
Abstract
The innate immunity of embryonic stem cells (ESCs) has recently emerged as an important issue in ESC biology and in ESC-based regenerative medicine. We have recently reported that mouse ESCs (mESCs) do not have a functional type I interferon (IFN)-based antiviral innate immunity. They are deficient in expressing IFN in response to viral infection and have limited ability to respond to IFN. Using fibroblasts (FBs) as a cell model, the current study investigated the development of antiviral mechanisms during in vitro differentiation of mESCs. We demonstrate that mESC-differentiated FBs (mESC-FBs) share extensive similarities with naturally differentiated FBs in morphology, marker expression, and growth pattern, but their development of antiviral mechanisms lags behind. Nonetheless, the antiviral mechanisms are inducible during mESC differentiation as demonstrated by the transition of nuclear factor kappa B (NFκB), a key transcription factor for IFN expression, from its inactive state in mESCs to its active state in mESC-FBs and by increased responses of mESC-FBs to viral stimuli and IFN during their continued in vitro propagation. Together with our previously published study, the current data provide important insights into molecular basis for the deficiency of IFN expression in mESCs and the development of antiviral innate immunity during mESC differentiation.
Introduction
E
The immune system of vertebrates consists of innate and adaptive immunity. Innate immunity responds to a broad range of pathogens in a nonspecific manner and provides the first line of defense through several mechanisms, including inflammation and innate immune response, whereas the adaptive immunity provides defense in a pathogen-specific manner through highly specialized immune cells. The innate immunity is well-developed in most, if not all somatic cells [7]. However, recent studies demonstrated that hESCs do not respond to a wide range of infectious agents [3,8]. Similarly, mESCs are susceptible to the cytopathic effect of bacterial and viral infection, but they do not show immune responses typically seen in differentiated cells [9,10]. We recently reported that mESCs are unable to express type I interferon (IFN) [11,12] and have attenuated responses to these cytokines [13]. Therefore, the IFN system, which is the central part of innate immunity in differentiated somatic cells [14], is not fully functional in ESCs. Together with the similar findings in induced pluripotent stem cells [15] and embryonal carcinoma [16], an underdeveloped antiviral innate immunity represents an intrinsic property of all pluripotent cells (reviewed in Guo et al. [17]).
While we do not yet completely understand the physiological implications of the underdeveloped innate immunity in ESCs, we can speculate from different perspectives. ESCs normally reside in the womb where they have limited exposure to pathogens and are likely protected by the mother's immune system [18]. However, a different conjecture could be made based on the fact that immune and inflammatory responses often have multiple impacts, including various adverse effects on infected cells, such as cell cycle inhibition or cell death [14,19]. These negative effects on ESCs could be detrimental to the organism's development since they are the progenitors for all ensuing tissues. On the other hand, it would be equally disastrous if ESCs do not have an effective antiviral mechanism to prevent viral infection. The recent discovery of RNA interference (RNAi) in mESCs offers a plausible solution to this dilemma [20]. RNAi is a major antiviral mechanism in plants and invertebrates that lack IFN-based innate antiviral immunity. It has been uncertain whether RNAi functions in mammals, where a well-developed IFN system can mount multiple forms of antiviral responses [21,22]. Using mouse models, it has been recently demonstrated that the RNAi mechanism is functional in mESCs, but its efficiency is significantly diminished in differentiated cells [20,23]. An emerging paradigm is that mammals may have adapted different antiviral strategies at different stages of development. By utilizing virus-specific and short-lived siRNA derived from invading viruses, a developing organism may prevent viral infection in ESCs and avoid potential negative effects associated with IFN, while later developed IFN-based antiviral mechanisms in somatic cells may confer powerful antiviral activities at multiple levels (reviewed in Pare and Sullivan [21]).
Based on the lack of effective responses of ESCs to various pathogens, it is apparent that the innate immunity is not “innate” to ESCs and it must be “acquired” by somatic cells during the process of organism development. It is conceivable that the attenuated immune and inflammatory responses in in vitro ESC-derived cells is likely attributed to the lack of the same function in ESCs and the process of their differentiation. This not only raises concerns for their therapeutic application, but also brings up several questions that overlap the basic sciences of ESC biology, developmental biology, and immunology. In this study, we attempted to address the question of whether the commonly used in vitro differentiation methods can generate ESC-derived cells with active antiviral innate immunity. Our findings provide valuable insights into this question and could be instructive for designing strategies that can generate ESC-derived cells with desired level of innate immunity for their use in regenerative medicine.
Materials and Methods
Cell culture
D3 and DBA252 mESCs were maintained in the standard mESC medium as previously described [12]. C3H10T1/2 cells [10T1/2, a line of mouse embryonic fibroblasts (FBs), ATCC] were cultured in DMEM with 10% fetal calf serum (FCS) and 100 U/mL penicillin and 100 μg/mL streptomycin. All cells were maintained at 37°C in a humidified incubator with 5% CO2.
mESC differentiation
Two common methods for in vitro differentiation of mESCs were used in this study. The first one was ESC differentiation through embryoid body (EB) formation as we previously described [24]. Briefly, mESCs (1 × 105 cells/mL) were suspended in a bacterial culture dish where they clumped and formed EBs. After incubation for 24 h, the medium was changed to leukemia inhibitory factor (LIF)-free medium containing 15% FCS to initiate differentiation. After incubation for 5 days, EBs were transferred to gelatin-coated cell culture dishes where the cells within the EBs grew out to form a monolayer. The cells derived from the monolayer, designated as mESC-differentiated cells (mESC-DCs), represent mixed populations of different cell types as we previously characterized [24].
In the second method, mESC differentiation was induced by retinoic acid (RA) according to the published method with some modifications [13]. Briefly, mESCs grown in low density (30%–40% confluence) in a cell culture dish were treated with 1 μM RA and the medium was refreshed three times during a 10-day period of differentiation. The differentiated cells formed a compact monolayer, which was trypsinized and disaggregated. The single cell suspension was reseeded in an uncoated cell culture dish, to which FBs quickly attached within ∼30 min. The floating cells in the medium were removed by changing the medium. This process was repeated two to three times and the resulting cells showed the typical morphology of FBs (designated as passage one, p1) and expressed several cell markers common to FBs as previously characterized [13]. Thus, mESC-differentiated FBs were named as mESC-FBs (ie, D3-FBs and DBA-FBs, differentiated from D3 and DBA mESCs, respectively). These cells were further propagated in DMEM with 10% FCS to different passages. The cells from a specified passage or from several consecutive passages were used for related experiments as described in individual experiments.
Cell proliferation, viability, and cell cycle analysis
Cell proliferation and viability were determined by Toluidine Blue staining as we previously described [25]. The absorbance at 630 nm of stained cells was measured with a BioTek ELx800 Microtiter Plate Reader. The absorbance values, which correlate with the amount of cellular content (proteins/DNA), were used as an indirect measurement of cell number. Cell cycle analysis by flow cytometry was performed after the cells were stained with Propidium Iodide. The cell cycle profiles were generated with the CFlow software as previously described [25].
