
Abstract
Our concept of cell reprogramming and cell plasticity has evolved since John Gurdon transferred the nucleus of a completely differentiated cell into an enucleated Xenopus laevis egg, thereby generating embryos that developed into tadpoles. More recently, induced expression of transcription factors, oct4, sox2, klf4, and c-myc has evidenced the plasticity of the genome to change the expression program and cell phenotype by driving differentiated cells to the pluripotent state. Beyond these milestone achievements, research in artificial cell reprogramming has been focused on other molecules that are different than transcription factors. Among the candidate molecules, microRNAs (miRNAs) stand out due to their potential to control the levels of proteins that are involved in cellular processes such as self-renewal, proliferation, and differentiation. Here, we review the role of miRNAs in the maintenance and differentiation of mesenchymal stem cells, epimorphic regeneration, and somatic cell reprogramming to induced pluripotent stem cells.
Introduction
E
Nuclear transfer consists of the transfer of the nuclei from a completely differentiated cell into an enucleated oocyte. This approach renders embryos and has allowed the cloning of animals from diverse species [3,4]. Fusion of two or more different cell types results in either proliferating hybrids or heterokaryons, which do not proliferate. Heterokaryons showed that previously inactivated and lineage-specific genes could be reactivated and expressed in other cell types, suggesting that a differentiated state is not irreversible and depends on the presence of certain molecular regulators.
It has been shown, using the mouse as an experimental model, that overexpression of four transcription factors OCT4, SOX2, KLF4, and C-MYC in completely differentiated cells enabled the generation of induced pluripotent stem cells (iPSCs). These cells not only expressed genes characteristic of the pluripotent state but also showed a similar morphology, culture requirements and allowed the formation of chimeras and germ line transmission [5,6].
Since Yamanaka's first report, factors used for the generation of iPSC have diversified. Among the recently tested molecules are microRNAs (miRNAs) [5], such as miR-200c, miR-302, and miR-369, which improve the reprogramming efficiency for iPSC generation [7]. miRNAs are small non-coding RNAs (sncRNAs) of 17–23 nucleotides in length, originally discovered in Caenorhabditis elegans [8]. The role of miRNAs in post-transcriptional gene regulation is highly conserved in animals. However, there are clear differences in miRNA biogenesis and their mode of action among kingdoms.
Animal miRNAs are transcribed by RNA polymerase II, giving rise to a primary transcript or pri-miRNA of several kilobases in length that is polyadenylated at its 3′ end and capped at its 5′ end. Pri-miRNAs possess characteristic stem-loop structures that render them suitable for further recognition and processing by RNase type III endonucleases [9]. The maturation process is then carried out by DROSHA, a member of the Rnase III family, which, together with the RNA-binding protein DGCR8, cleaves close to the double-/single-strand junction of the pri-miRNA. The resulting precursors (pre-miRNAs) are then exported from the nucleus to the cytoplasm by binding at their 3′ end to the Exportin 5 protein that mediates transfer through the nuclear pore complex [9,10].
In the cytoplasm, the pre-miRNA is processed to a mature miRNA by the action of DICER and its cofactors (extensively reviewed in Sand [9]). Pre-miRNAs will give rise to a stable and normally highly abundant mature miRNA strand, as well as to an ephemeral and much less abundant miRNA* strand. Once the miRNA is mature, it is loaded into the RNA-Induced Silencing Complex (RISC) and guided to its target messenger RNA (mRNA) [9,10].
Recognition of the target is achieved through an imperfect complementarity between the so-called miRNA seed sequence (originally described as the first seven nucleotides from the 5′ end of the mature miRNA) and the target sequence located at either the 3′ untranslated region (3′ UTR) of the mRNA or in coding sequences [8,9]. In animals, miRNAs control the expression of their targets by attenuating mRNA translation or by altering mRNA stability. miRNA-dependent fine regulation of gene expression influences several cell reprogramming events and developmental processes such as cell differentiation, cell proliferation, cell transformation, morphogenesis, and regeneration [12 –14]. In the next few paragraphs, we discuss different studies that evidence how miRNAs contribute to cell plasticity in diverse developmental contexts.
miRNA-Mediated Regulation of Mesenchymal Stem Cells Identity and Differentiation Capacity
Mesenchymal stem cells (MSCs) are a type of stem cells derived from adult tissues such as bone marrow, muscle, dermis, and adipose tissue. MSCs display a multi-lineage potential of differentiation, and thus, depending on the tissue of origin, these cell populations could give rise to adipocytes, neurons, cardiomyocytes, osteoblast, chondrocytes, or hepatocytes [15] (Fig. 1A). Because of their multipotency, they open the possibility of autologous cell transplantation, MSCs are relevant from a therapeutic point of view. However, due to the variety of tissues and pathways from which these cells are derived, a well-specified definition for MSCs does not exist.

Schematic representation of MSCs and their normal
Indeed, MSCs is a term that includes cell populations that meet a minimum of criteria: (1) MSCs show plastic adherence; (2) MSCs have expression of CD73, CD90, and CD105 and no expression of CD34, CD14 or CD11b, CD45, CD79a, CD19, or HLA-DR; and (3) MSCs are able to differentiate into osteoblasts, adipocytes, and chondrocytes in vitro [16].
To gain insight into the molecular events underlying the fate of the different MSCs populations, profiling of miRNAs was carried out by Trohatou et al. [17]. By collecting RNA samples of MSCs from amniotic fluid (AF-MSCs), bone marrow mesenchymal stem cells (BM-MSCs), and umbilical cord blood and using a microarray-based approach, they were able to find differentially expressed miRNAs, some of which were common to all MSCs sources whereas some of which were specific. Among their results, they found that miR-21 was the most upregulated miRNA in round-shaped AF-MSCs when compared with the other sources, and they identified SOX2 as a main target for this miRNA (Fig. 1A). The authors showed that not only miR-21 overexpression caused SOX2 inhibition, but also OCT4 and NANOG were downregulated in an indirect manner, resulting in an impairment of the proliferation rate and clonogenic potential.
