
Abstract
The specification of pluripotent stem cells into the bone-forming osteoblasts has been explored in a number of studies. However, the current body of literature has yet to adequately address the role of Wnt glycoproteins in the differentiation of pluripotent stem cells along the osteogenic lineage. During mouse embryonic stem cell (ESC) in vitro osteogenesis, the noncanonical WNT5a is expressed early on. Cells either sorted by their positive WNT5a expression or when supplemented with recombinant WNT5a (rWNT5a) during a 2-day window showed significantly enhanced osteogenic yield. Mechanistically, rWNT5a supplementation upregulated protein kinase C (PKC), calcium/calmodulin-dependent kinase II (CamKII) and c-Jun N-terminal kinase (JNK) activity while antagonizing the key effector of canonical Wnt signaling: β-catenin. Conversely, when recombinant WNT3a (rWNT3a) or other positive regulators of β-catenin were employed during this same time window there was a decrease in osteogenic marker expression. However, if rWNT3a was supplemented during a time window following rWNT5a treatment, osteogenic differentiation was enhanced both in murine and human ESCs. Elucidating the role of these WNT ligands in directing the early stages of osteogenesis has the potential to considerably improve tissue engineering protocols and applications for regenerative medicine.
Introduction
W
In the developing embryo, cells continually rely on the Wnt signaling network to regulate diverse lineage decisions [6,7]. For example, the importance of β-catenin has been clearly demonstrated in a number of studies examining the maturation of osteoblasts within long bones [8 –10], however, the same studies have also suggested that early osteoblast differentiation in the limbs may occur independently of β-catenin. In these early developmental stages, the noncanonical WNT5a is indeed the only known mouse Wnt protein to be expressed in limb mesenchyme [11,12] and Wnt5a−/− embryos are characterized by significantly shortened limbs and missing distal digits [13]. Additionally, many bones of the craniofacial region develop from a neural crest (NC) origin and proceed through different intermediate stages. However, many of the same signaling cascades are shared in the bones from these different embryonic origins. For instance, the induction and specification of NC cells in mammals is regulated by canonical Wnt signaling [14 –16]. Recent studies have also begun to unravel a dependence of NC delamination and migration on noncanonical Wnt signaling [17].
Given the importance of Wnt signaling in cell and tissue fate determination in vivo, it is not surprising that β-catenin affects osteogenesis of stem cells in vitro. For example, β-catenin supports gastrulation-like events at early stages of in vitro differentiation of pluripotent embryonic stem cells (ESCs) [18,19], which in turn aids the subsequent specification of mesenchymal cells with osteoprogenitor character [20].
Although the involvement of Wnt proteins and β-catenin has been described during early ESC specification, it is still unclear how WNTs contribute to the specification of osteoprogenitors during subsequent stages and whether these stages depend on noncanonical WNTs, as is suggested in vivo. Current examinations of the WNT signaling network on osteogenic differentiation in vitro has been primarily performed on multipotent cells [21,22], and therefore overlook key regulatory events that occur during early osteoprogenitor specification. Since ESCs are ultimately capable of differentiating into mature matrix-mineralizing osteoblasts from pluripotent progenitors [23,24] they offer a greater opportunity for understanding each stage of osteogenic differentiation. A potent enhancer of osteogenic yield is 1α,25(OH)2 vitamin D3 (VD3) [24]. Previous studies in our lab have shown that VD3 supplementation significantly enhances calcification and bone marker gene expression in ESC cultures and identified a critical time dependency of its pro-osteogenic effect [25,26]. As such, expression of osteocalcin (Ocn) is 2.7-fold higher when VD3 induction is initiated on d5 of differentiation compared with d3 [24].
Previous studies in our lab have also utilized global mRNA profiling to identify signaling cascades with the potential to regulate osteogenic commitment from ESCs [26]. Multiple signaling pathways, such as those triggered by bone morphogenetic protein, nitric oxide, and retinoic acid, were identified and experimentally exploited to enhance osteogenic yield [20,26,27]. In that screen, Wnt5a was identified among three Wnt ligands with differential expression as the only one whose mRNA expression was immediately upregulated upon osteogenic induction. Given the aforementioned activity of WNT5a in both the limb mesenchyme and migrating NC cells in the specification of osteoprogenitors in vivo, we sought to explore these putative relationships during osteogenic differentiation of ESCs. Ultimately, the knowledge gained from this study may be applied toward numerous questions facing cell and developmental biology.
Materials and Methods
Stem cell culture
Murine D3 ESCs (American Type Culture Collection) were expanded and differentiated along an osteogenic lineage as previously described [24]. Briefly, cells were routinely grown in monolayer cultures containing Dulbecco's modified Eagle's medium (DMEM; Gibco) supplemented with 4.5 g/L
H9 human embryonic stem cells (hESCs) were from WiCell and were cultured in Matrigel precoated plates with mTeSR (Stem Cell Technologies). Confluent cells were expanded and passaged (1:3–1:6) with accutase. Medium was changed every 2 days. Differentiation was initiated from overgrowing cultures in CDM [28] and osteogenic supplements added on d5 of differentiation. Human bone marrow-derived mesenchymal stem cells (hMSCs; Extem Biosciences) were from a 26-year-old Caucasian male (Lot No. 60515) and cultured on uncoated plastic in media comprised of 15% FBS, 50 U/mL penicillin, 50 μg/mL streptomycin, and 10 μg/mL basic fibroblast growth factor (bFGF) in DMEM/F-12 Nutrient Mixture. To induce differentiation, hMSCs were allowed to grow to 70% confluency and then switched to the osteogenic medium described above.
O9-1 murine NC stem cells were a kind gift from Dr. Robert Maxson (University of Southern California) and were cultured on Matrigel-coated plates in a medium comprised of 15% FBS, 0.1 mM nonessential amino acids, 1 mM sodium pyruvate, 55 μM β-mercaptoethanol, 100 U/mL penicillin, 100 μg/mL streptomycin, and 2 mM
Genetically modified ESC lines
A Wnt5a-GFP reporter ESC line was generated by transfecting an ApaL1 linearized and gel purified Wnt5a-eGFP reporter plasmid into the D3 ESCs, which uses the pEGFP-1 backbone from Clontech to drive GFP expression from the murine Wnt5a promoter (generously provided by Dr. D. Rancourt, University of Calgary). Transfection of ESCs and identification of clones were performed as previously described [25]. LEF/TCF-GFP ESCs were donated by Dr. I. Weissmann (Stanford University) and both Wnt5a−/− and Wnt5a+/− ESCs were kindly provided by Dr. Kang-Yell Choi (Yonsei University, Korea).