Cell treatment
The 10T1/2 cells and mESC-FBs were seeded at 70%–80% confluence or otherwise specified. The antiviral responses induced by polyinosinic: polycytidylic acid (polyIC, a synthetic dsRNA used as a viral analog) were performed by transfection of the cells with polyIC (300 ng/mL or otherwise specified) using DharmaFECT reagent (Thermo Scientific) [12]. The cellular responses to type I IFN were determined with recombinant IFNα (IFNα−2, 1 × 108 U/mg; eBioscience) and recombinant IFNβ (5 × 108 U/mg; PeproTech) as previously described [13]. To inhibit the expression or activation of dsRNA-dependent protein kinase (PKR), the cells were transfected with siRNA targeting PKR (Santa Cruz Biotechnology) or treated with a PKR inhibitor (imidazolo-oxindole, C16; Sigma) at the concentration of 1 μM, as described in our published study [12].
Viral stock preparation and cell infection
La Crosse virus (LACV, SM6 v3) and chikungunya virus (CHIKV, LR 2006 OPY1 strain) were propagated in Vero cells (African green monkey kidney cell line, ATCC). Titers of virus stocks were determined in Vero cells by plaque assay as previously described [26]. The cells were infected with LACV and CHIKV at multiplicity of infection of 5 and 1, respectively, or otherwise specified in individual experiments.
Real-time quantitative polymerase chain reaction
Total RNA was extracted using TRI Reagent (Sigma). cDNA was prepared using Moloney Murine Leukemia Virus Reverse Transcriptase (Sigma). Real-time quantitative polymerase chain reaction (RT-qPCR) was performed using SYBR Green Ready Mix (Bio-Rad) on a MX3000P RT-PCR system (Stratagene), as previously described [24]. The mRNA levels from RT-qPCR were calculated using the comparative Ct method [27]. β-actin was used as a calibrator for the calculation of relative mRNA of the tested genes. In the experiments with viruses, 18S rRNA was used as a calibrator due to the degradation of β-actin caused by viral infection. The sequences of the primer sets are as follows (F, forward; R, reverse)
β-actin, F: 5′-CATGTACGTAGCCATCCAGGC-3′ R: 5′-CTCTTTGATGTCACGCACGAT-3′
18s rRNA, F: 5′-GTAACCCGTTGAACCCCATT-3′ R: 5′-CCATCCAATCGGTAGTAGCG-3′
IFNβ, F: 5′-CCCTATGGAGATGACGGAGA-3′ R: 5′-ACCCAGTGCTGGAGAAATTG-3′
RIG-I, F: 5′-ATTCAGGAAGAGCCAGAGTGTC-3′ R: 5′-GTCTTCAATGATGTGCTGCAC-3′
TLR3, F: 5′-CTTGCGTTGCGAAGTGAAGAA-3′ R: 5′-CCAATTGTCTGGAAACACCCC-3′
MDA5, F: 5′-CGATCCGAATGATTGATGCA-3′ R: 5′-AGTTGGTCATTGCAACTGCT-3′
Flow cytometry
Cellular protein analysis by flow cytometry was performed according to our published method [12]. Briefly, treated cells were fixed with 3% paraformaldehyde and incubated with the antibodies against: TLR3 (toll-like receptor 3, BioLegend), IFNβ (BioLegend), RIG-I (retinoic acid–inducible gene I, Santa Cruz Biotechnology), PKR (Santa Cruz Biotechnology). With the exception of anti-TLR3 antibodies, which were preconjugated with PE (phycoerythrin), all other antibodies were detected with secondary antibodies that were either conjugated with FITC (fluorescein isothiocyanate) or rhodamine. The cells were then analyzed with an Accuri C6 flow cytometer (BD Biosciences). The fluorescence intensity, which correlates with the protein level detected with its specific antibody, was determined with the CFlow software [12].
Immunocytochemistry
Immunostaining was performed according to our published method [13]. The cellular location of nuclear factor kappa B (NFκB) was determined with its specific antibodies (Santa Cruz Biotechnology). The cells infected with CHIKV were detected with a monoclonal anti-CHIKV antibody 3585 (Abcam). The cells infected with LACV were detected with monoclonal antibodies against the Gc protein encoded by LACV genome (a gift from Dr. Samantha Soldan, the University of Pennsylvania School of Medicine) [28]. The cells were examined under an LSM 510 laser-scanning confocal microscope (Zeiss).
Statistical analysis
For statistical analysis, data are presented as the mean ± SD derived either from three independent experiments or from a representative experiment performed in triplicate that was performed at least twice with similar results. Statistical analysis was performed using a two-tailed and paired student's t-test. Differences are considered statistically significant when P < 0.05.
Results
mESC-DCs through EB formation have a limited capacity to express IFN
The stem cell state of mESCs is maintained by LIF. The basic principle of in vitro differentiation is that LIF withdrawal from the culture medium triggers spontaneous differentiation while the cell fate and differentiation rate can be influenced by different agents and growth conditions [2]. Undifferentiated mESCs grow in colonies in a cell culture dish. When deprived of cell adhesion and cultured in suspension, mESCs grow in aggregates and form structures known as EBs since they resemble an early embryo in structure and differentiation process [24]. As illustrated in Fig. 1A, mESCs were allowed to differentiate in the form of 5-day-old EBs (5dEBs) followed by further differentiation in a monolayer formed from the outgrowth of EBs (5dEB-5dMo). When differentiated cells in the monolayer were trypsinized and replated into a new culture dish, they attached and formed a new cell monolayer consisting of different cell types (collectively designated as mESC-DCs, Fig. 1A). Since mESCs are unable to express IFNβ in response to synthetic viral RNA analogs and live viral infection [11,12], we examined whether differentiation would change this deficiency. As shown in Fig. 1B, both polyIC (a synthetic dsRNA that has been commonly used as a viral RNA mimic) and LACV induced robust IFNβ expression in 10T1/2 cells (naturally differentiated FBs) as a positive control (>3,000-fold activation), whereas mESCs were basically unresponsive. Although mESC-DCs showed a notable increase of IFNβ expression compared to mESCs in response to polyIC (∼500-fold activation) or LACV infection (∼40-fold activation), such induction is substantially lower than 10T1/2 cells.
