
Abstract
Mesenchymal stem cells (MSC) have emerged as alternative sources of stem cells for regenerative medicine because of their multipotency and strong immune-regulatory properties. Also, human leukocyte antigen G (HLA-G) is an important mediator of MSC-mediated immunomodulation. However, it is unclear whether MSC retain their immune-privileged potential after differentiation. As promising candidates for cartilage tissue engineering, the immunogenic and immunomodulatory properties of chondro-differentiated MSC (chondro-MSC) require in-depth exploration. In the present study, we used the alginate/hyaluronic acid (Alg/HA) hydrogel scaffold and induced both bone marrow- and adipose tissue-derived MSC into chondrocytes in three-dimensional condition. Then, MSC before and after chondrocyte differentiation were treated or not with interferon γ and tumor necrosis factor α mimicking inflammatory conditions and were compared side by side using flow cytometry, mixed lymphocyte reaction, and immunostaining assays. Results showed that chondro-MSC were hypoimmunogenic and could exert immunosuppression on HLA-mismatched peripheral blood mononuclear cells as well as undifferentiated MSC did. This alloproliferation inhibition mediated by MSC or chondro-MSC was dose dependent. Meanwhile, chondro-MSC exerted inhibition on natural killer cell-mediated cytolysis. Also, we showed that HLA-G expression was upregulated in chondro-MSC under hypoxia context and could be boosted in allogenic settings. Besides, the Alg/HA hydrogel scaffold was hypoimmunogenic and its addition for supporting MSC chondrocyte differentiation did not modify the immune properties of MSC. Finally, considering their chondro-regenerative potential and their retained immunosuppressive capacity, MSC constitute promising allogenic sources of stem cells for cartilage repair.
Introduction
J
Numerous studies have shown that MSC have low immunogenicity and exert immunosuppressive function in vitro and in vivo [5 –7]. Moreover, the use of MSC for clinical applications has proven effective in acute and severe graft-versus-host disease [8]. So far, both contact-dependent mechanism and soluble factors have been reported to contribute to MSC-mediated immune regulation [6]. Cell–cell contact inhibition on lymphocytes has been reported via the adhesion molecules expressed on MSC that activate the programmed death 1 pathway [9]. Soluble factors secreted from MSC, including nitric oxide [10], indoleamine 2,3-dioxygenase, prostaglandin E2 [11], transforming growth factor-β1 (TGFβ1), hepatocyte growth factor [12], IL10 [13], heme oxygenase-1 [14], IL6 [15], and soluble human leukocyte antigen G5 (sHLA-G5) [16], have been involved in MSC-mediated regulation of immune response. Of note, HLA-G5 plays a critical role in MSC-mediated immunomodulation by suppressing T- and natural killer (NK) cell function and by inducing T regs [16,17].
However, it is unclear whether MSC retain their immune-privileged potential after multidifferentiation. Several studies have shown conflicting results regarding the immune properties of differentiated MSC [18 –20], which may hamper their clinical application. The goal of the present study was to figure out both immunogenicity and immunosuppressive function of MSC derived from bone marrow (BM) or adipose tissue (AT) (MSC-BM/-AT) after chondrocyte differentiation. For this purpose, MSC-BM/-AT were combined with biomaterial scaffold to induce chondrogenesis under hypoxia conditions. Then, chondro-MSC were harvested to study their immunogenic and immunosuppressive properties in allogenic settings. In all experiments, we analyzed the influence of inflammatory conditions [through interferon γ (IFNγ) and tumor necrosis factor α (TNFα) pretreatment] on chondro-MSC immune properties. Also, we explored the role of HLA-G in both MSC- and chondro-MSC-mediated immune regulation. Finally, the property of alginate/hyaluronic acid (Alg/HA) hydrogel scaffold whether immunogenic or toxic in terms of triggering immune response facing allogenic peripheral blood mononuclear cells (PBMC) was also investigated.
Materials and Methods
MSC culture and chondrocyte differentiation
This study was performed according to the Declaration of Helsinki and approved by the Ethics Committee and the Institutional Review Board of Commissariat a l'énergie atomique (CEA). Both MSC-BM and MSC-AT obtained from the REBORNE consortium center (
For immunocytochemical staining, MSC-BM/-AT and chondro-MSC-BM/-AT [involved in mixed lymphocyte reaction (MLR) or not] were suspended in α-MEM at 0.5 × 106–1.0 × 106 cells/mL. Then, cytospins were made using Superfrost/plus® slides (Merck) and Cytospin 3® (Shandon). After fixing with ice acetone for 15 min, slides were dry in the air and frozen at −20°C. The left cells were used in MLR and flow cytometry assays.