Analytical flow cytometry and FACS sorting
For analytical flow cytometry, cells were harvested at the days indicated. Single cell suspensions were generated with trypsin and cell numbers and viabilities were determined using a CASY cell counter. Two million cells per sample group were run through a Cytomics™ FC500 instrument (Beckman Coulter) against a control sample from time-corresponding wild-type cells or IgG controls. Experiments were repeated three times and individual biological replicates run with three technical replicates. All samples were run in appropriate scatter gates employed during analysis with the CXP software (Beckman Coulter) to avoid cellular debris and cell aggregates. For sorting experiments, cultures were harvested on day 5 and 7. Cells were resuspended in 1% FCS in phosphate-buffered saline (PBS) and sorted into six-well culture plates containing complete osteogenic medium using a BD FACSaria. Cultures were phenotypically analyzed on d25 with the GelCount™ system from Oxford Optronics. All flow experiments were performed at the University of Leipzig Flow Cytometry Core Facility.
RNA isolation, semi-quantitative and real-time quantitative polymerase chain reaction
Total RNA was isolated and transcribed into cDNA as described [24]. Quantitative polymerase chain reaction (PCR) was performed essentially as described using a SYBR green PCR master mix (Abgene) [30]. A melting curve was obtained for each PCR product after each run to confirm the presence of a single amplicon. Primers were designed with primer3 (
Biochemical determinations
Cells were rinsed in PBS and lysed in RIPA buffer [150 mM NaCl, 10 mM Tris, pH 7.2, 0.1% sodium dodecyl sulfate (SDS), 1% Triton X-100, 1% deoxycholate, and 5 mM EDTA] containing protease inhibitors (P-8340; Sigma-Aldrich). After gentle rocking for 30 min, lysates were collected, centrifuged, and an aliquot of the supernatant mixed with DC protein assay reagents from Bio-Rad. After 15 min incubation at room temperature, absorbance was read in a Safire II (Tecan) at 750 nm. Protein quantities in samples were taken from a bovine serum albumin standard curve. Alk Phos enzyme activity and calcium measurements were performed as described [31,32].
Protein expression analysis
Cells were harvested in 1× PBS (pH 7.4) containing 1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS, protease inhibitor cocktail (1:100; Sigma-Aldrich), sodium fluoride (10 mM; Sigma-Aldrich), and sodium vanadate (1 mM; Sigma-Aldrich). Protein concentrations were measured as described above and boiled samples were run on a 12% SDS–polyacrylamide gel. Blotted proteins were probed with a mouse monoclonal anti-CATNB (Invitrogen), goat polyclonal anti-GFP (Abcam), a mouse monoclonal anti-β-actin (Santa Cruz), mouse monoclonal anti-CamKIIα (Abcam), rabbit polyclonal anti-phospho-CamKIIα (T286), anti-JNK1, anti-phospho-JNK1 (T183, Y185), anti-JNK1/2, or anti-phospho JNK1/2 (T138, Y185) antibody (all Abcam) followed by horseradish peroxide-conjugated polyclonal anti-mouse (Santa Cruz), anti-goat (Invitrogen), or anti rabbit (Cell Signaling) IgG. Detection was performed with the ECL western blotting detection kit (Amersham). Blots were then incubated with stripping buffer (100 mM 2-mercaptoethanol, 2% SDS, and 62.5 mM Tris-HCl, pH 6.7) at 50°C for 1 h with occasional agitation and reprobed.
Immunocytochemistry
Cultures were fixed with 2% PFA in PBS for 20 min at 4°C. Unspecific binding was blocked with 10% FCS in PBS for 30 min at room temperature. β-Catenin was detected with either mouse anti-catenin beta-1 (Zymed) at 10 μg/mL, goat anti-WNT5a (R&D Systems) at 1:80, goat anti-TBRA (R&D Systems) at 1:80, mouse anti-PDGFRα (Abcam) at 200 μg/mL and rabbit anti-p75 (Abcam) at 1:250. Cells were overlaid with the first antibody in PBS, 10% FCS and incubated at 4°C over night. The secondary antibodies, a goat anti mouse IgG (H+L) Alexa Fluor 546-conjugated (Invitrogen), goat anti-rabbit Alexa Fluor 546 (Invitrogen), and donkey anti-goat Alexa Fluor 488 (Invitrogen) were diluted to 1:2,000 with 5% FCS in PBS and applied to the cells for 2 h at 4°C. Cells were imaged with a Leica fluorescence microscope.
Alizarin Red S staining
Cells were washed with PBS (without Ca2+/Mg2+) three times, fixed with methanol/acetone (7:3) at −20°C for 30 min. Following three rinses in distilled water, cells were stained with 2% (w/v) Alizarin Red S for 10 min at room temperature, and then extensively washed with water and PBS.
Proliferation assay
The proliferative capacity of the cells was determined using BrdU incorporation (BrdU labeling and detection kit III; Roche) according to the manufacturer's instructions. Fifty thousand cells per treatment group were inoculated on differentiation day 5 and treated as indicated. After 3 h, BrdU labeling solution (111 μM BrdU) was added and BrdU incorporation measured after an 18 h incubation period by adding anti-BrdU-POD, Fab fragments, at a final concentration of 200 mU/mL.
PKC activity assay
Cells were harvested in buffer containing 20 mM 3-(N-morpholino)propanesulfonic acid, 50 mM β-glycerophosphate, 50 mM sodium fluoride, 1 mM sodium orthovanadate, 5 mM ethyleneglycotetraacetic acid, 2 mM ethylenediaminetetraacetic acid, 1% NP40, 1 mM dithiothreitol, 1 mM benzamidine, 1 mM phenylmethanesulphonylfluoride, and 10 μg/mL leupeptin and aprotinin. PKC activity was measured using the Enzo Life Sciences PKC kinase activity kit as per the manufacturer's instructions.
Statistical analysis
Data are presented as mean ± standard deviation from three independent biological replicates, containing three technical replicates if not otherwise noted. Representative gel pictures were selected from a set of three. Comparison of two groups was made using Student's t-test for unpaired data. Comparison of more than two groups was conducted using ANOVA. A P-value of above or equal to 0.05 was considered significant.
Results
Wnt5a is expressed upon VD3 induction in ESCs
To explore the role of Wnt5a we first differentiated mouse ESCs following our previously established protocol (Fig. 1A) [24]. With this protocol, mineralized matrix, positive for OCN, appears black in phase contrast microscopy (Fig. 1A). In agreement with our former transcriptional profiling [26], cells treated with our osteogenic inducer VD3, demonstrated a significant increase of Wnt5a mRNA and WNT5a production on day 7 when examining the first 10 days of differentiation (Fig. 1B, C). Together, these data suggest that Wnt5a acted downstream of VD3. Thus, if VD3 was inducing osteogenesis via Wnt5a, we expected to find a similar osteogenic yield by omitting VD3 while supplementing cultures with recombinant Wnt5a (rWNT5a). However, against our hypothesis, rWNT5a treatment, for the duration of the osteogenic induction period (ie, days 5–30), resulted in reduced extracellular matrix (ECM) mineralization compared to VD3 induction (Fig. 1D, E).