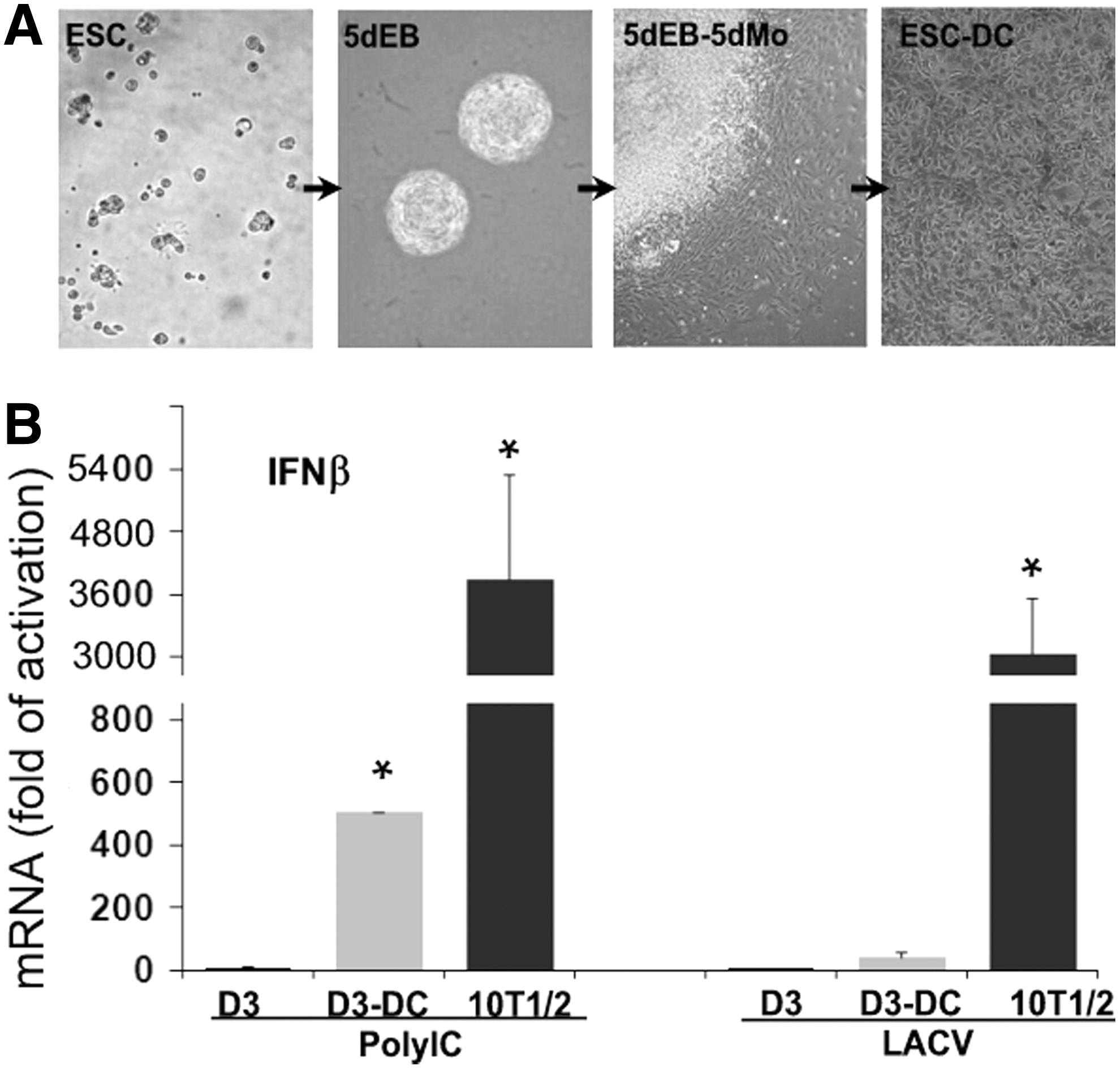
mESC-DCs generated from EB formation express limited IFNβ in response to polyIC and viral infection.
Characterization of mESC-FBs generated through RA induction
mESC-DCs generated from EB differentiation contain mixed cell types [24]. Therefore, the limited IFNβ expression shown in mESC-DCs (Fig. 1B) may reflect a common property of different types of cells in the preparation. The 10T1/2 cells are FBs isolated from a 14–17 day old C3H mouse embryo (Fig. 2A, C3H-ME) [29,30]. For a direct comparison, we generated mESC-FBs from two independent mESC cell lines (D3 and DBA) through RA-induced differentiation. RA is a vitamin A derivative that regulates several developmental processes during embryogenesis and strongly induces ESC differentiation in vitro [2]. As shown in Fig. 2A, mESC-FBs (D3-FBs and DBA-FBs) completely lost the morphology of ESCs and formed a uniform monolayer that is very similar to that formed from10T1/2 cells. mESCs are characterized by their high proliferation rate with about 60% of cells in the S phase, whereas the proliferation of mESC-FBs was dramatically slowed down with an overall similar cell cycle profile to 10T1/2 cells as indicated by the reduced cell population at S and G2/M phases (Fig. 2B), although D3-FBs and DBA-FBs had slightly higher growth rate than 10T1/2 cells (Fig. 2C). More importantly, mESC-FBs expressed several FB markers with similar patterns to 10T1/2 cells as we previously described [13]. mESC-FBs can be continuously cultured for at least 15 passages. Cryopreserved cells can be further subcultured up to 55 passages. mESC-FBs at different passages exhibit consistent morphology when regularly split every 3–4 days. More than 95% of the cells express smooth muscle actin as a marker of FBs, similar to 10T1/2 cells [13]. Since D3-FBs and DBA-FBs are very similar in the properties that have been examined, the experiments that determine their antiviral mechanisms were mainly performed with D3-FBs (the parental D3 ESCs are one of the commonly used cell lines in the literature), whereas DBA-FBs were used for confirmative analysis in select experiments.
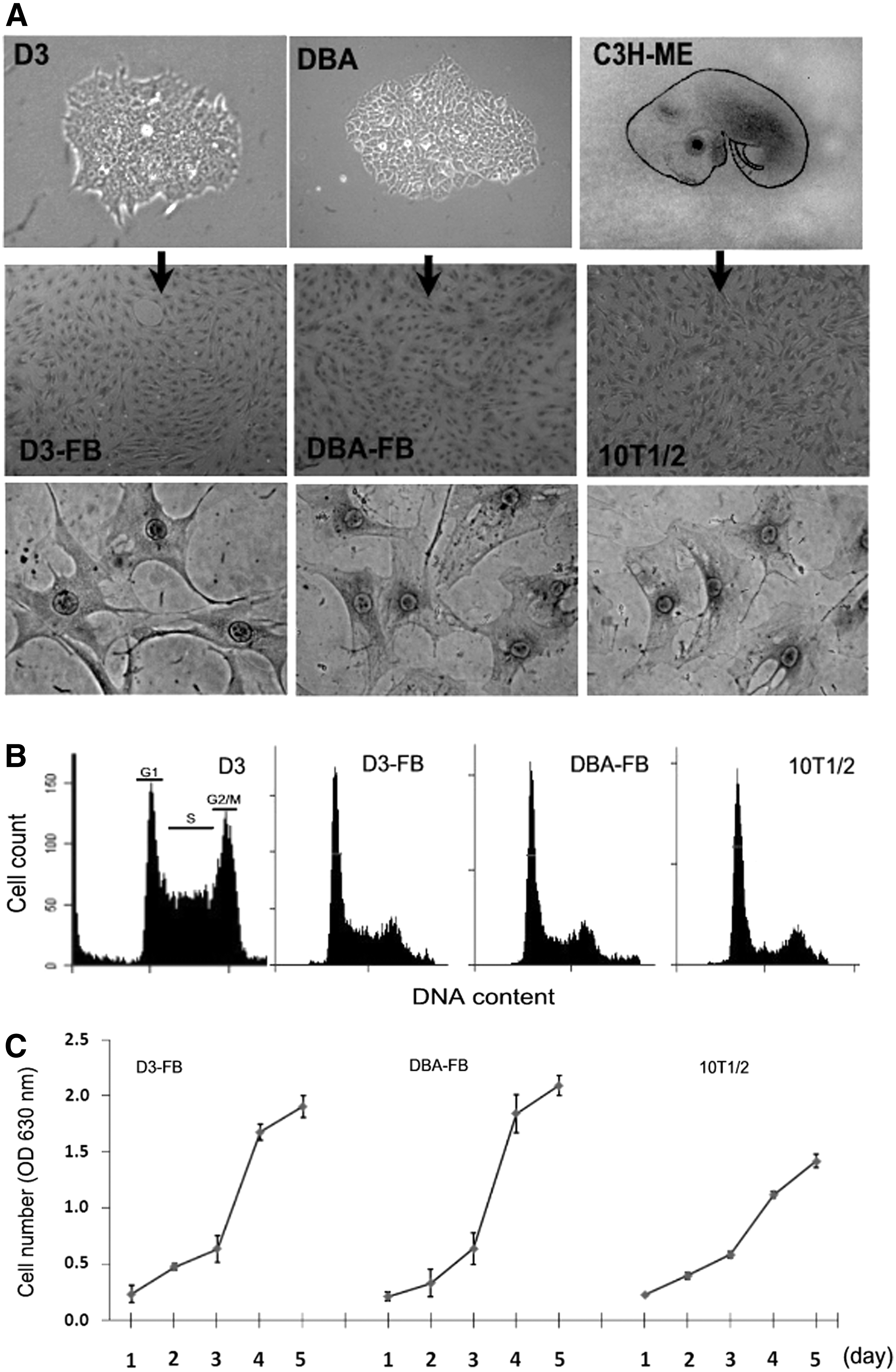
Generation and characterization of mESC-FBs.