Alcian blue staining
Chondro-MSC-BM/-AT on slides were stained with the Alcian blue solution for 30 min at room temperature. Then, slides were observed under a microscope (Leica), and photos were taken under × 10 magnification. Chondro-MSC-BM/-AT, in contrast to undifferentiated MSC-BM/-AT expressed glycosaminoglycan and were stained with Alcian blue.
Quantitative real-time polymerase chain reaction
Cells were extracted from Alg/HA hydrogels by chelating Ca2+ in alginate egg-box and were washed three times with PBS. RNA was extracted by using the RNeasy extraction kit (Qiagen) and further purified by the DNase kit (Qiagen). Then, cDNA was synthesized by reverse-transcription of mRNA by using the Iscript kit (BioRad). Finally, quantitative real-time polymerase chain reaction (qRT-PCR) was performed with the Quantities SYBR Green PCR kit (Qiagen) and cDNA obtained from samples, on a Light Cycler system (Roche Diagnostics) during 45 cycles to quantitatively analyze the expression of Sox9, aggrecan (Agg), type II collagen, and cartilage oligomeric matrix protein (COMP). Primers for qRT-PCR were listed as follows: RP29 forward primer: 5′-AAGATGGGTCACCAGCAGCTCTACTG-3′; reverse primer: 5′-AGACGCGGCAAGAGCGAGAA-3′. Sox9 forward primer: 5′-GAGCAGACGCACATCTC-3′; reverse primer: 5′-CCTGGGATTGCCCCGA-3′. Agg forward primer: 5′-TCGAGGACAGCGAGGCC-3′; reverse primer: 5′-TCGAGGGTGTAGCGTGTAGAGA-3′. Total type II collagen (Col II total) forward primer: 5′-ATGACAATCTGGCTCCCAAC-3′; reverse primer: 5′-GAACCTGCTATTGCCCTCTG-3′. COMP forward primer: 5′-ACAATGACGGAGTCCCTGAC-3′; reverse primer: 5′-TCTGCATCAAAGTCGTCCTG-3′. All cartilage-related gene expression was normalized to the expression of housekeeping gene RP-29.
Peripheral blood mononuclear cell
Blood was collected from healthy volunteers from the French Blood Establishment (EFS, Saint-Louis Hospital) after informed consent. Based on the density-gradient centrifugation, PBMC were isolated following the instructions of Ficoll-Paque Plus (GE Healthcare). PBMC were used as responding cells in MLR.
Mixed lymphocyte reaction
MLR was performed to test both immunogenic and immunoregulatory properties of MSC and chondro-MSC, licensed with inflammatory cytokines or not. For this purpose, MSC or chondro-MSC pretreated or not with 20 ng/mL IFNγ (Peprotech) and 30 ng/mL TNFα (R&D system) for 48 h were used as stimulator cells (immunogenicity assay) or third-party cells (immunosuppression assay) toward HLA-mismatched PBMC (responder cells). Human B lymphoblastoid cell line (LCL) 721.221 (ATCC), HLA class II-positive cells, was irradiated at 75 Gy dose to be used as stimulator in MLR. LCL cells stimulating allogenic PBMC were regarded as maximal MLR (100% alloproliferation). The ratio of PBMC and LCL seeded in each well of 96-well plate was 1:0.5, with a final concentration of 105 PBMC/well. In addition, MSC and chondro-MSC pretreated with inflammatory cytokines or not were prepared at various ratios (0.5 × 105, 0.25 × 105, 0.125 × 105, 0.06 × 105, and 0.03 × 105). To test the role of HLA-G played in MSC-mediated immunosuppression, 20 μg/mL 87G (anti-HLA-G antibody; Exbio) or isotype control antibody (Exbio) was added to MLR. MLR lasted for 6 days at 37°C in a humidified 5% CO2 air atmosphere. On day 5, [3H]-thymidine (1 μCi/well; PerkinElmer) was added to each well and incubated for another 18 h. Cells were then harvested on filtermats A, and thymidine incorporation into DNA was quantified using a β counter (Wallac 1450; Pharmacia). All experiments were performed in triplicate.