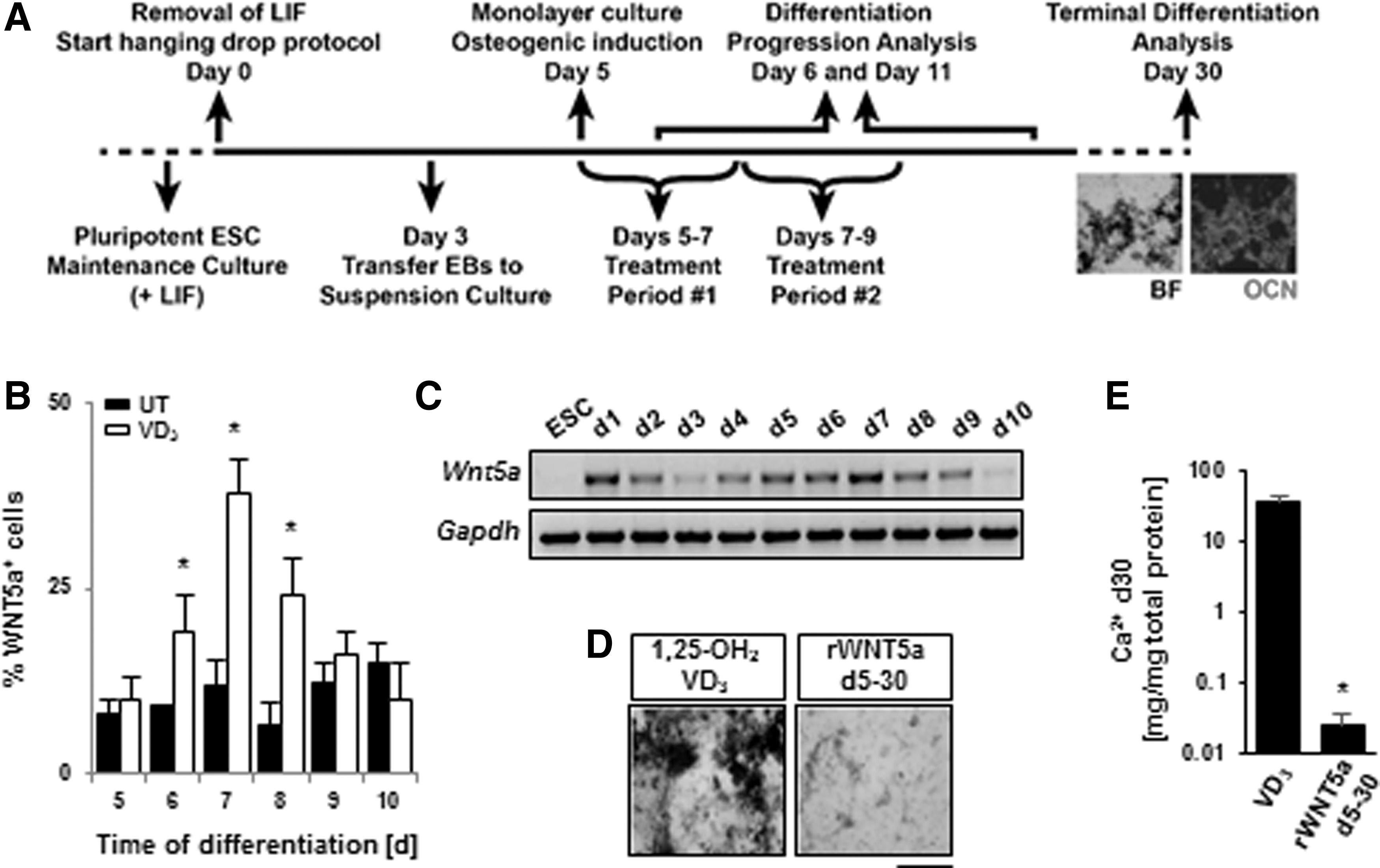
Wnt5a is expressed during early ESC osteogenesis.
Cells expressing Wnt5a show enhanced osteogenic capacity
To further investigate whether endogenous Wnt5a expression marks a population with a greater potential to differentiate into osteoblasts, we utilized an ESC line that expressed GFP under the control of the Wnt5a promoter. GFP detection was found to temporally correlate to Wnt5a mRNA levels (Fig. 2A). GFP+ cells, sorted on day 7, showed enhanced calcification and Alk Phos activity compared to the GFP− fraction when measured at the end of the differentiation period (Fig. 2B–D). Conversely, GFP+ cells sorted on day 5 exhibited reduced calcification when compared to GFP− cells sorted on the same day. In both cases, these differences in mineralization could be detected as early as day 11 (Fig. 2E). Furthermore, the degree of mineralization in day 7 GFP− cells could be rescued if cultures were additionally supplemented with rWNT5a (Fig. 2F). Day 5 GFP− cells could acquire GFP expression by day 7, potentially explaining their increased capacity for osteogenesis (Fig. 2G). The reduced calcification capability in the d5 GFP+ cells may suggest that the cells reporting Wnt5a expression on that day may be a separate lineage that never acquires osteogenic properties. Thus, it appeared that the varying capacity of these GFP reporter cells to differentiate along the osteogenic lineage was dependent on endogenous Wnt5a expression and highly time-specific.