Additionally, miR-21 triggered a G0/G1 cell cycle arrest along with a decrease in the protein level of Cyclin E1 and Cyclin A, as well as downregulation of CDC25. In parallel, the expression of cyclin-dependent kinase inhibitors was upregulated. Therefore, miR-21 appears to regulate MSCs proliferation at different levels of the cell cycle. Overexpression of miR-21 causes a decrease in adipogenic and chondrogenic differentiation, whereas it promotes osteogenic differentiation by a mechanism still not well understood [17].
A group of downregulated miRNAs, miR-143, miR-487, miR-326, and miR-199*, was identified in all MSCs sources, compared against the expression level measured on control experiments [17]. Although consequences of the downregulation of these miRNAs were not addressed by Trohatou et al., it has been reported that miR-143 has a role as a tumor suppressor in colon with RAS and ERK1/2 transcripts reported as miR-143 targets [18]. miR-326 is upregulated in multiple types of sclerosis and it is also involved in regulation of the differentiation of Th-17 cells [19], whereas members of the miR-199 family are involved in myocardial regeneration [20].
miRNA profiling of MSCs populations has also been conducted in adipose tissue-derived MSCs (AMSCs) and bone marrow (BMSCs) of old and young human individuals [21]. The most downregulated miRNA in BMSCs was miR-935, whereas miR-153 was the most downregulated in AMSCs, both of which were compared against the expression measured in other MSCs sources. Notably, members of the miRNA family let-7 were also downregulated as a function of the age in both AMSCs and BMSCs. The most upregulated miRNAs were miR-510 in BMSCs and miR-335 in AMSCs [21].
Because these reported miRNAs have targets that are found in other organisms, it is interesting to explore whether they also perform similar roles in their respective MSCs populations. For instance, SLC5A3 (Solute carrier family 5 member 3), a co-transporter that maintains the intracellular levels of myo-inositol, is a target of miR-935 in medulloblastoma cells, where it acts by affecting phosphatidylinositol synthesis and probably alters PI3K signaling, which is an important pathway mediating cell growth, invasion, and apoptosis resistance in cancer [22].
It has recently been reported that overexpression of let-7 increases osteogenic differentiation in human AMSCs, whereas it also suppresses adipogenic differentiation. This regulation was mediated by the HMGA2 gene that is a target of let-7 and encodes the High Mobility Group AT-Hook 2 protein [23]. This protein has roles in the regulation of body size, growth of cardiomyocytes, and adipogenic differentiation [24]. Inhibition of the endogenous HMGA2 reproduces the effect of let-7 overexpression, suggesting that HMGA2 mediates the effect of let-7 on osteogenic/adipogenic differentiation [23].
Studies of Pandey et al. showed that let-7 is downregulated in relation to the age of the organism [21]. This insight, together with Wei's data supporting the ability of let-7 to promote osteogenic differentiation, indicates that the let-7/HMGA circuit acts on the age-dependent decrease of osteogenic capability and increases adipogenic differentiation.
miR-335 is another age-dependent upregulated miRNA, the expression of which decreases during BM-MSCs differentiation. Overexpression of miR-335 in human MSCs (hMSCs) causes a reduction in proliferation without increasing apoptosis, as well as a decline in the migratory response to serum and an in vitro wound-healing test. In addition, both adipogenic and osteogenic differentiation are also reduced, particularly, osteogenic differentiation was almost abolished by the action of miR-335, which appears to target Runt-related transcription factor 2 (RUNX2), an osteogenic transcription factor [25]. These results are in agreement with the age-dependent decrease of osteogenic differentiation, since an age-dependent increase of miR-335 potentially inhibits such a developmental process.
Different studies have recently shown that specific miRNAs can be employed to direct MSCs differentiation to a specific fate. In humans, mesenchymal/stromal stem cells (hMSC) have been proposed as an ideal source for tissue engineering and tissue regeneration strategies [26]. Liver and pancreas degenerative diseases are responsible for millions of deaths annually worldwide. Searching for alternative treatments, molecular medicine has focused on miRNAs, which have been shown to be effective for the reprogramming of hepatocytes and insulin-producing cells (IPCs) from MSCs.
In 2013, microarray analysis identified a group of miRNAs, miR-1246, miR-1290, miR-148a, miR-30a, miR-424, and miR-542-5p, as specifically upregulated during hepatic differentiation of hMSCs [27]. Later that year, this group of miRNAs, in combination with miR-122 (highly expressed in liver), was used to induce hMSCs to hepatocyte-like cells (iHep; Fig. 1B). Transplantation experiments in injured mouse liver showed that transplanted iHep resulted in the improvement of liver function, as evidenced by an increase in the concentration of hepatic function enzymes (ALT and AST) and the concentration of serum albumin [28].
Recently, MSCs from bone marrow samples of healthy humans were reprogrammed to IPCs by the overexpression of miR-375 and the concomitant downregulation of miR-9 (Fig. 1B) [29]. MSCs acquired an identity of IPCs by expressing PDX1, NGN3 (as markers of pancreatic development) and by the production of insulin and glucagon at day 21 after lentiviral transduction. IPCs were able to secrete insulin in response to glucose in a dose-dependent manner [30]. miR-375 is an miRNA previously implied in pancreatic islet development [31], whereas miR-9 is a negative regulator of insulin secretion from IPCs in response to glucose by targeting the OC-2 transcription factor [32]. Despite these interesting reports, the reprogramming mechanisms of MSCs to IPCs as well as the in vivo functionality of IPCs should be investigated in more depth.
