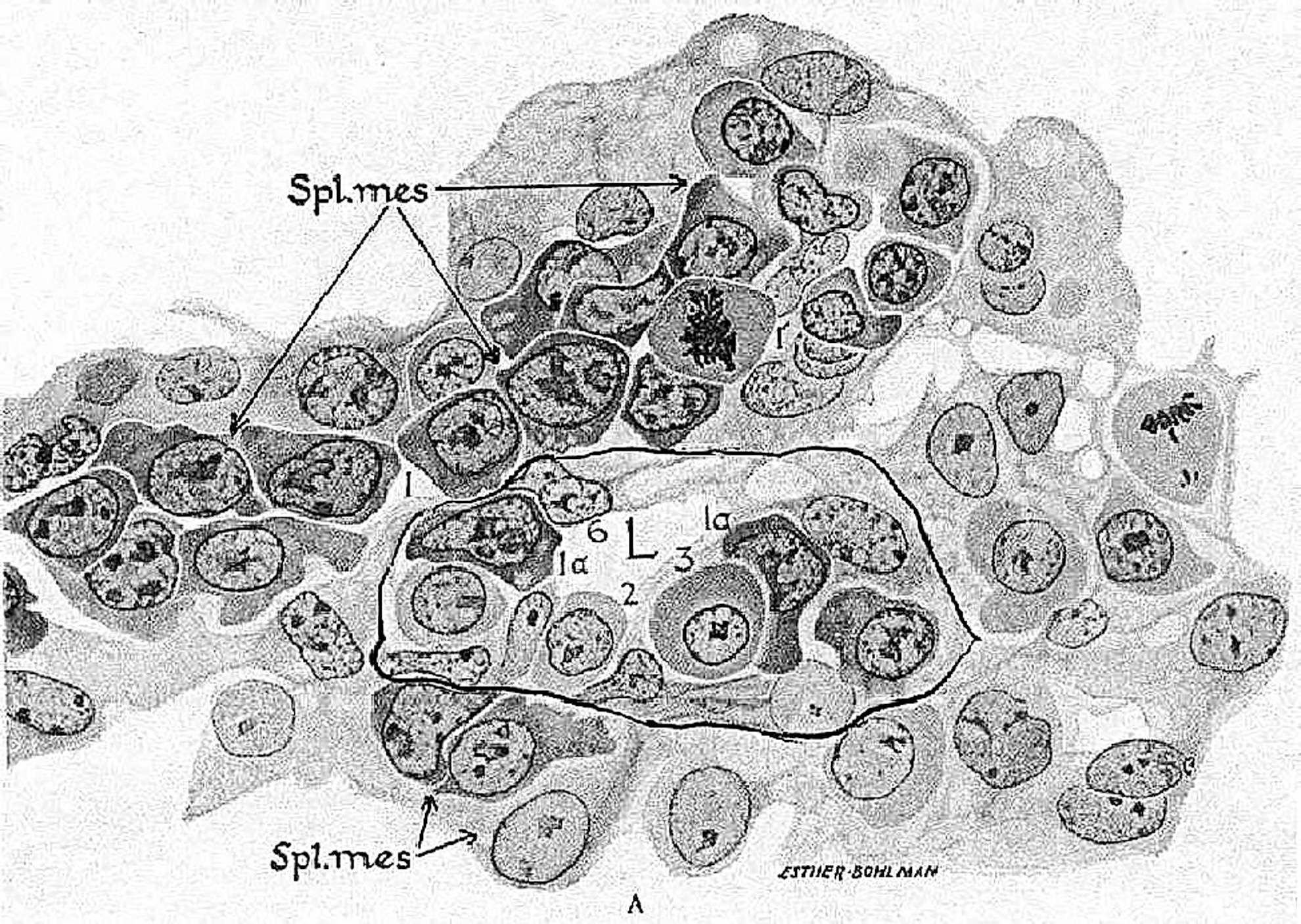
Blood island from a yolk sack of a 24-day human embryo. The splanchnic mesoderm differentiates into hematogenic tissue, consisting of hemangioblasts. In the center, a blood island contains endothelial and primitive erythrocytes. L, lumen of the capillary, surrounded by significant amount of hemangioblasts. Microphotography from Maximov A and Bloom W. (1957). A Textbook of Histology, 7th edition. W.B. Saunders Company, Philadelphia, PA and London.
This proliferation of liver trabeculae is accompanied by sprouting of capillaries from ductus venosus, which interdigitate with the endodermal liver cords in a complex manner. These branches of the vitelline veins dissipate into numerous hepatic sinusoids. Liver hemopoiesis originates from the vitelline vein capillaries bringing the hemangioblasts and forming blood islands-like structures between the endodermal liver cords. This is how the liver becomes the main hemopoietic organ during early fetal life. But the growth of the vitelline veins does not stop here. They continue to spread along the dorsal mesentery, branching into the spleen, mesonephros, cartilages at the beginning of ossification, and many other organs. This would explain why Medvinsky found temporary hematopoietic precursors in the para-aortic mesenchyme [3]. When transplanted in vivo or in organ cultures, the hemangioblasts undergo temporary differentiation. Medvinsky defends the idea that AGM (aorta, gonads, mesonephros) are the source for migratory stem cells seeding not only the adult but the blood islands in the yolk sack as well, “indicating migration of these cells in various directions.” Logically, this is inadmissible, because the blood islands produce erythroblasts before the connection of extra- and intraembryonic vessels. Hemopoiesis in the AGM area has never been observed. I was convinced that Medvinsky found the hematogenic tissue introduced from the yolk sack by ductus venosus, but I did not know what the morphological appearance of the hemangioblasts during late development is.
Now after I read the excellent article of Rai et al. [4], I was very pleased to see the morphology of the nodes and ducts, which explains what is the fate of the yolk sack hemangioblasts in adults. The vitelline vessels spread and give rise to numerous blood vessels, which are not named vitelline any more, but still containing the hemangioblasts, which become organized into nods and ducts system (NDS), as the fetus develops to adult individual. The cells found in NDS are hemangioblasts and all kinds of blood cells (erythrocytes, all types of granulocytes, and megacaryocytes) [4]. This is an indication that NDS is connected with the yolk sack hemopoietic tissue. There is no other possibility for the appearance of hemangioblasts and blood cells in NDS. A bridge connects the two studies, which reveal the origin and development of a new anatomical system. It is amazing how the two investigations complement each other. I did not know about the existence of the Bonghan corpuscles and ducts. However, the authors describing this system have not been familiar with the embryonic hemopoiesis. Combined, the two articles elucidate what is the origin of NDS, what is the fate of the embryonic yolk sack hemangioblasts, how they do a widespread network, and how they can contribute to new angiogenesis and regeneration.
Now the widespread Bonghan system is not any more a mystery. It enriches our knowledge about the existence of a new anatomical entity in adult individuals connected with late development of the yolk sack hemopoiesis, which is a completely new concept.
Footnotes
Author Disclosure Statement
No competing financial interests exist.