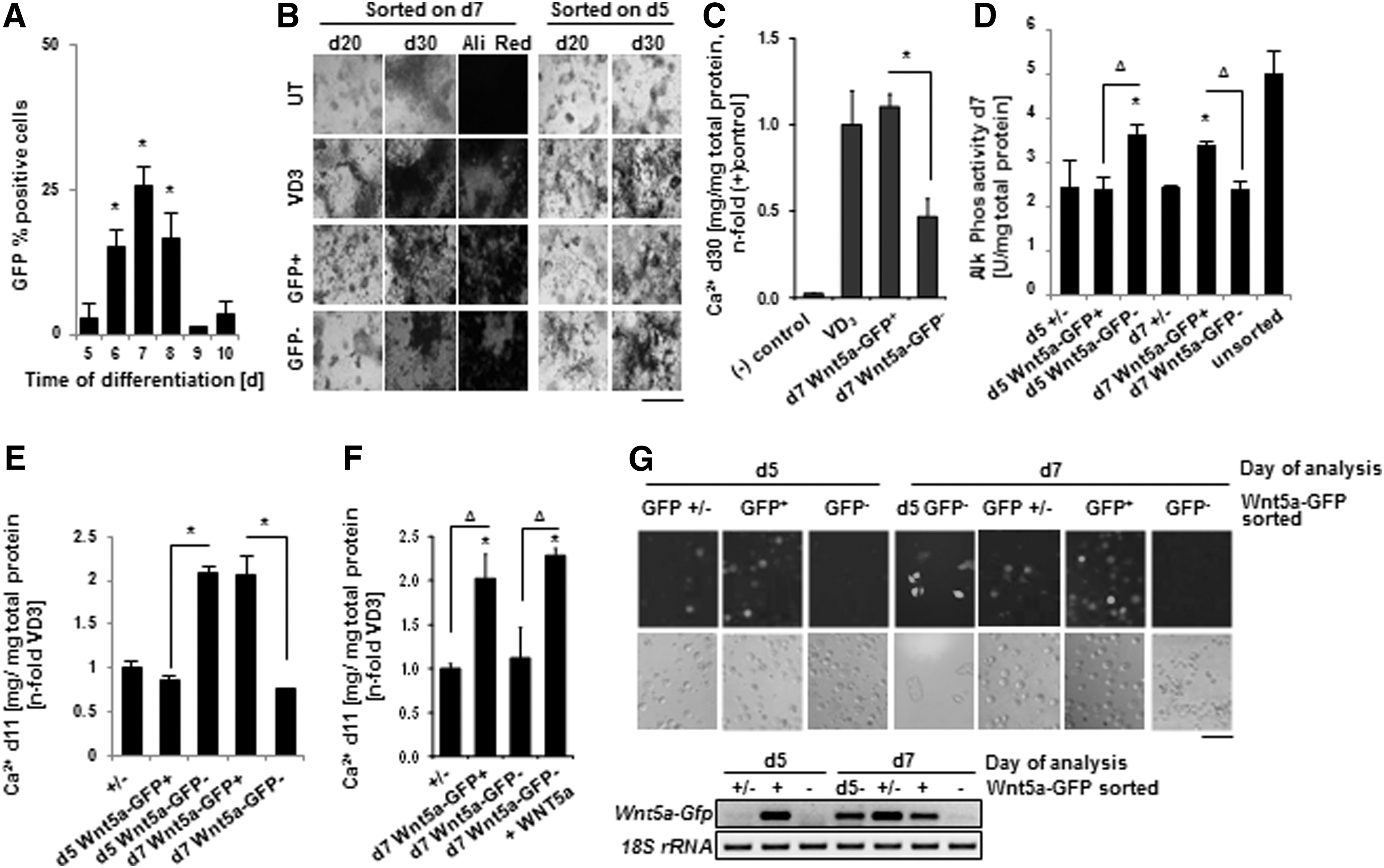
A Wnt5a positive population is enriched for cells with osteogenic potential.
Stage-specific WNT5a supplementation augments mineralization
In light of previous studies, where Wnt5a expression correlated to early stages of osteogenic differentiation [11,12], prolonged rWNT5a treatment in our in vitro system (Fig. 1D, E) may potentially mask its pro-osteogenic effects at earlier stages. To test this hypothesis, ESCs were treated with different concentrations of rWNT5a, with and without VD3, from days 5 to 7 only, which is the time corresponding to the period of highest endogenous mRNA expression following osteogenic induction (compare Fig. 1B). Treatment with rWNT5a without and with VD3 resulted in a significant increase in Ca2+ content as compared to our negative control containing the mineralizing substrates β-glycerophosphate and ascorbic acid (Fig. 3A). This result demonstrated that rWNT5a could functionally mimic the VD3-induction of osteogenesis in regards to calcium deposition when supplemented during this time window. In addition, rWNT5a supplementation was able to rescue the calcification defect found in Wnt5a −/− ESCs when applied between days 5 and 7 of the differentiation (Fig. 3B).

rWNT5a enhances osteogenic differentiation of murine ESCs.
Just 24 h after initial rWNT5a supplementation, the expression of the osteoprogenitor markers alkaline phosphatase (Alp) and osteopontin (Opn) was significantly increased (Fig. 3C), followed by increased Ocn and bone sialoprotein (Bsp) messages at subsequent stages of the differentiation (Fig. 3D). Concurrent to the increase in these osteoblast-specific markers, there was also a significant drop in the chondrocyte-specific Aggrecan (Agg) and collagen type II (Col2a1) expression (Fig. 3D). These results were in contrast to culture treatments in the same time window with agents that act in a canonical Wnt fashion, such as rWNT3a or lithium chloride (LiCl), which demonstrated the opposite pattern of expression.
Wnt5a supplementation increases osteogenic commitment at the expense of proliferation
The rise of bone-specific marker expression following rWNT5a treatment may be attributed to these markers being direct downstream targets of activated signaling cascades, or supplementation may act through more indirect means, such as altering progenitor population specification. Given that Wnt5a has previously been demonstrated to negatively affect cell proliferation [33], BrdU incorporation in differentiating ESCs was measured following 24 h of treatment. Both VD3 and rWNT5a treatment inhibited proliferation, independently and when supplemented simultaneously (Fig. 3E). In conjunction with the upregulation of bone-specific markers, these data suggest a shift from proliferation toward differentiation following treatment with VD3, rWNT5a, or when used simultaneously.
The rWNT5a pro-osteogenic effect likely occurs through the enhancement of mesenchymal or osteoprogenitor specification
The rWNT5a time window of action and Wnt5a expression coincides with a period of differentiation at which Eng and Nt5e, encoding for the mesenchymal markers CD105 and CD73, are expressed (Fig. 3F). In addition, both treatments increased the expression of Snai1 and Snai2 indicating the upregulation of genes typically involved in epithelial to mesenchymal transition (EMT) [34,35]. When specifically examining day 7 we found that both treatment groups demonstrated an increase in mesenchymal marker expression (eg, Nt5e, Ngfr), which was concomitant with both a reduction of adipocyte (eg, Glut4 and Pparg) and chondrocyte markers (eg, Sox9, Col2a1, and Col10a1) (Fig. 3G). On this day cells that strongly stained for WNT5a protein co-stained for the mesenchymal marker PDGRFα using specific antibodies (Fig. 3H). Collectively, these results suggest that Wnt5a may participate in the transition of cells toward a mesenchymal fate, which then follow a primarily osteoprogenitor fate at the expense of chondro- and adipogenic progression.
Both VD3 and Wnt5a decrease nuclear levels of β-catenin
Despite the fact that canonical Wnts and β-catenin are typically thought to determine the fate of osteogenic cells [8,36], it appeared that in ESCs Wnt5a, a noncanonical Wnt, contributed to the specification of mesenchymal/osteoprogenitor cells. While noncanonical Wnts, have been classically viewed to act independently of canonical Wnt signaling, a growing body of evidence has uncovered numerous instances of cross-talk between these pathways [37,38]. Reports have also indicated that cells expressing a profile of specific frizzled receptors, co-receptors, and cytoplasmic regulators of β-catenin showed activation of canonical Wnt signaling cascades in response to Wnt5a [39 –41]. Thus, we next sought to determine the relationship between Wnt5a and β-catenin during the time period in which this noncanonical Wnt has a pro-osteogenic effect.