Type I IFN expression mechanism is induced, but not effectively, during mESC differentiation
Although mESC-FBs showed considerable similarities to 10T1/2 cells in morphology, growth pattern (Fig. 2), and cell marker expression [13], mESC-FBs at the early passages (2–10 passages) expressed very low levels of IFNβ (Fig. 3A) and IFNα (data not shown) in response to polyIC, which was similar to mESC-DCs (Fig. 1B). At the mRNA level, the IFNβ expression capacity of D3-FBs was increased along with their continued in vitro propagation, but it was still substantially lower than 10T1/2 cells (Fig. 3A). It is known that signaling molecules that mediate immune responses are usually upregulated by the initial stimulus, forming a positive feedback loop that can in turn boost the cellular responses to subsequent immune challenges (known as the priming effect) [31 –33]. Based on this phenomenon, we primed D3-FBs with a low concentration of polyIC (50 ng/mL) for 24 h, a treatment that did not cause detectable cytotoxicity. After 3 days, the primed cells were split and replated in a new cell culture dish, where they were transfected with polyIC (300 ng/mL) a second time. As shown in Fig. 3B, the primed cells showed a significantly increased IFNβ mRNA expression compared with unprimed cells in response to the second dose of polyIC stimulation.
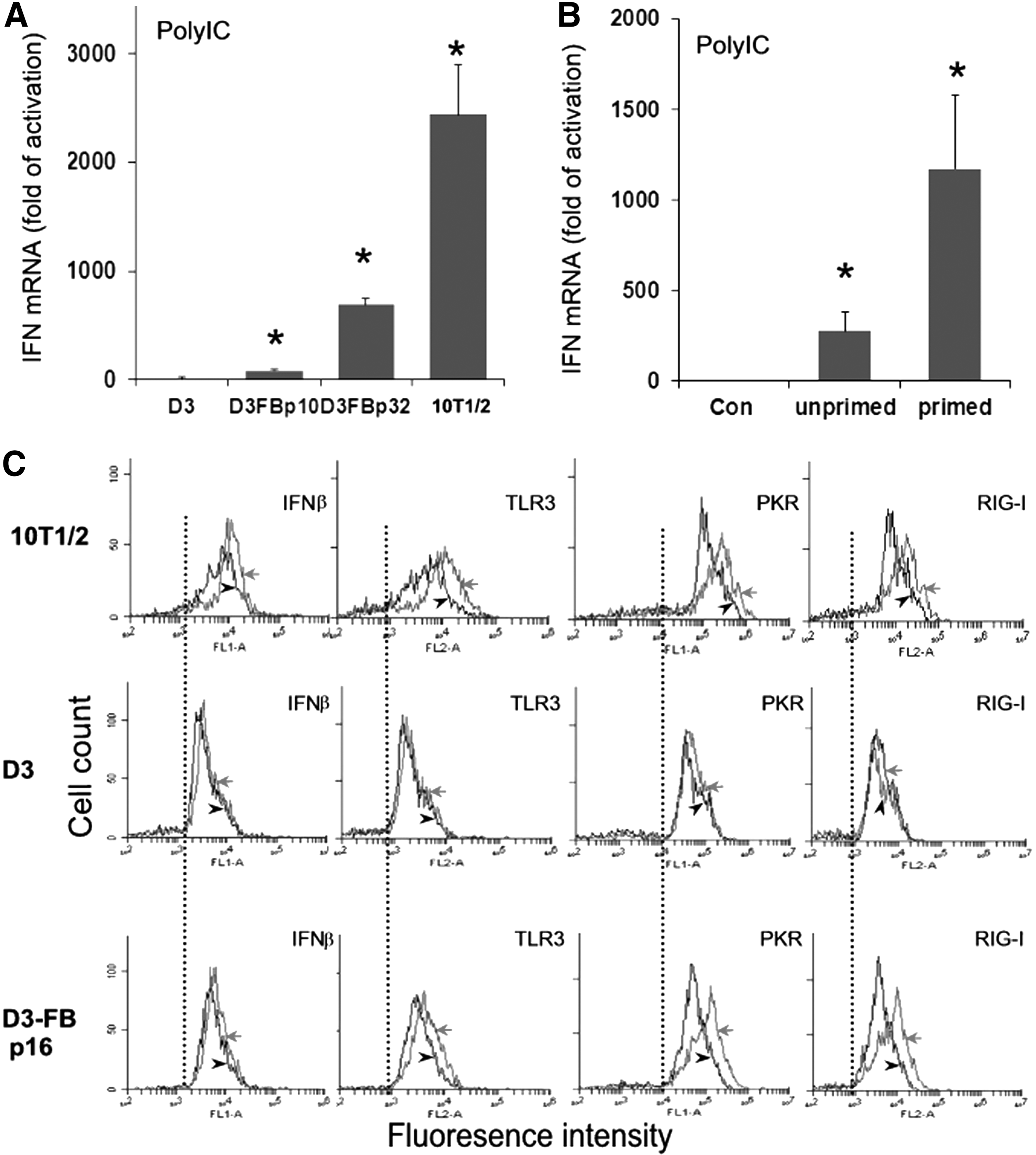
PolyIC-induced IFNβ expression in mESC-FBs and the effect of priming.
We further examined the expression of IFNβ and viral RNA receptors in response to polyIC at the protein level by flow cytometry. In 10T1/2 cells, polyIC induced the expression of IFNβ, concurrent with upregulation of three major viral RNA receptors: TLR3, RIG-I, and PKR as indicated by the increased florescence intensity (Fig. 3C, red lines). The same treatment did not induce the expression of these genes in D3 cells. However, in D3-FBs (p16), polyIC induced a significantly increased expression of PKR and RIG-I, to a lesser extent TLR3, but an increase of IFNβ was not detected (Fig. 3C). Similar results were obtained in D3-FBs passage 40 (data not shown). It is noted that the basal levels of TLR3, PKR, and RIG-I in control cells (black lines, the dotted lines were set as references) were the lowest in D3 cells. They are increased in D3-FBs, but lower than in 10T1/2 cells. Similar expression profiles for MDA5 were observed (data not shown). We speculate that the failure to detect IFNβ in D3-FBs by flow cytometry could be due to the low sensitivity of this method, which is unlike RT-qPCR that can detect very low-level changes of mRNA. Overall, our results suggested that the differentiation process could induce the IFN expression mechanism, but not to the level in naturally differentiated 10T1/2 cells. It is noted that transfection efficiency of polyIC is similar among D3, D3-FBs, and 10T1/2 cells as determined by the expression of eGFP from its synthetic mRNA transfected to these cells (data not shown) with the method that we have previously described [11]. Therefore, the different expression levels of IFN in response to polyIC transfection among the three cell types are attributed to their intrinsic properties.