In addition, to evaluate whether hydrogel scaffold affected the immune properties of MSC, MSC-BM/-AT combined with Alg/HA hydrogel scaffold or not were put in MLR. MSC-BM/-AT alone were used as control. Three MSC-BM/-AT concentrations were performed (0.4 × 105, 0.2 × 105, 0.1 × 105) in MLR.
To test HLA-G expression, MSC-BM/-AT or chondro-MSC-BM/-AT were seeded in a six-well plate at 5 × 105cells/well, and then, 5 × 105 irradiated LCL and 1.0 × 106 PBMC were added to MSC to initiate the immune response. After 6 days of incubation, MSC-BM/-AT and chondro-MSC-BM/-AT were harvested to evaluate HLA-G expression by flow cytometry.
Cytotoxicity assays
The cytolysis activity of peripheral blood NK cells against the K562 (human erythroleukemia cell line, ATCC) target cells was assessed in 4-h 51Cr-release assays, in which effector NK cells (25 × 104cells) were mixed with 5,000 51Cr-labeled targets [100 μCi of 51Cr sodium chromate (PerkinElmer)] in U-bottomed microliter plates. NK cells were obtained from three distinct healthy donors. For inhibition assays, MSC-derived chondrocytes from two distinct BM samples were previously incubated at various ratios with effector cells (NK cells) for 1 h at 37°C, and then, 51Cr-labeled targets (K562 cells) were subsequently added to the coculture. All three cell types were maintained together during the 4-h cytotoxicity assay at 37°C in a humidified 5% CO2 incubator. Finally, 50 μL of the supernatant was collected for liquid scintillation counting (Wallac 1410; Pharmacia). The percentage of specific cytolysis was calculated as follows: specific cytolysis (%) = (count per minute [CPM]Experimental release − CPMSpontaneous release)/(CPMMaximum release − CPMSpontaneous release) × 100%. Spontaneous release was determined by incubation of labeled target cells with medium. Maximum release was determined by solubilizing target cells in 0.5% Triton X-100. Results are presented as mean ± SEM; each sample was performed in triplicate.
Flow cytometry
Flow cytometry was used to detect the expression of costimulatory molecules, including CD40, CD80, and CD86, as well as MHC-II molecules (HLA-DR) and HLA-G on MSC or chondro-MSC (primed with IFNγ and TNFα or not, and involved or not in MLR). The indicated phycoerythrin (PE)-conjugated mAb (Beckman Coulter) was incubated at 4°C for 30 min with MSC-BM/-AT or chondro-MSC-BM/-AT prelicensed or not with cytokines. PE-conjugated IgG2a was used as isotype control. LCL cells (expressing high level of HLA-DR) were used as positive cells.
Besides, HLA-G intracytoplasmic expression level on MSC-BM/-AT or chondro-MSC-BM/-AT (primed or not with cytokines, and involved or not in MLR) was assessed by flow cytometry. Cells were fixed in 3% paraformaldehyde for 20 min at room temperature. Then, cells were permeabilized by 0.1% saponin (Sigma) for 20 min. Later, the anti-HLA-G5/-G6 2A12 mAb (1:100 dilution; Exbio) prepared in 0.1% saponin solution was added to cells for 30 min at 4°C. After washing twice with 0.1% saponin, cells were stained with PE-conjugated rabbit anti-mouse IgG antibodies (1:100 dilution; Beckman Coulter) for another 30 min. Cells incubated with the second antibodies were used as negative control. M8G5 cells (expressing high levels of HLA-G5) were used as positive control.
To test HLA-G membrane-bound expression on MSC (involved in MLR), the fluorescein isothiocyanate (FITC)-conjugated anti-CD14 antibody (Beckman Coulter) and PE-anti-HLA-G MEMG9 (Beckman Coulter) were incubated with cells for 30 min at 4°C. Then, cells were collected for flow cytometry analysis. In addition, to define the expression of sHLA-G5 by MSC or chondro-MSC, cells were fixed, permeabilized, and stained with the anti-HLA-G5 mAb 2A12 for 30 min. After incubation with PE-conjugated anti-mouse antibody for another 30 min, cells were washed and further labeled with anti-CD14-FITC for 30 min. Finally, cells were detected by using the Epic XL4 flow cytometer.