First, we examined the distribution of the β-catenin during days 5–7 of our osteogenic induction protocol (Fig. 4A). Here, we found that β-catenin levels in whole cell lysates remained relatively consistent during these 3 days. However, β-catenin levels in the nucleus were noticeably reduced on days 6 and 7. Next, by employing a LEF/TCF-GFP reporter line as a functional readout of β-catenin transcriptional activity, we detected a decrease in Gfp mRNA expression following VD3, rWNT5a, and rWNT5a+VD3 treatment, which corresponded to a decrease in Ctnnb1 mRNA expression (Fig. 4B) and a decrease in nuclear β-catenin levels (Fig. 4C). We also noted a plasma membrane localization of β-catenin following 24 h of VD3, rWNT5a, and rWNT5a+VD3 treatment (Fig. 4D). When examining supplementation with rWNT3a or the β-catenin agonist LiCl, we found that treatment caused an increase in GFP expression (Fig. 4E). Thus, it appears that modulation of β-catenin nuclear localization, at this time point, could yield predicable outcomes of osteogenic output.
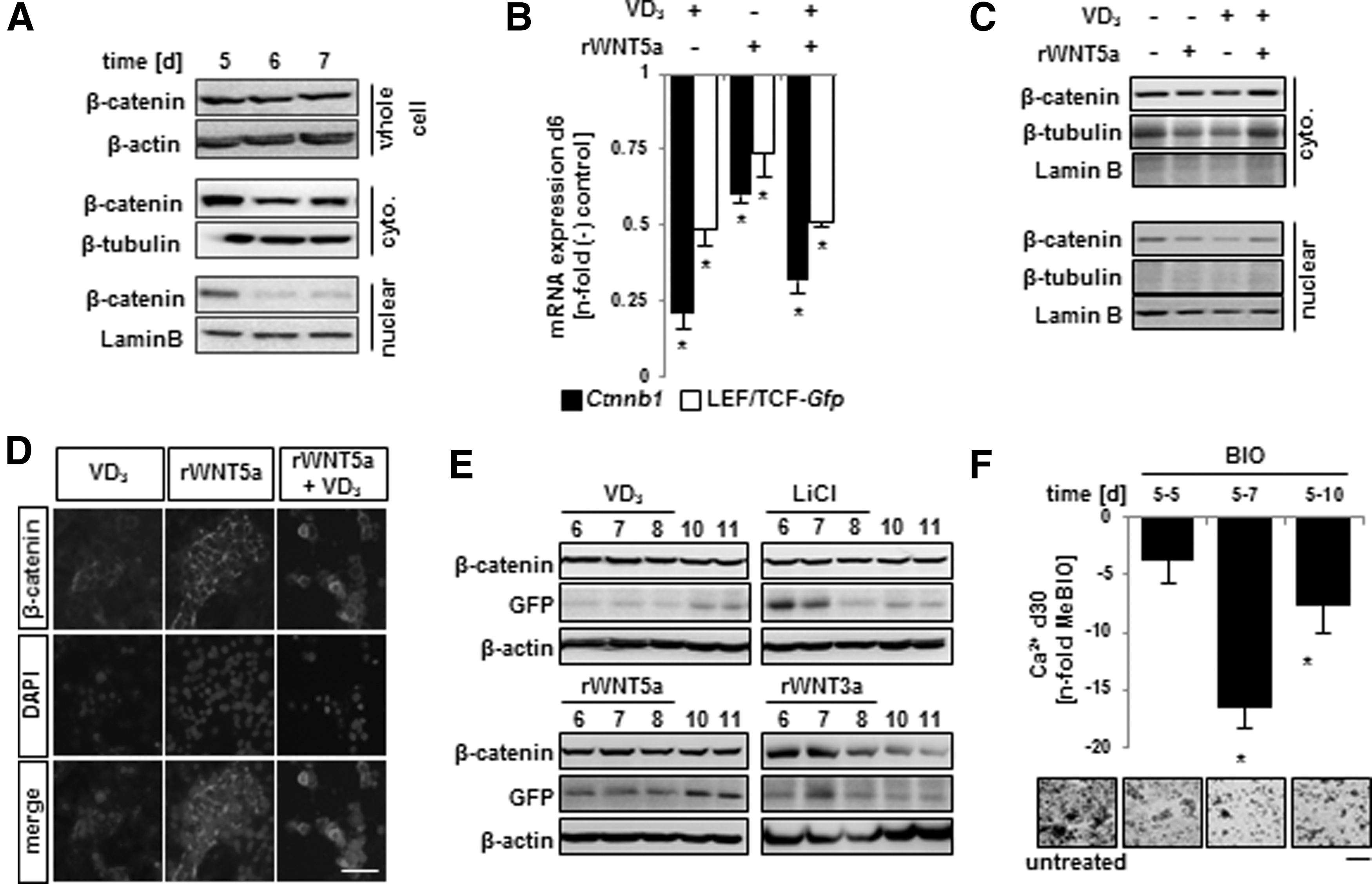
The time window and action of Wnt5a coincides with low levels of nuclear β-catenin.
We tested the role of nuclear β-catenin on directing osteogenic output by utilizing 6-bromoindirubin-3′-oxime (BIO), which inhibits a direct inhibitor of β-catenin: glycogen synthase kinase-3 (GSK-3). Treatment with BIO dramatically reduced subsequent mineralization of the cultures (Fig. 4F). Additionally, when compared to the day 5–7 treatment group, both shorter and longer BIO treatments were not as effective in reducing mineralization; highlighting the importance of this time window and low β-catenin nuclear levels in directing osteogenic differentiation. Taken together these data support the pro-osteogenic role for both VD3 and rWNT5a through the downregulation of nuclear β-catenin and its correlated reduction in transcriptional activity.
Inhibition of downstream effectors of Wnt5a suppress osteogenesis
To address how Wnt5a signaling specifically affected β-catenin transcriptional activity, we examined three known downstream effectors of Wnt5a signaling: Ca2+/calmodulin-dependent protein kinase II alpha (CAMKIIα), JNK, and PKC. All three of these downstream effectors were detectable during the window of pro-osteogenic action of Wnt5a (Fig. 5A). Following osteogenic induction or when cultures were additionally supplemented with rWnt5a, an upregulation of the alpha isoform of PKC was noted, and to a lesser extent the beta isoform (Fig. 5B). In a PKC activity assay, rWNT5a treatment produced a significant increase of activity above negative controls following analysis at 15, 45 min, and 12 h post-treatment (Fig. 5C). However, a statistical difference in PKC activity was only detected at the 12 h period following VD3 supplementation. Thus, although both treatment types achieve an increase in PKC activity, they appear to do so with different kinetics. Consequently, the combinatorial treatment of rWNT5a+VD3 achieved a significant upregulation of PKC activity at all time points examined. We also detected a clear upregulation of CamKIIα in addition to Jnk1 and Jnk2 expression following both VD3 and rWNT5a treatment (Fig. 5B). In addition, supplementation with both of these osteogenic inducers resulted in an increase in active CAMKIIα and JNK1/2 as determined by the presence of activating phosphoryl groups after 45 min, although JNK1 was only minimally changed (Fig 5D). However, in contrast to PKC activation, this effect did not persist when examined at 12 h post-treatment (data not shown).