Upregulation of viral RNA receptors was also observed when mESC-FBs were infected with live viruses. In response to LACV infection, IFNβ expression in early passage D3-FBs (p5) was slightly higher than in undifferentiated D3 cells, but was markedly increased in the cells at passage 50 (Fig. 4A). Like polyIC priming, LACV infection also stimulated the expression of viral RNA receptors (Fig. 4B). In addition to LACV, we also analyzed the cell response to the infection of CHIKV, a virus that is known to effectively infect FBs, and a similar pattern of IFNβ expression to LACV was observed (Fig. 4C).
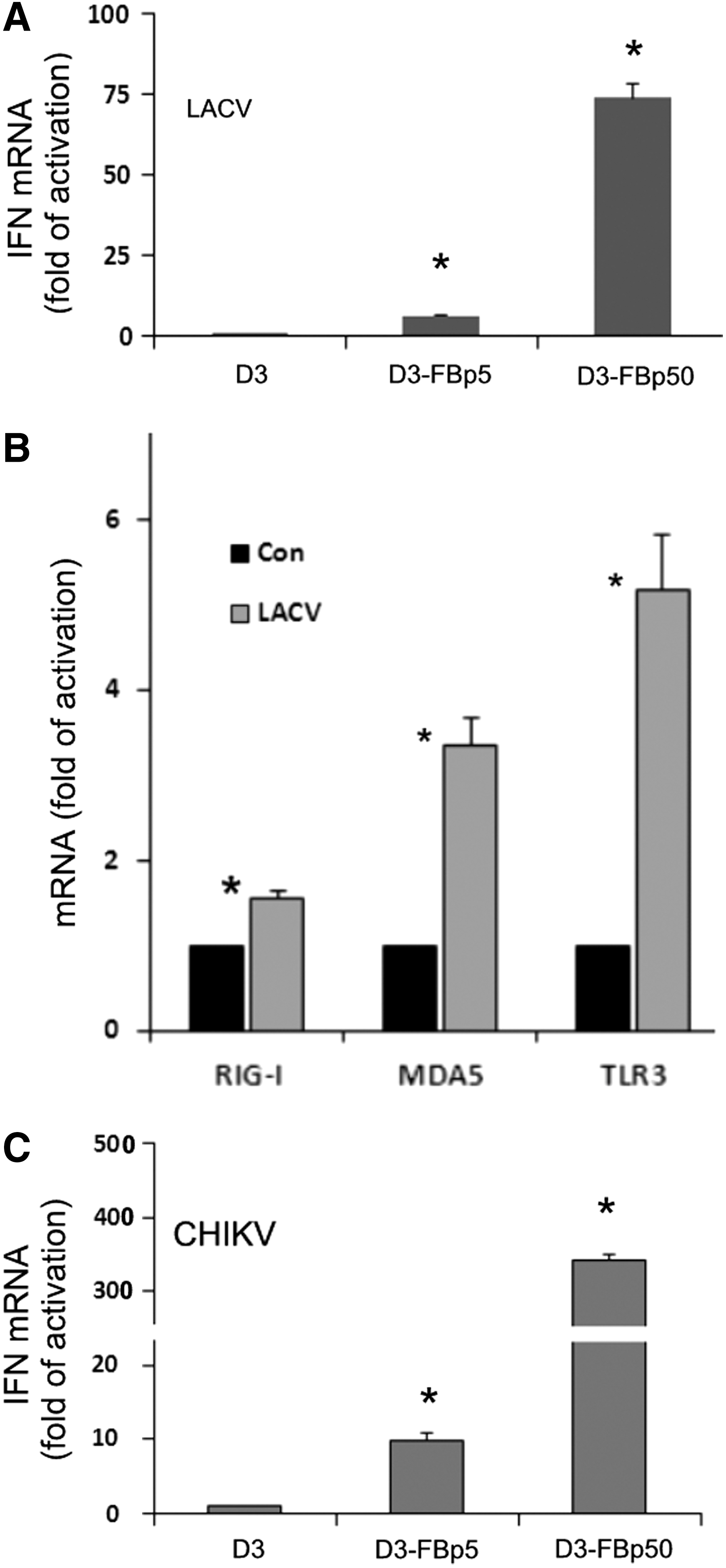
Viral infection-induced expression of IFNβ and viral RNA receptors in mESC-FBs. D3 cells, and D3-FBs at the indicated passages were infected with LACV
The NFκB pathway is not functional in mESCs, but is activated in mESC-FBs in response to viral infection
The above results indicated that the molecular mechanism that mediates IFN expression is inducible, although not effectively, by the differentiation process. In seeking the molecular mechanism that underlies this observation, we analyzed the activation status of NFκB in response to viral infection. NFκB is an essential transcription factor for IFN expression [34,35]. Taking advantage of the dramatic difference in morphology between mESCs and mESC-FBs, we used a coculture model, where the colony of D3 cells can be easily distinguished from the large flattened D3-FBs in the same dish as illustrated in Fig. 5A (Con, phase). The cocultured D3 and D3-FBs were exposed to CHIKV or LACV under the identical condition. The infected cells were identified by the antibodies that recognized a CHIKV or LACV-specific protein (AB3583 or Gc, bright green color). In undifferentiated control cells, NFκB was detected in the cytoplasm of both D3 cells and D3-FBs (Con, red color). Its activation, as indicated by its detection in the nucleus, only took place in viral-infected D3-FBs (CHIKV and LACV, indicated by arrow), but not in viral-infected D3 cells (CHIKV and LACV, indicated by arrowhead). It is noted that uninfected cells, both D3 cells and D3-FBs, had their NFκB retained in the cytoplasm (CHIKV and LACV, cells with dark green background) as seen in the control cells. Quantitative analysis of CHIKV-infected cells showed that about 65% of infected D3-FBs were positive for nuclear NFκB staining, whereas essentially no nuclear NFκB was detected in any CHIKV-infected D3 cells (Fig. 5B), regardless of the percentage of infected cells in colonies with different sizes. Similar results were observed in cells infected with LACV (data not shown). The exclusive detection of NFκB in the nucleus of viral-infected D3-FBs clearly demonstrated that the NFκB pathway is functional only after differentiation. The same results were obtained when D3 cells and D3-FBs were cultured separately (data not shown).