Cartilage has a poor regenerative capacity, and millions of people suffer from articular cartilage damage [33]. Devising strategies to reprogram MSCs and to induce chondrogenic differentiation is, therefore, an intense area of research. Among the molecular candidates that influence such differentiation processes in a positive manner are the miRNAs miR-210 and mir-630 that are positive regulators of chondrogenesis [26,34]. miR-210 alters mitochondrial metabolism by targeting and inhibiting the expression of iron-sulfur (Fe-S) cluster scaffold proteins, ISCU1 and ISCU2 [35]. miR-630 directly and indirectly targets both MAP-Kinase/Erk signaling and TGFb signaling, which mediate chondrogenic differentiation.
miR-140 also stimulates in vitro chondrogenesis by downregulating the protein levels of the small GTPase RALA in a regulatory network where SOX9 and ACAN are upregulated by the indirect upstream action of miR-140 [36]. On the other hand, miR-181, miR-34a, and miRN-495 are negative regulators of chondrogenesis [26,34] and the downregulation of miR29a and the upregulation of its target FOXO3a are necessary for MSCs differentiation into chondrocytes [37].
miRNA-mediated adipogenic differentiation from MSCs has also received significant attention. Recent findings of such a differentiation process are summarized in a review by Hamam et al. [38]. In this review, the role of miRNAs directly or indirectly regulating adipogenic differentiation of MSCs is explained by the targeting of various genes. Indirectly, adipogenic differentiation may be influenced by the action of miR-143 on its targets ERK5 and MPAPK7. Also, miR-355 targets RUNX2), a pivotal player in osteoblast and chondrocyte differentiation. Finally, some miRNAs directly regulate adipogenic differentiation; this is the case of miR-146b, which targets Sirtuin 1 (SIRT1), an inhibitor of PPARγ. PPARγ is a key factor in the molecular network that controls adipogenesis by regulating several adipocyte-related enzymes.
Cell Plasticity and Cancer Stem Cell Identity
Among the most studied cell transformation events are those that reprogram a normal cell into a carcinogenic one, with the potential to generate a malignant tumor. How this occurs and how a tumor is maintained are major unsolved questions. The cancer stem cell (CSC) hypothesis has been widely accepted to explain the maintenance of a tumor [39 –41]. This theory tackles the question of why a tumor might regrow after surgery and why therapy that is focused on proliferative cancer cells is not always successful. The potential to generate a new tumor is sustained for a limited amount of cells in the tumor tissue called CSCs. Similar to other stem cells, CSCs have the capacity to divide asymmetrically, to self-renew, and to give rise to all the cell types present in the tumor. The CSC theory also establishes that tumoral tissue shows a hierarchical developmental organization that correlates the heterogeneity of the cells found in the tumor with several degrees of differentiation [40]. If a CSC is able to generate a tumor, how does the CSC originate? There are at least two possible scenarios that can explain the acquisition of the CSC identity. One possibility is that an adult stem cell goes through a malignant transformation and becomes a CSC [42,43] (Fig. 2A). The other possibility is that a differentiated cell dedifferentiates and acquires stem-cell-like properties [40 –44] (Fig. 2B). Regardless of the path a cell might take to become a CSC, the process involves an event of molecular reprograming, where miRNAs may play a role either in the maintenance of a differentiated status or in the transformation into a cancer cell [45]. In the next section, we review recent studies demonstrating the involvement of miRNAs in the formation and maintenance of CSCs.
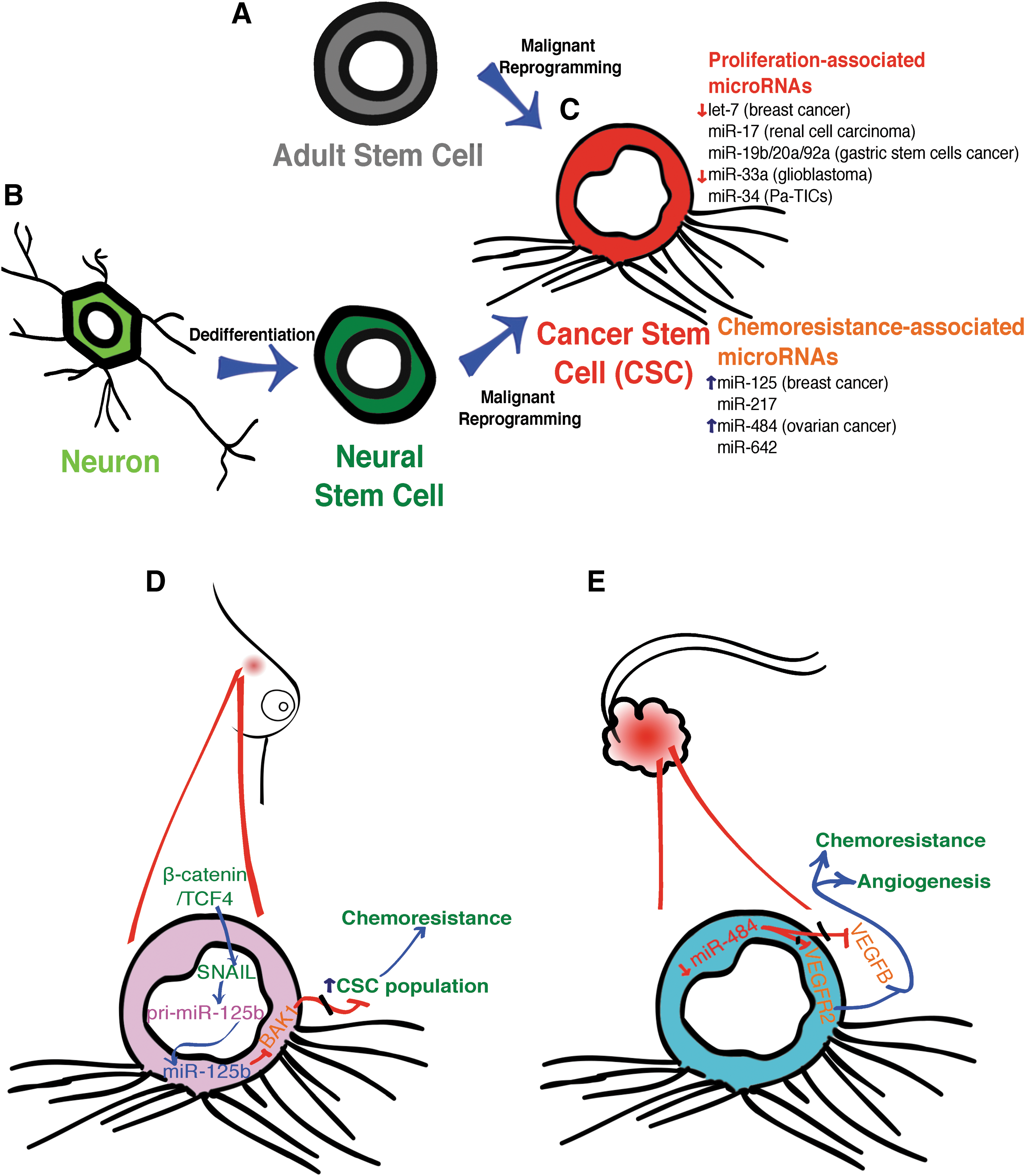
Schematic representation of CSCs and how these cells are originated. Two different possibilities have been hypothesized to give rise to CSCs. In the first possibility
The contribution of miRNAs to the regulation of the CSC state