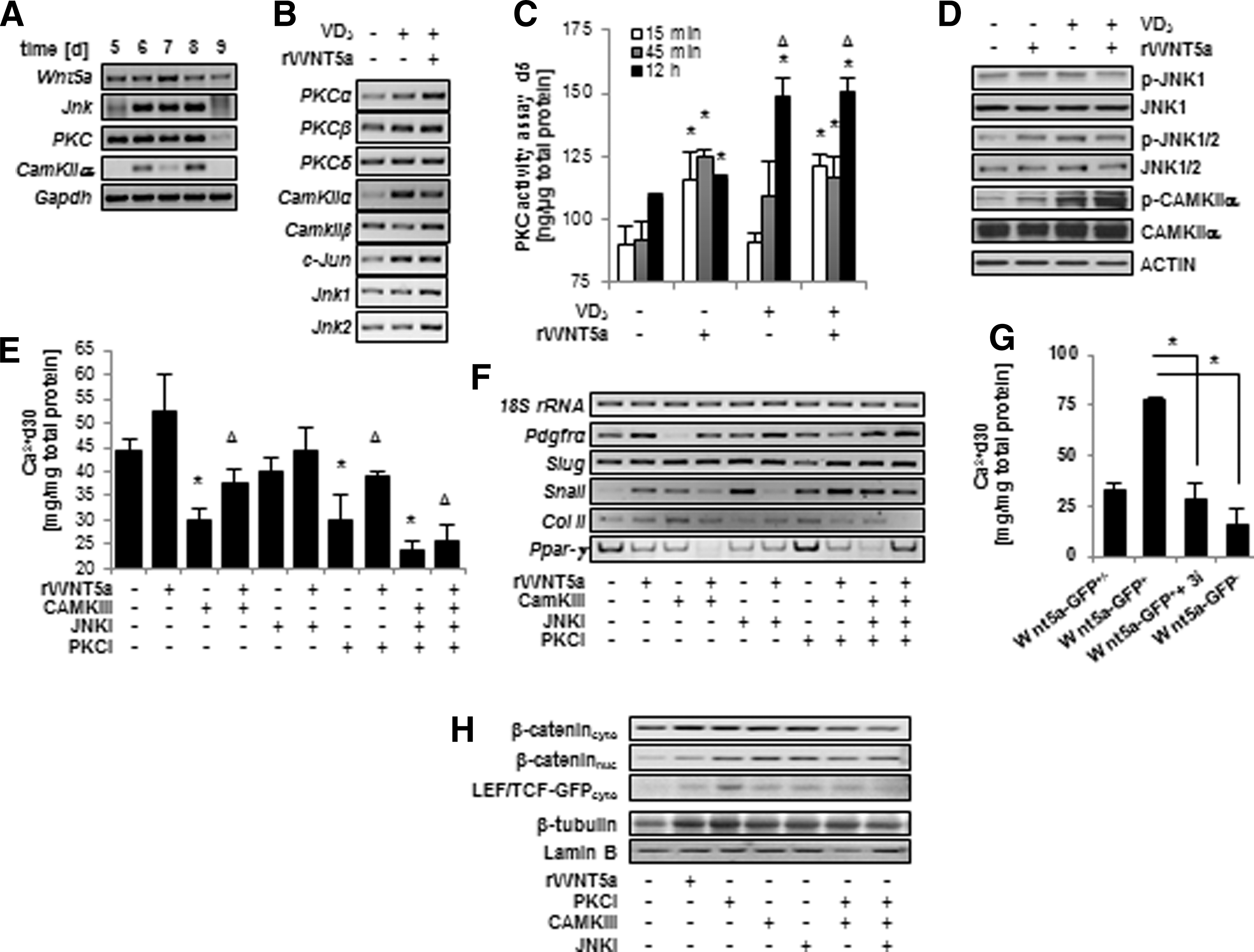
Wnt5a signals through CAMKII, JNK, and PKC to enhance osteogenesis.
If these downstream effectors mediated the effect of rWNT5a on osteogenic differentiation and β-catenin localization, then their inhibition should abrogate these effects. Initial morphological examination of our cultures revealed that chemical inhibition of all three effectors reduced mineralized ECM. Individually, each inhibitor treatment during days 5–7 significantly reduced calcium deposition, with PKC and CAMKIIα inhibition resulting in the strongest impact on ECM calcification (Fig. 5E). We also detected a downregulation of Snai2 following PKC inhibition and a dampened Pdgrfα and Snai1 expression following CAMKII inhibition (Fig. 5F). At the same time, treatment with both inhibitors correlated to an upregulation of adipogenic and chondrogenic genes. With Pdgrfα being associated with mesenchymal progenitors and Snai1 and Snai2 being involved in EMTs [34,35], these immediate changes suggest that PKC and CAMKII could regulate the migration and mesenchymal potential of differentiating ESCs. The described changes in early mesenchymal and migratory gene expression in response to the inhibitors indeed correlated with an inhibition of calcification in the reported Wnt5a+ population (Fig. 5G). Furthermore, we found an upregulation of β-catenin in the nuclear fraction of cells treated with each inhibitor; further supporting the role of WNT5a and its effectors as negative modulators of nuclear β-catenin and its subsequent transcriptional activity (Fig. 5H).
Wnt3a supplementation at later stages is pro-osteogenic
Since prolonged rWnt5a exposure beyond days 5–7 inhibited osteogenesis (Fig. 1D), and the GSK-3 inhibitor, BIO, had the strongest inhibitory effect during this same, narrow time period (Fig. 4E), it appeared to us that regulation of β-catenin is a tightly coordinated process, and that restoring its stabilization after day 7 was maybe critical for proceeding through the later stages of differentiation. Therefore, we sought to augment β-catenin stabilization using the canonical Wnt signaling molecule WNT3a, and LiCl at a stage directly following the period of Wnt5a action.
Following VD3-mediated induction of osteogenesis, we detected an upregulation of Wnt3a mRNA expression starting later than the previously detected upregulation of Wnt5a mRNA (Fig. 6A). Thus, we chose to follow the endogenous mRNA expression and treat cultures with recombinant Wnt3a (rWNT3a) at 50 ng/mL on days 7–9. Supplementation with rWNT3a during this period significantly increased calcium deposition and Alk Phos enzymatic activity (Fig. 6B). Since the increase of endogenous Wnt3a expression on day 7 corresponded with an increase in nuclear β-catenin protein (Fig. 6C), we presumed treatments that stabilize β-catenin during this period would also increase osteogenic differentiation. A 24-h treatment with either rWNT3a or LiCl resulted in an increase of Gfp expression in the LEF/TCF reporter line (Fig. 6D).