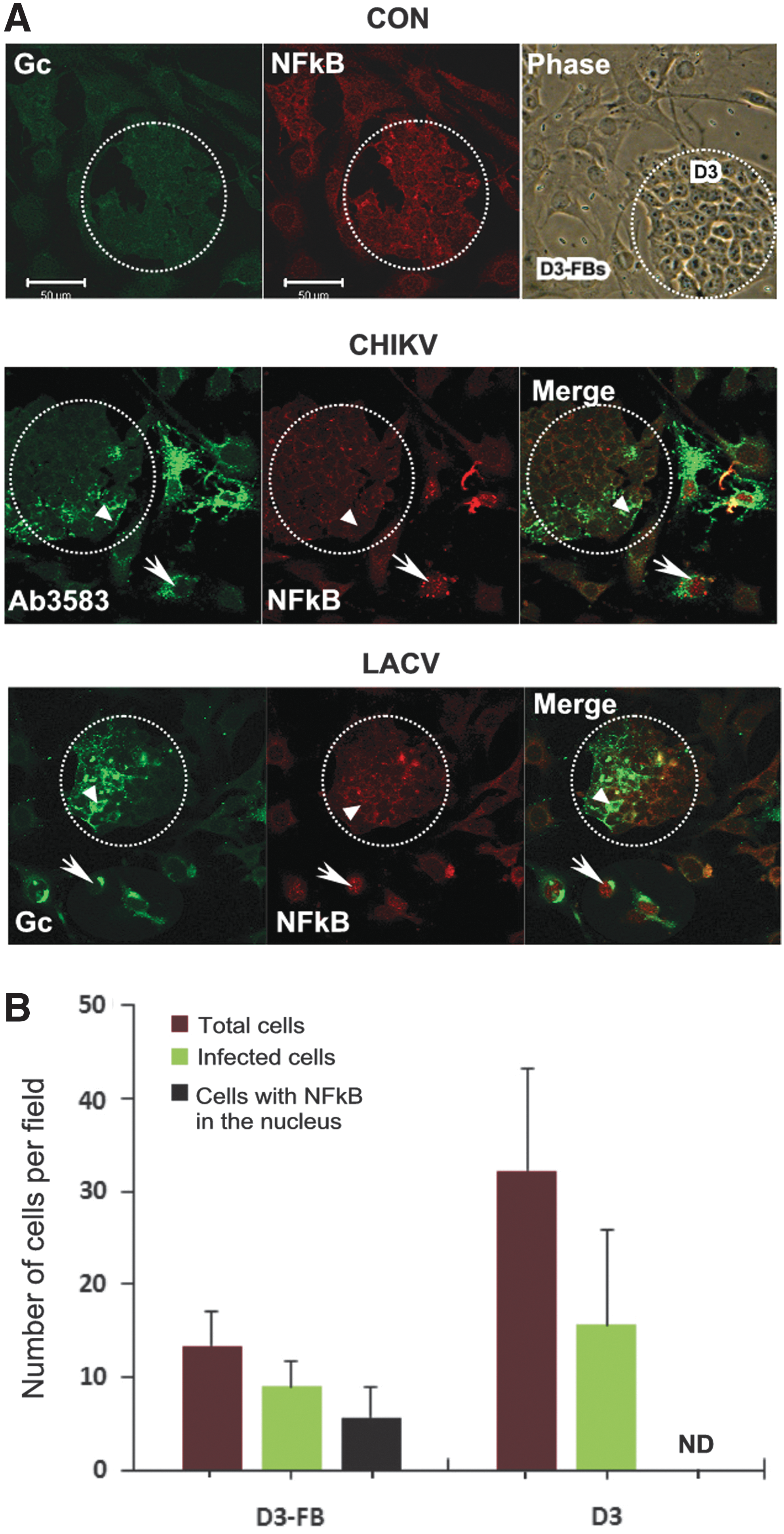
The activation status of NFκB in mESCs and mESC-FBs in response to viral infection.
mESC-FBs are susceptible to cytopathic effects of viral infection and are protected by type I IFN
We compared the responses of D3-FBs and 10T1/2 cells to the infection with LACV and CHIKV. Both viruses caused cell death as indicated by the reduction of cell number (Fig. 6A) and morphological changes (Fig. 6B). Cells infected with CHIKV were characterized by the lysis of the cytoplasm, whereas the nucleus was mostly intact until the late stage of cell death (Fig. 6B, CHIKV). LACV infection caused a similar cytopathic effect on mESC-FBs, but with less potency (data not shown). Overall, mESC-FBs were less susceptible to the cytopathic effects of CHIKV and LACV than 10T1/2 cells, but they showed the same pattern of lytic cell death at a longer incubation time.
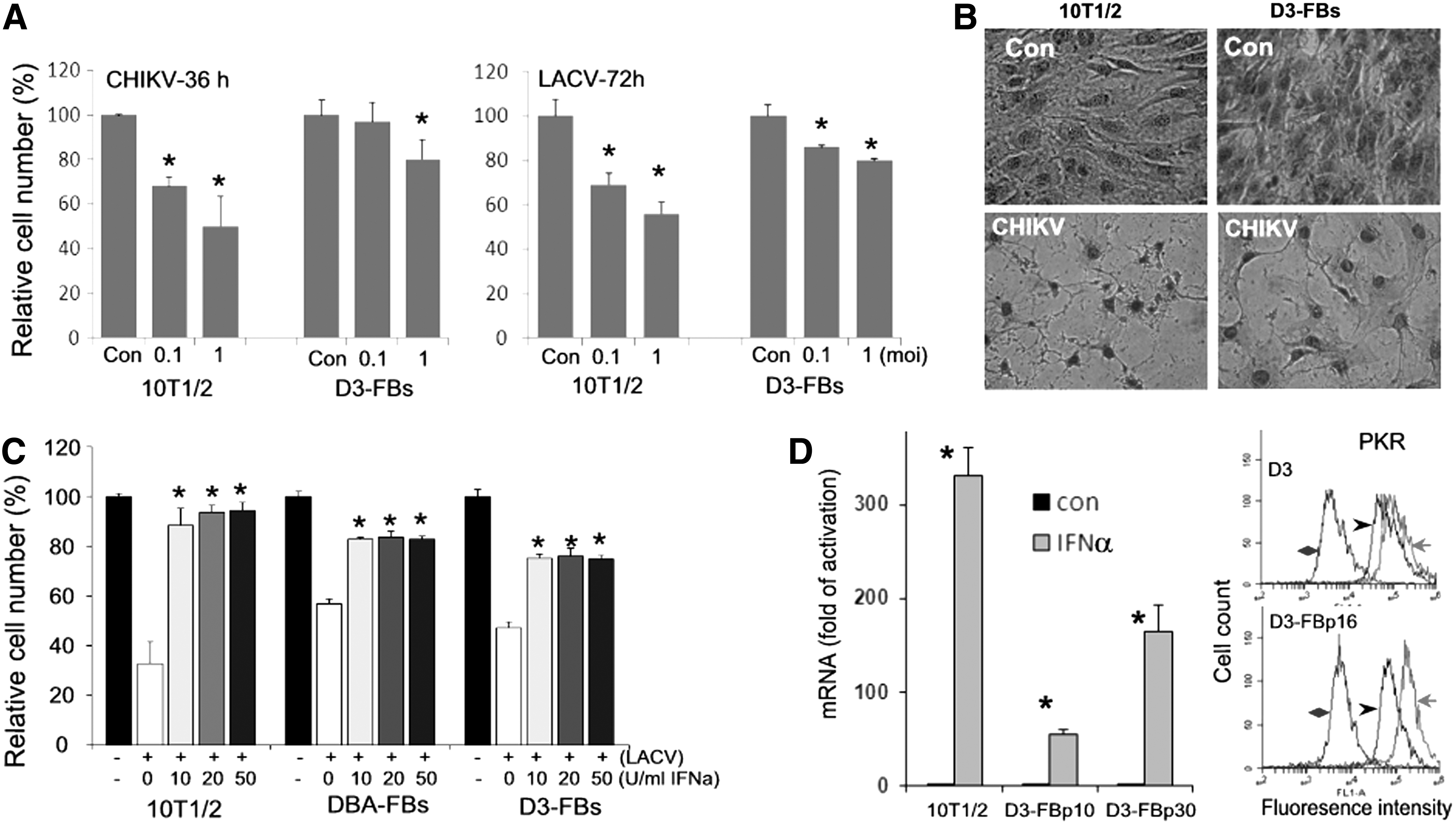
Viral infection-induced cytopathicity and the effect of IFNα.