The role of miRNAs in the regulation of CSC attributes such as self-renewal, clonogenicity, and tumorigenicity has been clearly established for several types of cancer (Fig. 2C). One of the first miRNAs reported to be involved in CSC fate was let-7 in breast cancer cells. let-7 Expression is downregulated in breast tumour-initiating cells (BT-ICs) when compared with breast cancer cell lines [46]. Low expression of let-7 correlates with an increase of its targets, RAS and HMGA2 [47]. RAS contributes to self-renewal, whereas HMGA2 sustains a multipotent differentiation capability. Silencing of RAS decreases self-renewal properties, as it reduces mammosphere formation as well as clonal expansion. In contrast, silencing of HMGA2 disrupts the proportion of undifferentiated cells in BT-ICs cultivated as mammospheres or in a monolayer [46]. A similar role was demonstrated for miR-34 in pancreatic tumour-initiation cells (Pa-TICs), a subpopulation of the pancreatic cancer cell line MiaPaCa2. Pa-TICs express surface markers CD44+/CD133+ [48], and they are characterized by low levels of miR-34 and high levels of miR-34 targets, including BCL2, NOTCH1, and NOTCH2. BCL2 encodes an anti-apoptotic protein [49], whereas NOTCH1 and NOTCH2 encode receptors of the Notch signaling pathway (reviewed extensively in Aithal and Rajeswari [50]). An increase in the level of miR-34 results in decreased self-renewal capability of Pa-TICs, which was measured by tumorsphere formation in vitro, and tumor formation in mice [48]. miR-34c is downregulated in BT-ICs due to hypermethylation of two sites in the promoter region of the miR-34 cluster. Hypermethylation decreases the capability of Specificity Protein 1 (SP1) to bind to the miR-34c promoter and to induce miR-34c expression. Similar to Pa-TICs, miR-34c restoration in BT-ICs suppresses self-renewal, reduces the number of mammosphere formation, and decreases the population expressing BT-ICs markers [51].
The regulation of self-renewal and tumor formation competence by miRNAs has also been documented in several cancers, such as miR-33a in glioblastoma, miR-17 in renal cell carcinoma, and miR-19b/20a/92a in gastric CSCs [52 –54]. In addition to self-renewal and tumor formation capability, chemotherapy resistance and metastasis are further characteristics that are associated with CSC.
In 2004, Hirschmann-Jax et al. identified the equivalent of the “side population” (SP) found in several kinds of mammalian stem cells in neuroblastoma cancer tumors collected from patients [55]. SP from neuroblastoma tumors shows a remarkable capacity to excrete cytotoxic drugs. Such capacity was based on the high expression of ABCG2 and ABCA3 transporters. Neuroblastoma SP also displays stem-cell properties such as self-renewal and the ability to generate SP and non-SP cells. SP for other types of cancer, including mammary gland adenocarcinoma, ovary teratocarcinoma, kidney Wilms tumor, Ewing sarcoma, and small-cell carcinoma of the lung, has also been identified [55]. This study provided, for the first time, evidence of the existence of the SP showing characteristics of CSCs, and this, together with their capacity to efflux drugs, may help explain cancer relapse due to resistance to chemotherapy [55].
The regulation of such chemoresistance by miRNAs has been documented in the case of ovarian cancer, where a particular expression profile of miRNAs has been associated to it (Fig. 2C). In this study, a microarray approach linked 23 miRNAs to chemoresistance and the subsequent/consequent/expected misregulation of miR-484, miR-642, and miR-217 was experimentally validated. Overexpression of miR-484 in ovarian cancer cell lines inoculated in nude mice confers in vivo sensitivity to carboplatin and taxol. This effect was mediated by two miR-484 targets, VEGFB (Vascular Endothelial Growth Factor B) and VEGFR2 (Vascular Endothelial Growth Factor Receptor 2), both of which promote angiogenesis in the tumor microenvironment. Inhibition of VEGF signaling by miR-484 results in impairment of the tumor vascular system and an increase of necrosis in tumoral tissue (Fig. 2E). Based on these results, it was proposed that blocking VEGF signaling could improve ovarian cancer drug sensitivity [56].
Another example of chemoresistance modulation by miRNAs is provided by miR-125b in breast cancer (Fig. 2D). This effect is mediated through SNAIL, a zinc finger transcription factor that activates miR-125b promoter in a β-catenin/TCF4-dependent fashion. miR-125b inhibits the translation of BAK1, a pro-apoptotic protein [57], leading to an increase in chemoresistance. In parallel, overexpression of miR-125 promotes chemoresistance by increasing the CSC population within the breast cancer cells. This effect is also mediated by the BAK1 protein [58].