Immunocytochemical staining
To confirm chondrogenic induction, anti-type II collagen antibody (Collagen II, Thermal) was diluted at 1:100 and incubated with slides from MSC-BM/-AT and chondro-MSC-BM/-AT for 30 min followed by permeabilization with Tween-Tris-HCl solution. Then, slides were incubated with the anti-mouse PE-conjugated antibody for 20 min. After washing, slides were stained with DAPI (Vectashield; Vector) and analyzed by confocal microscopy (Leica) under × 20 magnification. Similarly, slides of MSC and chondro-MSC were stained with the anti-HLA-G antibodies 2A12 (recognizing the sHLA-G5 and HLA-G6 proteins) and 4H84 (recognizing all HLA isoforms) (1:100 dilution; Exbio).
Statistical analysis
Statistical analysis was performed using GraphPad Prism 6.0 (GraphPad Software, Inc.). All data are presented as mean ± SEM. The Mann–Whitney test was used to determine the significance between two groups. The difference was considered as significant when the P value was below 0.05.
Results
Analysis of chondro-MSC-BM/-AT
MSC-BM/-AT were seeded into Alg/HA hydrogel scaffolds to perform chondrocyte differentiation under hypoxia condition in vitro. Cells embedded in hydrogel beads were analyzed by microscopy under × 20 and × 40 magnification (Fig. 1A). To determine the efficiency of chondrogenic differentiation, qRT-PCR was performed to detect the specific cartilage-related gene expression on MSC-BM embedded into Alg/HA hydrogels. From our results, the relative expression of cartilage-related genes, including Sox9, Agg, Col II total, and COMP, was upregulated gradually during the process of chondrogenic differentiation, especially on day 28 (P < 0.05) (Fig. 1B). Besides, to identify chondrocytes derived from MSC-BM/-AT, Alcian blue staining and immunostaining of type II collagen were performed. We show that in contrast to the undifferentiated MSC, chondro-differentiated MSC expressed proteoglycans and type II collagen, as they were positively stained with Alcian blue and collagen II antibody, respectively (Fig. 1C, D).

MSC-BM/-AT differentiated into chondrocytes in vitro.
Chondro-MSC-BM/-AT retained hypoimmunogenicity even after licensing with the inflammatory cytokines IFNγ and TNFα
To determine whether the hypoimmunogenic potential of MSC remained after chondrogenic differentiation, MSC-derived chondrocytes were analyzed for (1) their expression of costimulatory (CD40, CD80, CD86) and HLA class II (HLA-DR) molecules by FACS and (2) their capability of stimulating HLA-mismatched PBMC in MLR. At first, we harvested the MSC-BM embedded in Alg/HA hydrogels on day 7 and 14 during the chondrogenic differentiation and evaluated their surface markers. Our data revealed that the percentage of cells positively expressed CD40, CD80, and CD86 and changed differently during the process of chondrogenic differentiation, while HLA-DR was always absent on chondro-MSC-BM (Fig. 2). When cells were treated with IFNγ and TNFα, the expression of surface markers CD40, CD80, and CD86 showed change, while it was not the same case in HLA-DR expression, as shown in Fig. 2.

Expression of costimulatory molecules and HLA-DR on MSC-derived chondrocytes. The expression of costimulatory factors, including CD40, CD80 (B7-1), and CD86 (B7-2), as well as HLA-DR, was tested on MSC-BM during the process of chondrogenic differentiation on day 7 and 14. Cells pretreated with IFNγ and TNFα (with stimulation) or not (without stimulation) were also considered. Numbers on the right of each graph represent the percentage of positive cells (top) and the MFI calculated on the positive cells compared to the respective isotype control (bottom). White curves indicated cells stained with isotype control and black curves indicated cells stained with the corresponding antibody. HLA, human leukocyte antigen; IFNγ, interferon γ; MFI, mean fluorescence intensity; TNFα, tumor necrosis factor α.