Type I IFN are best characterized for their antiviral activity. We have previously shown that although mESCs cannot express these cytokines [12], they could weakly respond to IFN and express IFN-stimulated genes (ISGs). mESCs were protected from LACV- and CHIKV-induced cell death by exogenously added IFNα, but at a concentration that was much higher than that needed to protect 10T1/2 cells [13]. Therefore, we tested the effects of IFNα on viral-infected mESC-FBs. As shown in Fig. 6C, pretreatment with IFNα significantly reduced LACV-induced cell death of mESC-FBs at 10 U/mL. This concentration is about 10 times lower than that needed to protect mESCs from LACV-induced cell death (>100 U/mL) [13]. Similar results were observed in CHIKV-infected mESC-FBs (data not shown).
Correlating with the increased ability to mediate the antiviral activity of endogenously added IFNα, D3-FBs express higher mRNA levels of ISG15 than in D3 cells in response to IFNα (Fig. 6D, graph). We further examined the expression of PKR, which is another well-characterized ISG [36]. IFNα induced PKR expression in D3 cells as determined by flow cytometry, however, the induction in D3-FBs is significantly higher than in D3 cells, as judged by the greater increase of fluorescence intensity relative to control cells (red line vs. black lines, Fig. 6D, flow profile). Together, these data support the conclusion that mESCs can respond to IFN, but at a much lower level than differentiated cells. However, the IFN response mechanism is further developed in mESC-FBs after differentiation.
PolyIC-induced cell death of mESC-FBs is mediated by PKR
We have previously shown that polyIC transfection activated PKR in mESCs and caused cell cycle inhibition, but did not cause apparent cell death [12]. However, mESC-FBs showed an increased sensitivity to polyIC-induced cell death similar to 10T1/2 cells, as indicated by the reduced cell number (Fig. 7A). Unlike the cell death induced by viral infection that was attenuated by IFNα treatment (Fig. 6A), polyIC-induced cell death was not affected by IFNα (Fig. 7A) or by IFNβ (data not shown). Furthermore, polyIC-induced cell death was characterized by the formation of multiple intracellular vacuoles while the cytoplasm is largely intact at the earlier stage of cell death (Fig. 7B). These results indicate that the protecting activity of IFN is limited to the cytopathic effect of viral infection, suggesting that the cell death caused by viral infection and polyIC transfection occurs through different mechanisms.
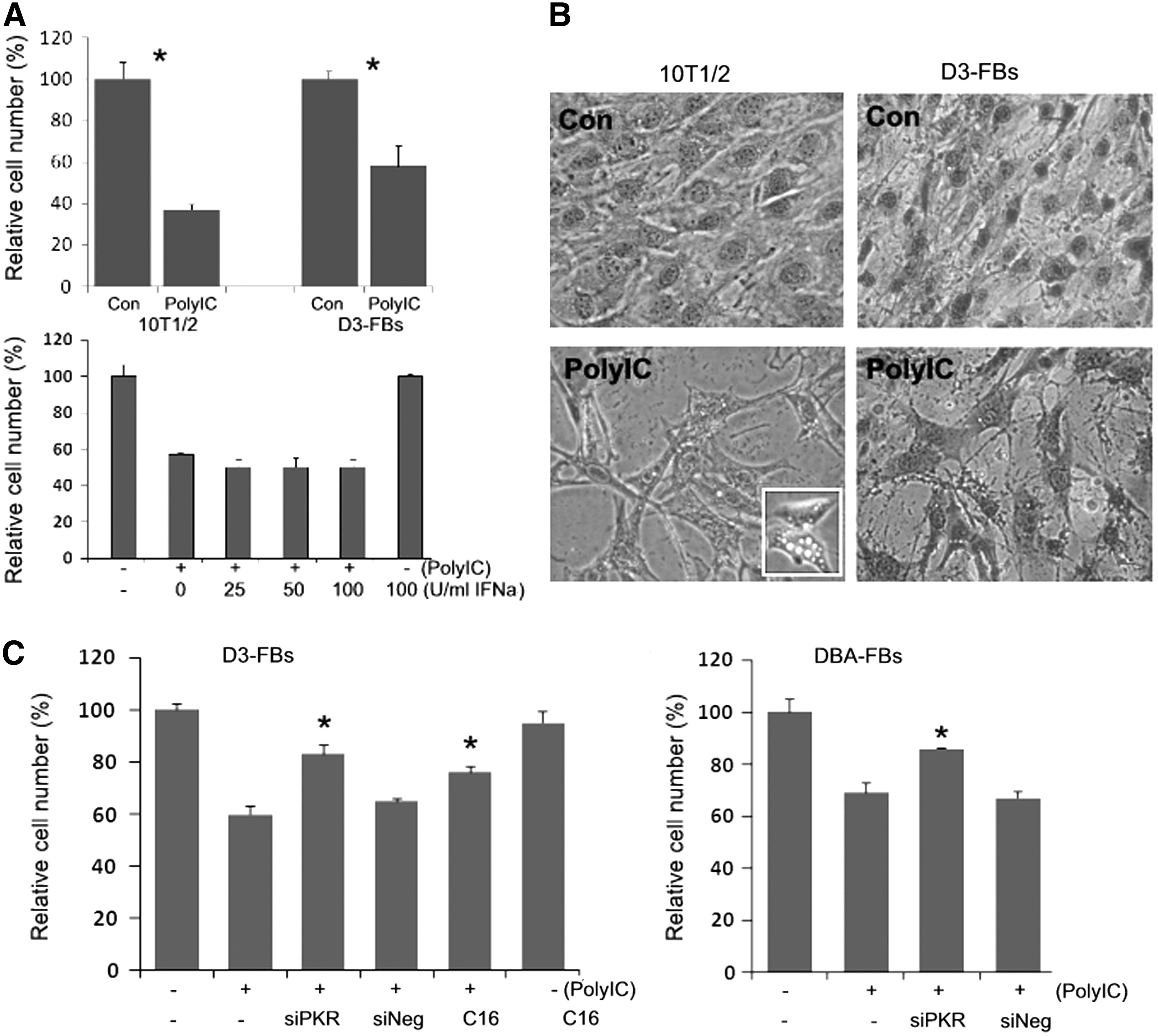
PolyIC-induced cytotoxicity and the role of PKR.
Activation of PKR has been implicated in polyIC-induced death of somatic cells [37]. Since polyIC significantly increased the expression of PKR in D3-FBs, but not in D3 cells (Fig. 3B), it is likely that the increased PKR expression contributes to polyIC-induced cell death in mESC-FBs. To test this hypothesis, we used RNAi to knock down PKR in mESC-FBs. In a previous study, we have shown that transfection of mESCs with a cocktail of three different sequence-specific siRNAs against PKR could specifically knock down PKR to a level undetectable by western blot analysis [12]. The same treatment significantly attenuated polyIC-induced cell death in both D3-FBs and DBA-FBs. A similar result was obtained when the cells were treated with a PKR inhibitor (C16) (Fig. 7C). These results suggested that PKR activation is at least partly responsible for polyIC-induced cell death of mESC-FBs.