In summary, miRNAs participate in the establishment and maintenance of the CSCs program and also modulate the capacity of these cells to resist chemotherapy. These features make miRNAs potential molecular targets for the treatment of a variety of cancers through the disruption of CSCs population maintenance.
Cell Plasticity and Epimorphic Regeneration of Complex Structures
Epimorphosis is a process through which some organisms are able to re-establish an injured organ or member [59]. This process involves the formation of a blastema, a group of proliferative cells from which the injured tissue will regrow [55]. Epimorphic regeneration in amphibians has been divided into three phases: wound healing, dedifferentiation, and redevelopment [60,61] (Fig. 3C). The blastema is a structure formed by a heterogeneous group of progenitor cells with a restricted potential for differentiation. Evidence from newt species and Ambystoma mexicanum (axolotl) suggest that such progenitor cells result from the reprogramming of completely differentiated cells that experiment with some degree of dedifferentiation [62,63].

This can also be observed during the regeneration of the caudal fin in zebrafish, where lineage tracing experiments demonstrated that osteoblasts dedifferentiate and contribute to the blastema cells. These osteoblasts differentiate and replace only bone tissue, showing the restricted potential of blastema cells to differentiate in a specific cell lineage [64]. This way, epimorphic regeneration involves several reprogramming events; a dedifferentiation step is followed by a differentiation phase, suggesting that blastema cells should be highly plastic. Such plasticity depends on the regulation of gene expression at different levels [65]. miRNAs provide a level of fine-tuning the regulation of gene expression needed for practically all reprogramming events, including those occurring during epimorphic regeneration.
The role of miRNAs during the regulation of cell plasticity in a regeneration process was first investigated by Tsonis et al. in 2007 [66]. In adult newt, the regeneration of lens and inner ear hair cells involves a transdifferentiation step, defined as the conversion of the identity of a completely differentiated cell into another cell type [65]. Using microarrays, they identified miRNAs differentially expressed between intact irises and regenerating irises. let-7 was highlighted, because it was downregulated in regenerating irises and in addition, let-7 directly targets oncogenes such as myc, hmga, and ras [47]. In this way, let-7 downregulation would allow cell proliferation during lens regeneration [66]. The relevance of this study is based on the establishment of miRNAs as modulators of cell plasticity for a regenerative process.
In the next paragraph, we summarize the studies performed to unravel the role of miRNAs in epimorphic regeneration for the recovery of complex structures such as tails, fins, and limbs.
miRNAs associated with epimorphic regeneration
The involvement of miRNAs in epimorphic regeneration was first described by Yin et al. in 2008 [67]. Using an miRNA microarray approach, the authors discovered a set of miRNAs that showed differential expression during fin regeneration in zebrafish. The expression levels of these miRNAs appeared to depend on the fibroblast growth factor (FGF) signaling. FGF signaling is an essential pathway for regeneration, since it regulates cell proliferation of the blastema cells [68]. From the set of miRNAs, the authors focused on miR-133, which displayed high levels of expression in the un-injured adult caudal fin but it was downregulated during blastema formation and low levels of miR-133 remained until 14 days post-injury [67].
Although the mechanism by which FGF signaling downregulates miR-133 has not been described, this event promotes the expression of the miR-133 target Mps1 gene encoding a protein kinase. Mps1 function is related to the spindle checkpoint, when all chromosomes should be attached to the mitotic spindle before cells proceed to anaphase during mitosis [69]. The ultimate effect of miR-133 downregulation and the concomitant upregulation of Mps1 is the proliferation of blastema cells for successful regeneration of the fin [67].
Thatcher et al. demonstrated that a crucial component of the miRNA biogenesis pathway is required for regeneration, since inhibition of Dicer using morpholinos resulted in the inhibition of regeneration [70]. The same laboratory identified differentially expressed miRNAs from microarray experiments in adult fins, regenerating fins and fins with apparently complete regeneration. From this study, miR-203 was identified as an interesting candidate based on its strong downregulation and on the identity of one of its targets, lef1 gene encoding a transcription factor that is regulated by the Wnt signaling pathway [71]. The Wnt pathway has been shown to be involved in controlling the formation of the basal epidermal layer and proliferation in underlying cells, which will give rise to the blastema in A. mexicanum, zebra fish, and Xenopus [71, 72].
In fact, Lef1 is a marker of the basal epidermal layer in blastema formation [58,71]. Thatcher et al. demonstrated that miR-203 targets the 3′ UTR of lef1, and the negative regulation of lef1 is a limiting step for fin regeneration [70]. The studies described earlier indicated an intricate regulation of miRNAs during blastema formation and revealed a crosstalk between essential signaling pathways such as FGF and Wnt that are involved in regeneration.
The role of miRNAs in the regulation of epimorphic regeneration was also explored in the axolotl by monitoring changes in the expression levels of a set of highly conserved miRNAs in samples of 3 day blastema in regenerating tail. From this screening, the miR-196 family was the most significantly upregulated at this time point. miR-196 induces pax7 downregulation by targeting its 3′ UTR [73]. Pax7 is restricted to the dorsal region of the spinal cord [74,75]. Inhibition of miR-196 results in shorter regenerated tails and when miR-196 is inhibited, the expression level of Pax-7 remains high and its expression domain appears to be expanded, suggesting that miR-196 could play a role in the correct patterning during tail regeneration [73].
Meis2, a protein involved in the proximo-distal patterning in limb regeneration [76], is downregulated on inhibition of miR-196. Meis2 is highly expressed in the proximal region relative to distal blastema during A. mexicanum limb regeneration. It has been hypothesized that Meis2 serves as a signal to determine how much tissue should be regenerated [69].