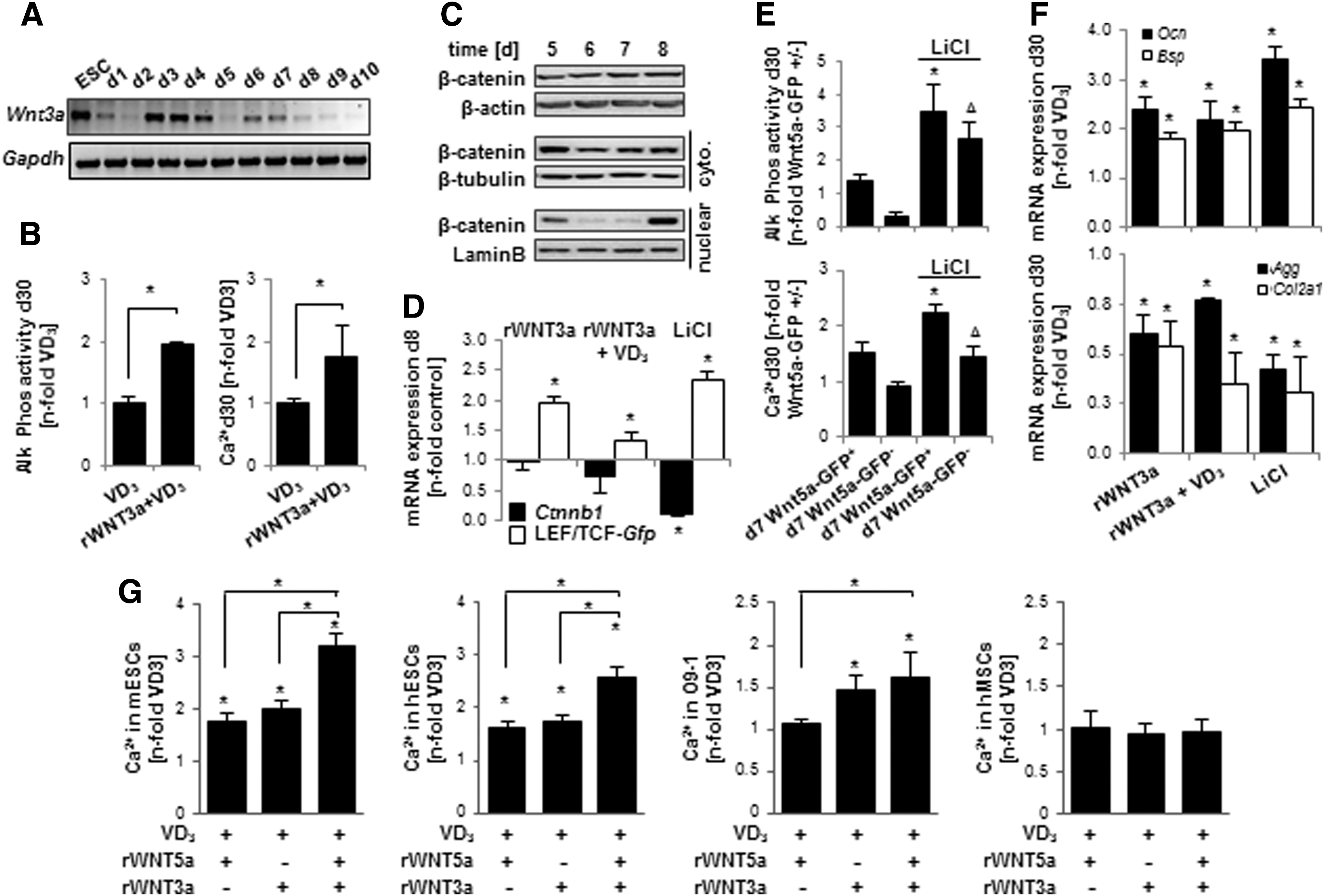
Recombinant Wnt3a enhances osteogenic differentiation of ESCs when applied during a period following rWNT5a treatment.
We also tested this hypothesis by treating sorted Wnt5a-GFP ESCs with LiCl. LiCl treatment increased Ca2+ deposition and Alk Phos activity in both GFP+ and GFP− populations, when supplied from days 7 to 9 (Fig. 6E). Thus, while the pro-osteogenic effect of LiCl treatment was not dependent on previous Wnt5a expression, the greatest osteogenic yield arose from cells with Wnt5a reporter activity on day 7 that were then additionally treated with LiCl until day 9. Moreover, rWNT3a or LiCl treatment from days 7 to 9 enhanced the expression of Ocn and Bsp, and decreased the expression of Agg and Col2a1, when measured on day 30 (Fig. 6F). These data are in direct contrast to the marker expression pattern seen when cells were treated with rWNT3a or LiCl from days 5 to 7 (Fig. 3D).
rWnt5a and rWnt3a act sequentially in ESCs, but not in NC or mesenchymal stem cells
Together, the results presented above indicated a correspondence of increased osteogenic output with increased nuclear β-catenin activity during differentiation days 7–9, which followed a WNT5a driven period of low nuclear β-catenin activity during days 5–7. Treatment with both recombinant Wnt proteins in a sequential manner indeed proved superior to either treatment alone (Fig. 6G), both in murine ESC and hESCs. However, this synergistic effect was not noted in mouse NC stem cells, in which only the rWNT3a treatment led to enhanced calcification. In human mesenchymal stem cells, neither of the recombinant WNT supplementations (each at a 50 ng/mL concentration) increased calcification over VD3. In summary, these data suggest that the pro-osteogenic effects of both rWNT5a and rWNT3a are highly dependent on the differentiation stage of a cell.
Discussion
The importance of Wnt signaling and β-catenin in relation to in vivo and in vitro osteogenesis has been explored in a number of studies; however, the current body of literature has yet to fully address the role of this signaling network on osteogenic differentiation of ESCs. Utilizing an in vitro pluripotent stem cell model to study differentiation progression, we examined the role of the noncanonical WNT5a at earlier stages of differentiation than what can be achieved when employing multipotent MSCs. Here, we provide evidence that not only does endogenous expression of Wnt5a mark a population of cells on day 7 with higher osteogenic capacity, but that supplementing cultures with rWNT5a can enhance osteogenesis when applied from days 5 to 7.
Initial examination revealed that Wnt5a mRNA and WNT5a protein is upregulated following osteogenic induction with VD3. Sorting cells based on Wnt5a reporter expression revealed a clear difference in osteogenic potential in terms of Alk Phos activity and calcium deposition in the ECM. Additionally, Wnt5a−/− cells exhibit a defect in osteogenic differentiation following VD3 induction, which could be rescued by rWNT5a supplementation. These results suggest that the increase of WNT5a expression is directly contributing to osteogenic differentiation, and it is not just a consequence of VD3 treatment. Although there have been some studies demonstrating a pro-osteogenic effect of Wnt5a on naïve MSCs or juvenile osteoblasts [21,22], to our knowledge, this effect has not been reported during the early stages of ESC differentiation. In fact, our limited experiments with human MSCs suggest that the same concentration of rWNT5a has no supportive effect, which may suggest that WNT5a supplementation in ESC cultures targets a population of cells that has not yet committed to a mesenchymal state.