Discussion
The underdeveloped innate immunity has been recently characterized as a unique property of pluripotent cells as we have recently discussed (reviewed in Guo et al. [17]). Since most types of somatic cells have an effective innate immune system, the attenuated immune and inflammatory responses reported in several ESC-derived tissue cells [3 –6] raise the question of whether the commonly used in vitro differentiation methods can generate cells with competent innate immunity like in vivo differentiated cells. However, it is not clear whether these findings represent all types of ESC-derived cells or just the observations in the cell types that have been investigated. Furthermore, none of the published studies has investigated antiviral responses under the condition of live viral infection. Herein, we provide valuable insights into the molecular basis for the deficiency of IFN expression in mESCs and the development of antiviral innate immunity during their differentiation.
Differentiation of ESCs through EB formation is the most commonly used method in the literature [1]. Our results demonstrated that mESC-DCs by this method showed a limited capacity to express IFNβ, which likely represents a common property of multiple cell types since mESC-DCs consist of mixed cell lineages [24]. We used 10T1/2 cells for comparison for two reasons. First, their embryonic origin makes them particularly suitable since they were FBs derived from 14–17 day old mouse embryos [29,30], a time frame similar to in vitro EB differentiation. Second, FBs are considered to be the major tissue stromal cells that modulate tissue immunity and inflammation since they express large amounts of IFNβ and other inflammatory cytokines in response to immune stimuli [38,39]. The mESC-FBs that we generated through RA-induced differentiation share extensive similarities with 10T1/2 cells, but these cells, especially at early passages, displayed a significantly lower capacity to express IFNβ than 10T1/2 cells, similar to mESC-DCs differentiated from EB formation. Therefore, the lack of an effective IFN expression capacity is a major property of mESC-FBs that differs from naturally differentiated 10T1/2 cells. While the mechanism to express type I IFN during differentiation lagged behind the development of other cellular features (ie, morphology, proliferation rate, cell cycle profile, and cell marker expression), it was nonetheless inducible. Our analysis of NFκB in mESCs and mESC-FBs provides an explanation for the deficiency of type I IFN expression in undifferentiated mESCs and the acquisition of its active status in differentiated mESC-FBs. NFκB is an essential transcription factor for the expression of IFN and inflammatory cytokines. Although the functionality of NFκB in ESCs have been an issue of controversy [40,41], our data clearly demonstrated its inactive status in viral-infected mESCs, which was also noted by other investigators in mESCs that were treated with TNFα [42], the best studied NFκB activator in differentiated somatic cells. We believe that the low expression levels of viral RNA receptors [12] (Fig. 3C) and the inactive status of NFκB could be major reasons for the attenuated antiviral responses in mESCs. However, we cannot rule out the possibility that there are other components essential for innate immunity that may not be sufficiently expressed or not functional as well.
The upregulation of viral RNA receptors and activation of NFκB by viral infection in mESC-FBs signifies the acquisition of antiviral innate immunity during differentiation. Our data suggest that the two commonly used in vitro differentiation methods as described in this study can only partly turn on the genes that regulate immune responses. We speculate that the reason for this phenomenon could be that the expression of these genes is not needed because the differentiation takes place under a sterile culture condition. It is known that the expression of immune genes can be upregulated by pathogenic stimuli [31 –33]. PolyIC treatment, like acute viral infection, can upregulate the expression of certain immune genes in immune cells and FBs [33,43]. Similar to this finding, polyIC priming induced expression of viral RNA receptors in mESC-FBs, resulting in an increased response to subsequent polyIC transfection. We speculate that the priming effect of polyIC is likely through a mechanism that was recently described in FBs, by which polyIC induces the expression of multiple innate immune genes through chromatin remodeling [43]. Similar to polyIC, viral infection also induced the expression of viral RNA receptors. Therefore, we propose that the development of antiviral mechanisms can be accelerated by immunostimuli (ie, polyIC or viral infection) during ESC differentiation.
As part of the IFN-based antiviral mechanism, the capacity to respond to IFN is underdeveloped, but not completely deficient in mESCs [13]. Our data indicated that the IFN response mechanism is further advanced in mESCs-FBs, as indicated by their increased ability to mediate the antiviral effect of IFNα and to induce the expression of ISGs (ISG15 and PKR). It is interesting to note that IFN treatment showed the expected antiviral activity against viral-induced cell death of mESC-FBs, but were unable to protect the cells from polyIC-induced cell death. While the cell death caused by viral infection is a very complicated process that could involve numerous proteins and is often virus-dependent [7], we believe that polyIC-induced cytotoxicity in mESC-FBs is likely involved in PKR upregulation/activation, which is known to cause cell death of differentiated somatic cells [37]. We have previously shown that polyIC causes cell cycle inhibition, but not significant cell death [12], therefore, the increased sensitivity of mESC-FBs to the toxicity of polyIC is likely due to their higher expression level of PKR than in mESCs in response to both polyIC and IFNα. These results suggest that the differentiation process also promotes the development of cellular mechanisms regulated by the PKR pathway.
Most of the current differentiation methods use certain growth factors or cytokines to promote ESC differentiation toward a particular cell lineage of interest. For example, VEGF and bFGF are commonly used to promote ESC differentiating into vascular endothelial cells [44 –47], whereas PDGF and TGFβ are used for smooth muscle cell differentiation [44,48]. We have not been aware of any strategies for the differentiation of structural tissue cells that contain factors with the consideration of innate immunity development. We speculate that lacking immunostimulation during in vitro differentiation could be a major reason that accounts for the ineffective development of innate immunity in ESC-derived cells.
The attenuated innate immunity in ESC-derived cells is a complex issue that may have different implications for their therapeutic application. On one hand, it is known that a strong immune and inflammatory reaction of grafted cells to the host environment could augment the inflammation in the wounded area, therefore, negatively impacting tissue repair and healing. In this context, an attenuated innate immunity in ESC-derived cells could be beneficial. On the other hand, the functionality of transplanted ESC-derived cells could be compromised if they do not have a competent innate immunity. For instance, endothelial cells are not only critical for vascular function, but also act as innate immune surveillance cells to sense and combat pathogens in the circulation [49,50]. Conceivably, the use of ESC-derived endothelial cells for vascular tissue repair could be a concern if they do not have a competent innate immunity [3]. At the present time, it is not clear to what degree the attenuated innate immunity that we have characterized in ESC-FBs applies to other ESC-derived cell types and how this property may affect their cell type-specific functions, such as the aforementioned endothelial cells. To fully understand these questions, it will be essential to have a complete characterization of ESC-derived cells by in vitro and in vivo studies.
Conclusions
The lack of a functional innate immunity in ESCs has recently attracted much attention as an important subject in basic ESC biology, immunobiology, developmental biology, and regenerative medicine. Based on our previous work and current study, we conclude that mESCs have underdeveloped antiviral mechanisms; they are deficient in expressing type I IFN, have limited responsiveness to IFN, and have a basic functional PKR pathway. These antiviral mechanisms can be induced during differentiation and could be modulated by the differentiation environment. This knowledge is not only valuable to understand basic ESC biology, but also instructive to design strategies that can generate ESC-derived cells with desired levels of innate immunity suitable for regenerative medicine.
Footnotes
Acknowledgments
This work was supported a grant from the National Institutes of Health (1R15GM109299-01A1 to Y-LG). The authors thank Dr. John F. Anderson (Connecticut Agricultural Experiment Station) for providing LACV, Dr. Robert B. Tesh (University of Texas Medical Branch) for providing CHIKV, and Dr. Samantha Soldan (University of Pennsylvania School of Medicine) for providing LACV Gc antibodies. They also thank Mississippi-IDeA Network of Biomedical Research Excellence for the use of the imaging facility.
Author Disclosure Statement
No competing financial interests exist.