More recently, the role of miR-125 during axolotl tail regeneration has been investigated by the Echeverry lab [77]. Comparative analyses between rat (a mammal with a limited capacity of regeneration) and axolotl revealed that the levels of miR-125b were eight times higher in axolotl during spinal cord regeneration, and a sudden decrease of up to 40% in the levels of miR-125b was observed in axolotl whereas in rat the decrease was only 1%. Based on this observation, it was hypothesized that a significant and earlier reduction in the levels of miR-125 in axolotl relative to the rat was important for the upregulation of miR-125 targets, which might be necessary for regeneration [77].
To test this hypothesis, the authors artificially increased miR-125 levels in axolotl (which under normal conditions would be decreased). An increase of miR-125 led to defects in sprouting of axons; whereas an inhibition of miR-125b (to levels comparable to those observed in rat) did not hamper axonal regeneration but, in contrast, it promoted axon degeneration at the caudal site of the injury. Deposition of fibrin that resembles the formation of a glial scar in non-regenerative species was also observed [77]. These results collectively indicate that precise modulation of miR-125 levels is necessary for proper regeneration.
Semaphorin 4D (sema4D) mRNA was identified as a target of miR-125b. Sema4D is expressed in oligodendrocytes and upregulated near a spinal cord injury. Overexpression of Sema4D inhibits axonal regeneration in axolotl [73]. In addition, Sema4D inhibits the growth of cerebellar and sensory neurites in vitro [78]. Furthermore, early overexpression of miR-125 in rats caused a downregulation of Sema4D that created a permissive condition for spinal cord regeneration with an improvement in locomotion abilities and a decrease in GFAP-positive scar tissue. Similar to axolotl, miR-125 overexpression in rats improved the regeneration capability of the animals.
However, Sema4D is not the only target of miR-125, as it modulates the expression of further 23 target genes; among them, there are genes linked to glial scar formation, microtubule stabilization, neurite outgrowth, and axon guidance, whose downregulation and effects on regeneration are awaiting to be uncovered [77].
In the context of limb regeneration in axolotl (Fig. 3C), a recent study focused on the medium limb blastema stage to identify miRNAs that regulate the patterning of the new limb. The study showed that at this stage miR-21 was the most expressed miRNA relative to a non-regenerating stump [79]. miR-21 has several putative targets, but because of its implication in limb patterning during embryonic mouse development, authors focused on the Jagged1 gene whose 3′ UTR has a single target site for miR-21 [79, 80].
In the medium limb blastema, upregulation of miR-21 correlates with the absence of Jagged1 protein and it was speculated that downregulation of Jagged1 by miR-21 could repress the proliferation of blastema cells and accelerate differentiation, thereby enabling commitment to a particular cell fate [79] (Fig. 3C). In agreement with this hypothesis, several human cancers have shown that inhibition of Jagged1 decreases cell proliferation whereas its upregulation has an oncogenic effect [81 –83]. Jagged1 acts as a ligand of Notch signaling that effectively participates in cell differentiation and patterning [84].
Our current knowledge of the regulation of cell dedifferentiation/differentiation in epimorphic regeneration mediated by miRNAs is still scarce. To gain insight into the molecular events involved in the miRNA-based control of gene expression during the recovery of a member, it is necessary to dissect the mechanisms involved in the expression and interplay between miRNAs and their putative targets during distinct stages of regeneration of the caudal fin in zebra fish as well as the tail and limbs in axolotls.
miRNAs and the Regulation of the Pluripotent State
miRNAs expressed specifically in mouse and human embryonic stem cells (mESC and hESC respectively; Fig. 3A) have been reported, and a subset of miRNAs whose expression is restricted to the pluripotent state has been identified. In 2003, Houbaviy et al. [85] characterized three libraries of miRNAs prepared from different sources: (1) RNA from mESCs cultivated in conditions that allow the maintenance of the pluripotent state in culture, (2) mESCs grown under conditions supporting pluripotency less effectively than those used for the first source but, unlike the first, the second source had no contamination from feeder cells, and (3) mESCs cultivated in conditions inducing differentiation. Degree of differentiation under each condition was assessed by monitoring mRNA levels of Oct-4.
Sequencing of the isolated small RNAs resulted in the identification of 53 interesting miRNAs, from which a cluster of six miRNAs were suggested to be specific of mESC: miR-290, miR-291, miR-292, miR-293, mir-294, and miR-295. These miRNAs were expressed in mESC grown on feeders plus LIF, whereas they were downregulated or not expressed after in vitro differentiation as embryoid bodies during 14 days of culture [85]. The expression of those miRNAs was absent in mouse adult tissue, suggesting that these species are specific for the pluripotent state.
Using hESCs, a similar study was performed by Suh et al. in 2004 [86]. cDNA libraries from hESC grown under pluripotent conditions were generated. The level of differentiation was assayed by reverse transcription polymerase chain reaction of OCT4 mRNA levels as well as by the expression of cell surface markers of pluripotency SSEA-1, SSEA-3, and SSEA-4. Thirty-six potential miRNAs were identified from a set of 733 non-redundant sequences. From these 36, 16 corresponded to previously identified miRNA sequences, as detailed by Shu et al. in 2004 [86]. Among the remaining 20, 3 are common to the earlier described in mESC: miR296, miR-301, and miR-302a and 17 were different.
Interestingly, in this study, a cluster of miRNAs composed by miR-302b, miR-302c and miR-302a, and miR-302d and miR367 are expressed in both mESC and hESC and may have conserved roles in mammalian pluripotent stem cells; whereas miR-371, miR-372, and miR-373 are identified as the human homologs of the mouse miR-290 cluster [86].
Several studies have been carried out to understand how miRNAs specify the pluripotent state and contribute to the maintenance of pluripotency. We discuss those reports that involved pluripotent-state-specific miRNAs in a pathway or process relevant for the maintenance of pluripotency. Recently, it was shown that pluripotency transcription factors bind to the promoters and control the expression of certain ESC-specific miRNAs. Quantitative real time-PCR assays showed a reduction in the levels of pri-miRNAs-290, 295, 302, and 367 when Oct4 is depleted in mESC [87], revealing a positive feed-forward loop for stemness maintenance.