To explore the role of WNT5a in supporting ESC osteogenesis we began treating cultures with rWNT5a. Here, we report that rWNT5a could serve as an osteogenic inducer in the absence of VD3 when it is applied during days 5–7 of the differentiation. Longer supplementation periods did not have the same pro-osteogenic effect; a result that parallels other studies demonstrating the effect of rWNT supplementation being highly contingent on the specific stage of differentiation [21,22,42]. We also found that rWNT5a treatment elicited a decrease in LEF/TCF-GFP reporter activity and that pro-osteogenic treatments, including VD3, also were shown to reduce reported β-catenin transcriptional activity. Reviewing our previous examination of genome-wide transcriptional changes following osteogenic induction [26], some of the most strongly downregulated transcripts are known targets of WNT/β-catenin signaling (eg, cyclin D2, TDGF1, and plasminogen activator inhibitor-1). Furthermore, treatments that stabilize β-cat during this narrow time period, such as rWNT3a or LiCl supplementation, resulted in reduced osteogenic marker expression. Therefore, a strong argument can be made that the concurrent downregulation of WNT/β-catenin signaling during the initial stages of osteogenic induction facilitates differentiation along this lineage. Interestingly, we found that on days 7–9 upregulation of WNT/β-catenin signaling could increase osteogenic yield in ESCs. Thus, we have shown that noncanonical and canonical Wnts support early osteogenesis from ESCs in a sequential and opposing manner.
We also forward the hypothesis that Wnt5a may—at least partially—contribute to the specification of cells to adopt a mesenchymal-osteoprogenitor fate based on the alterations in mRNA expression and cellular identity upon rWNT5a supplementation. For example, the decrease in expression of the adipocyte-specific transcription factor peroxisome proliferator-activated receptor-gamma (PPARγ) in rWNT5a-treated cells goes hand in hand with the reported suppression of PPARγ and induction of Runx2 expression in the ST2 mesenchymal precursor cell line by Wnt5a [43]. Since adipocytes and osteoblasts arise from the same mesenchymal precursors, it is plausible that rWNT5a treatment promotes osteogenesis at the expense of adipogenesis through the regulation of lineage-specific transcription factors.
On the other hand, upon rWNT5a treatment and in Wnt5a:GFP+ cells, we found altered Snai1 and Snai2 expression, which are typically associated with EMTs, which are required for NC cells to delaminate, primarily in the cranial NC [34,44]. In a multitude of other cells, normal and cancerous, Wnt5a contributes to the regulation of these EMT events [45]. Given this link among Wnt5a, EMT, and NC delamination, the altered expression of Snai1 and Snai2 could be indicative of Wnt5a regulating NC cell migration during ESC osteogenesis. Although the migratory behavior of Wnt5a+ cells was not directly examined here, irregular distribution of calcified regions in Wnt5a− cells and β-catenin redistribution to the plasma membrane would underscore the plausibility of this explanation. Of note, osteogenic differentiation from NC progenitors occurs directly from mesenchyme without chondrogenic intermediates as seen in mesoderm-derived bone tissue. Thus, reduced chondrocytic-specific collagen expression seen in rWNT5a-treated cultures may be the result of an NC-like shift in progenitor specification. While further studies are needed to define these progenitors with higher osteogenic capacity, the fact that only rWNT3a is pro-osteogenic in NC cells, while rWNT5a had no effect also contribute to such a conclusion. The differential response noted between the different cells could possibly indicate that rWNT5a participates in the commitment of a NC population during ESC differentiations and that rWNT3a then supports differentiation of these NC cells into a mesenchymal-osteoprogenitor population. Indeed, canonical WNTs have been reported to participate in the differentiation of NC cells [46,47].
This hypothesis seems fitting given that the timeline of events during ESC osteogenesis coincides with the suggested sequence of events during embryogenesis and in vivo NC induction. For instance, Basch et al. have put forth the idea that in vivo NC induction may occur simultaneously with gastrulation, thereby making it a very early embryonic event [48]. During ESC differentiation, gastrulation-like events as indicated by presence of TBRA have been described between day 2.5 and 5 after differentiation induction [20,49], right before the time window of our reported action of Wnt5a. By extension this would suggest that NC induction in ESCs could occur concomitantly with the first upregulation of Tbra expression, but right before specification of the Wnt5a+ progenitors. In fact, we have previously shown that activation of canonical Wnt signaling at a time of gastrulation in ESCs can enhance the osteogenic output [20]. We have attributed this effect to the upregulation of Tbra, and hence a β-catenin-dependency on mesodermal osteogenesis. In the light of our newer findings, these past results may also suggest a dependency of NC induction by β-catenin, which has been put forth for avian and amphibian embryos, but is still debated in mammalian systems [50].
Since Wnt5a may signal through canonical cascades, depending on the receptor make-up of the cell [39 –41], it is therefore possible that the pro-osteogenic role of Wnt5a was mediated by β-catenin. However, in the present work, multiple lines of evidence support the negative regulation of β-catenin by Wnt5a including the following: increased expression of canonical Wnt antagonists following rWNT5a treatment, a redirection of β-catenin from the nucleus to the plasma membrane, and attenuated activity of a LEF/TCF-GFP reporter. Ishitani et al. found that suppression of canonical Wnt signaling was achieved by WNT5a through CAMKII activation [51], a Wnt5a effector whose activation was also found here. Additionally, we found activation of JNK and PKC upon rWNT5a, which was also found in cells supplemented with VD3. JNK, specifically, has been implicated in the migration of cranial NC cells [52] providing another link between Wnt5a and VD3 and its potential role in NC migration. Although VD3 has been classically characterized to act through the vitamin D receptor (VDR) [53], VDR appears to contribute to bone formation postnatally [54]. Instead, the membrane-associated protein PDIA3 (protein disulfide isomerase A3) has been suggested to promote early osteogenesis through rapid activation of PKC [55,56]. Therefore, PKC may be a common node for which the osteogenic induction by VD3 and Wnt5a is mediated.
In summary, modulation of β-catenin by WNT5a through its effectors can enhance in vitro osteogenic differentiation of ESCs, potentially through a mesenchymal-osteoprogenitor stage. However, the narrow time window through which both WNT5a and WNT3a act to support osteogenesis and their differential effects in diverse cell lines that represent different stages of commitment highlights the need for investigators to pay close attention to the precise activation of these signaling cascades to improve differentiation methodologies and to gain insight into developmental processes.
Footnotes
Acknowledgments
Parts of this work were supported with funds from the German Federal Ministry of Education and Research (BMBF grant no. 0313453) and start-up funds from the University of California Riverside to N.I.z.N. The authors thank Beatrice Kuske, Sylvia Taube, and Dr. Vuk Savkovic and the technical staff at the Core Unit Fluoreszenz-Technologien of the Interdisciplinary Center for Clinical Research Leipzig for technical assistance. We are furthermore grateful to Dr. Derrick E. Rancourt (University of Calgary, Canada), Dr. Irving Weissman (Stanford University, USA), and Dr. Kang-Yell Choi (Yonsei University, Korea) for generously providing the Wnt5a-GFP plasmid, LEF/TCF-GFP ESC line, and Wnt5a knockout and heterozygote ESC lines, respectively.
Authors Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.