In addition, miR-302 was reported as a target of the Wnt/β-catenin pathway, which is a well-known regulator of mESC pluripotency [88,89]. Transcription of miR-302 is indirectly regulated by β-catenin through two non-canonical Tcf/lef binding sites at the miR-302 promoter. Tcf3 binds to such sites and acts as a repressor of miR-302 but on translocation of β-catenin to the nucleus, β-catenin releases Tcf3 from the Tcf binding sites at the miR-302 promoter, allowing miR-302 transcription [90]. This evidence indicates that at least a part of the effects of the pluripotent transcription factors are carried out through miRNAs and hence, miRNAs are capable of orchestrating the pluripotent and/or the differentiation status of the cells.
Regarding the downstream targets of the specifically expressed miRNAs in ESCs, evidence suggests that the miR-290 cluster acts at several levels. It controls the epithelial-to-mesenchymal transition (EMT), a cellular mechanism that throughout vertebrate embryonic development allows the migration of cells during gastrulation, delamination of neural crest cells, somite dissociation, and palate and lip fusion [11]. During EMT, cells lose their epithelial characteristics, acquiring migratory mesenchymal features; this process allows the pluripotent cells in the epiblast to acquire the identity of each of the three germ layers during gastrulation [91]. miR-290 cluster targets the NFκB subunit p65 at its coding region and contributes to the maintenance of pluripotency by translationally repressing p65 and preventing the activation of the canonical NFκB pathway that triggers differentiation of mESC by induction of EMT [11].
Activity of miR-294 and miR-302 interferes with the G1/S restriction point and maintains mESCs in an actively dividing state. These so-called ESC cell cycle miRNAs (ESCC miRNAs) target regulators of the G1/S restriction point, including CDKN1A, RBL1, and RBL2 [92,93]. Another mechanism associated with ESCC miRNAs is the antagonism of miRNAs that are able to induce differentiation. Among these are let-7, miR-26a, miR-99b, miR-193, miR-199a-5p, and miR-218; the mechanism of such antagonism has been only partially explored, but it has been proposed that miR-294 antagonizes the activity of let-7, as shown by the change in the expression levels of c-MYC (a target of let-7 ), through an unknown intermediate repressor of the let-7 activity [93,94].
In hESC, the most highly expressed cluster is miR-302/367; this cluster has a dual role, as it regulates cell-cycle progression while inhibiting apoptosis [95]. Regulation of the hESCs cell cycle involves the positive transcriptional regulation of OCT4/SOX2 that binds to the miR-302 promoter and induces its transcription. miR-302 translationally represses CYCLIN D1 and CDK4 and promotes ESC-like cell cycle with a long S phase and a short G1 phase [96]. It is worth mentioning that during mouse embryonic development, the expression pattern of Oct4 is similar to that of miR-302 in the epiblast [96]. These data suggest that regulation of the pluripotent cell cycle through OCT4/miR-302 and CYCLIN D1 could also be relevant during mouse embryonic development.
In parallel, miR302/367 directly targets two sites in the 3′ UTR of the pro-apoptotic gene BNIP3L and this way, miR302/367 inhibits apoptosis in hESCs. Both cell-cycle progression and apoptosis inhibition promote self-renewal of hESC [95].
A role for the miR-290/295 cluster in the regulation of apoptosis in mESC has also been reported. This dominant cluster directly targets apoptotic regulators Caspase 2 and Ei24, the latter of which is a transcriptional target of p53 [97].
Although several studies have been carried out to identify targets of ESCs-specific miRNAs, not all targets have been experimentally validated and placed in the context of a pathway [98 –100]. Also, it has to be taken into account that there are both redundant and unique miRNAs participating in the fine-tuning of pluripotency, depending on the ESC population that may vary not only among species but also depending on the ESC state in the same species [101]. Despite recent findings contributing to our current knowledge of the mechanisms by which ESC-specific miRNAs modulate the expression of genes involved in the maintenance of pluripotency in mESCs and hESCs, functional characterization of many putative targets of ESC-specific miRNAs needs to be carried out to understand how they regulate the pluripotent state.
miRNA approaches for inducing somatic cell reprogramming to the pluripotent state
In addition to reports of ESCs-specific miRNAs, it has been also demonstrated that certain miRNAs improve the efficiency of obtaining iPSCs (Fig. 3B). For instance, expression of mimic miR-294 in combination with Oct4, Sox2, and Klf4 enhanced the efficiency of iPSC formation from mouse fibroblasts. The miR-294 iPSC line allowed chimera formation, the irrefutable proof of pluripotency along with germ line contribution [102].
In a more recent approach, expression of the miR-302/367 cluster using lentivirus was combined with an inhibition of the histone deacetylase HDAC2 to reprogram hESC and mESC, with a higher efficiency than OKSM [7]. A disadvantage of using lentiviral vectors is the possibility of transgene integration in the host genome. Just 2 months later, an approach that avoids transgene integration in the host genome was reported [103]. In this study, reprogramming of mouse and human somatic cells was achieved by transfecting mature miR-200c, miR-302, and miR-369, an approach that represents great potential for clinical applications [104].
Knocking down components of the miRNA biogenesis machinery such as Ago2, Dicer, and Drosha decreases reprogramming efficiency of mouse fibroblast with OKSM [105]. Dicer-null mouse fibroblasts fail to produce ESC-like cells [104], providing additional evidence that miRNAs play a remarkable role in somatic cell reprogramming. However, the mechanisms by which miRNAs promote the reprogramming of somatic cells to iPSCs are still unclear.
Somatic cell reprogramming has been described as a multistep process divided into initiation, maturation, and stabilization phases. During the initiation phase, mesenchymal-to-epithelial transition (MET) is required for the acquisition of the proper epithelial characteristics of ESCs. In fact, blocking MET impairs the reprogramming process of mouse fibroblasts [106].
In accordance to the relevance of MET in reprogramming, another study showed that BMP signaling stimulates reprogramming by accelerating the progression to the maturation phase by the action of the BMP7 gene product. The BMP7 protein enhances the expression of miR-205 and miR-200 families, which, in turn, directly inhibit the translation of Zeb1 and Zeb2. ZEB1 and ZEB2 are transcriptional repressors of the E-cadherin gene that encodes a protein involved in cell adhesion in epitheliums (Fig. 3B). Therefore, BMP7 promotes MET by enhancing the expression of E-cadherin and the acquisition of epithelial markers in mouse reprogrammed cells [107]. The mechanism by which BMP7 increases the expression of those miRNAs has not yet been described.
A more recent study demonstrates that OCT4 and SOX2 positively regulate the expression of the miR-200 family in the mouse. Both OCT4 and SOX2 bind to the promoter region of the miR-200 cluster and activate its transcription [108]. A similar mechanism of MET/EMT regulation is observed in hESC (Fig. 3B). Human miR-302b and miR-372 target the 3′ UTR and inhibit the translation of the TGFBR2 mRNA that encodes the type II receptor of the transforming growth factor. TGFBR2 enables carcinoma cell migration by stimulating EMT [109]. As a consequence, miR-302b and miR-372 inhibit TGFβ induction of EMT by decreasing the levels of ZEB1 and ZEB2, thus allowing the upregulation of epithelial markers [110]. Taken together, these studies provide evidence on the importance of miRNa-mediated MET regulation for iPSCs generation from human and mouse somatic cells, revealing conservation of the process among species.
Specific miRNAs are also expressed during the initiation phase of mouse fibroblast reprogramming. For instance, miR-181 inhibition results in a 50%–60% reduction of reprogrammed colonies [100]. Potential targets of miR-181 were assessed, and seven of them were confirmed: Bptf, Cpsf6, Nr2c2, Bclaf1, Nol8, Ig2bp2, and Marcks. Proteins encoded by these genes showed downregulation during day 3 of reprogramming, when endogenous miR-181 is activated and its targets are repressed, indicating that they act as barriers for reprogramming. However, a detailed mechanism and functional validation of miR-181 targets and its role in modulating reprogramming remain to be elucidated [100].
miR-29b transcription is also induced early during reprogramming in mice cells by SOX2 that binds directly to the miR-29b promoter. As a consequence, mir-29 inhibits the translation of Dnmt3a and Dnmt3b genes encoding de novo DNA methyltransferases [111] (Fig. 3B). Translational inhibition of DNA methyltransferases results in reduced DNA methylation levels on the promoter of the Cdh1 gene, increasing E-cadherin expression and triggering MET [111]. Furthermore, mESC-specific miRNAs are also involved in the stabilization phase of the cell reprogramming process. For example, overexpression of the miR-302 cluster in partially reprogrammed cells allows the transition to full-reprogrammed cells.
It has been shown that miR-302 targets the 3′ UTR of MBD2 [112], which encodes a DNA-binding protein that binds selectively to methylated DNA and mediates the suppression of its targets. Partially reprogrammed cells show high levels of MBD2 on OCT4 and NANOG promoters, and it was demonstrated that overexpression of the miR-302 cluster leads to downregulation of MBD2, enables NANOG expression, and generates fully reprogrammed iPSCs [112].
Moreover, miR-302 is also involved in the modulation of OCT4 levels by inhibiting NR2F2 translation. NR2F2 is a transcription factor that binds directly to the promoter of OCT4 and represses its transcription. Therefore, miR-302 also promotes reprogramming through indirect induction of OCT4 gene expression [113]. Studies described earlier are representative examples of how miRNAs promote cell reprograming to pluripotency by translationally regulating several proteins involved in the regulation of MET/EMT as well as by key pluripotency transcription factors.
Concluding Remarks
The body of knowledge accumulated through decades of research has evidenced the ability of a completely differentiated cell to reprogram its differentiation status. This reprogramming relies on the ability of the nucleus to change the gene expression program of the cell, as elegantly demonstrated by Gurdon in 1962 [3]. The view that the differentiation status could be reversible led to the development of many reprogramming techniques that have undercover the plasticity of a cell to change its program and to acquire distinct differentiation states or identities. In fact, a cell can change its expression program in the context of an adult organism, as in the case of cell malignant transformation to a CSC [43,44], or a degree of dedifferentiation followed by the re-development phase in epimorphic regeneration [62,63]. These examples of cell plasticity have been correlated with the action of specific miRNAs.
The number and variety of targets modulated by the action of one single miRNA made them suitable tools for direct reprogramming and modulation of complex processes, including cancer development and epimorphic regeneration. Direct reprogramming approaches using miRNAs have been employed to generate somatic cell types such as induced hepatocytes and IPCs from MSCs [28,30]. miRNAs have also been applied for somatic cell reprogramming to the pluripotent state [102].
This use of mature miRNAs represents a step forward for the potential clinical application of stem cells and facilitates the development of new approaches to convert iPSCs or MSCs into somatic cells of clinical relevance. On the other hand, miRNAs play an important role in shaping the fate of CSCs and cancer chemoresistance [46,56] and constitute potential molecular targets for cancer treatment. We predict that the study of miRNAs role in all these reprogramming events will become a more intense area of research to understand and potentially modulate cell reprogramming in adult organisms.
Footnotes
Acknowledgments
The authors wish to thank Alejandro Morales-Tapia for critically reviewing this article and Daniel Fuentes for his help in downloading references of difficult access. Work at A.C.-R. and L.B. laboratories has been supported by the Swedish Research Link Program (grant no. 014-9040-114152-32).
Author Disclosure Statement
No competing financial interests exist.