Since the expression level of HLA-DR may reveal the efficiency of IFNγ and TNFα treatment, we tested its expression on MSC-BM/-AT and chondro-MSC-BM/-AT pretreated with IFNγ and TNFα or not. Data showed that without treatment MSC-BM/-AT and chondro-MSC-BM/-AT did not express HLA-DR. When prelicensing with IFNγ and TNFα, the expression level of HLA-DR on MSC-BM/-AT (Fig. 3A, B) was upregulated, especially on MSC-BM. By contrast, no change of HLA-DR expression was observed on chondro-MSC-BM/-AT (Fig. 3C, D). To further explore their immunogenicity, MSC-BM/-AT and chondro-MSC-BM/-AT pretreated with IFNγ and TNFα or not were used as stimulating cells in allogenic settings facing HLA-mismatched PBMC used as responding cells. The MLR based on the use of HLA class II+ LCL stimulating cells facing the HLA-mismatched PBMC constituted the positive control. MSC or MSC-derived chondrocytes were diluted in a series of concentrations (5 × 104, 2.5 × 104, 1.25 × 104, 6 × 103, 3 × 103) and thus used at various responders: stimulator ratios when facing allogenic PBMC. Our results indicated that no PBMC alloproliferation was observed in response to various doses of MSC-BM/-AT even after licensing with inflammatory cytokines (Fig. 3E, F). Also, this pattern was mirrored in chondro-MSC-BM/-AT (Fig. 3G, H). The mean alloproliferation percentage (obtained from three distinct healthy donors) toward MSC-BM or chondro-MSC-BM and MSC-AT or chondro-MSC-AT is shown in Tables 1 and 2, respectively. It is of note that chondro-MSC-BM prelicensing with IFNγ and TNFα slightly increased their immunogenicity when they were at lower doses (6 × 103 and 3 × 103; n = 3, P < 0.05, Table 1).

MSC-BM/-AT and chondro-MSC-BM/-AT showed hypoimmunogenicity even primed by IFNγ and TNFα. Expression of HLA-DR was evaluated by flow cytometry on MSC-BM/-AT
PBMC from healthy individuals were used as responder cells toward MSC-BM and chondro-differentiated MSC-BM, primed with IFNγ and TNFα or not, as stimulating cells at five different responder:stimulator (R:S) ratios.
Data are mean ± SEM of alloproliferation percentage obtained with three distinct donors. This percentage was calculated considering PBMC stimulated by irradiated LCL as 100% alloproliferation.
BM, bone marrow; IFNγ, interferon γ; LCL, lymphoblastoid cell line; MSC, mesenchymal stem cells; PBMCs, peripheral blood mononuclear cells; TNFα, tumor necrosis factor α.
PBMC from healthy individuals were used as responder cells toward MSC-AT and chondro-differentiated MSC-AT, primed with IFNγ and TNFα or not, as stimulating cells at five different responder:stimulator (R:S) ratios.
Data are mean ± SEM of alloproliferation percentage obtained with three distinct donors. This percentage was calculated considering PBMC stimulated by irradiated LCL as 100% alloproliferation.
AT, adipose tissue.
MSC-derived chondrocytes display immunosuppressive properties on T-cell alloproliferation as well as on NK cell-mediated cytotoxicity
To assess whether the immunosuppressive capacity of MSC-BM/-AT changed after chondrogenic differentiation, we first performed MLR using MSC-BM/-AT or chondro-MSC-BM/-AT as third-party cells facing HLA-mismatched PBMC. LCL was served as stimulating cells to induce T-cell alloproliferation. MSC-BM/-AT and chondro-MSC-BM/-AT pretreated with IFNγ and TNFα or not were prepared at five different concentrations to evaluate the immunosuppressive threshold in MLR. PBMC obtained from three to five distinct donors were used. Our data revealed that both MSC-BM (Fig. 4A) and MSC-AT (Fig. 4B) elicited potent immunosuppression on allogeneic PBMC. Also, this inhibition was MSC dose dependent, declining with the lower dosages of MSC. In addition, when MSC-BM/-AT were at lower concentrations, T-cell alloproliferation inhibition significantly dropped in the IFNγ- and TNFα-treated group (Fig. 4A, B), indicating that the inflammatory cytokines weakened the immunosuppressive capacity of MSC-BM/-AT (**P < 0.01, *P < 0.05, n = 3). It is of note that MSC-AT retained their regulation of immune response after chondrocyte differentiation. While chondro-MSC-BM induced T-cell alloproliferation inhibition at high doses but not at low dosages (0.06 × 105 and 0.03 × 105, Fig. 4C, D), indicating that chondro-MSC-BM may have lost their immunomodulatory capacity during chondrocyte differentiation. In addition, the weakened immunosuppressive capacity caused by cytokine pretreatment was not found with chondro-MSC-BM/-AT.
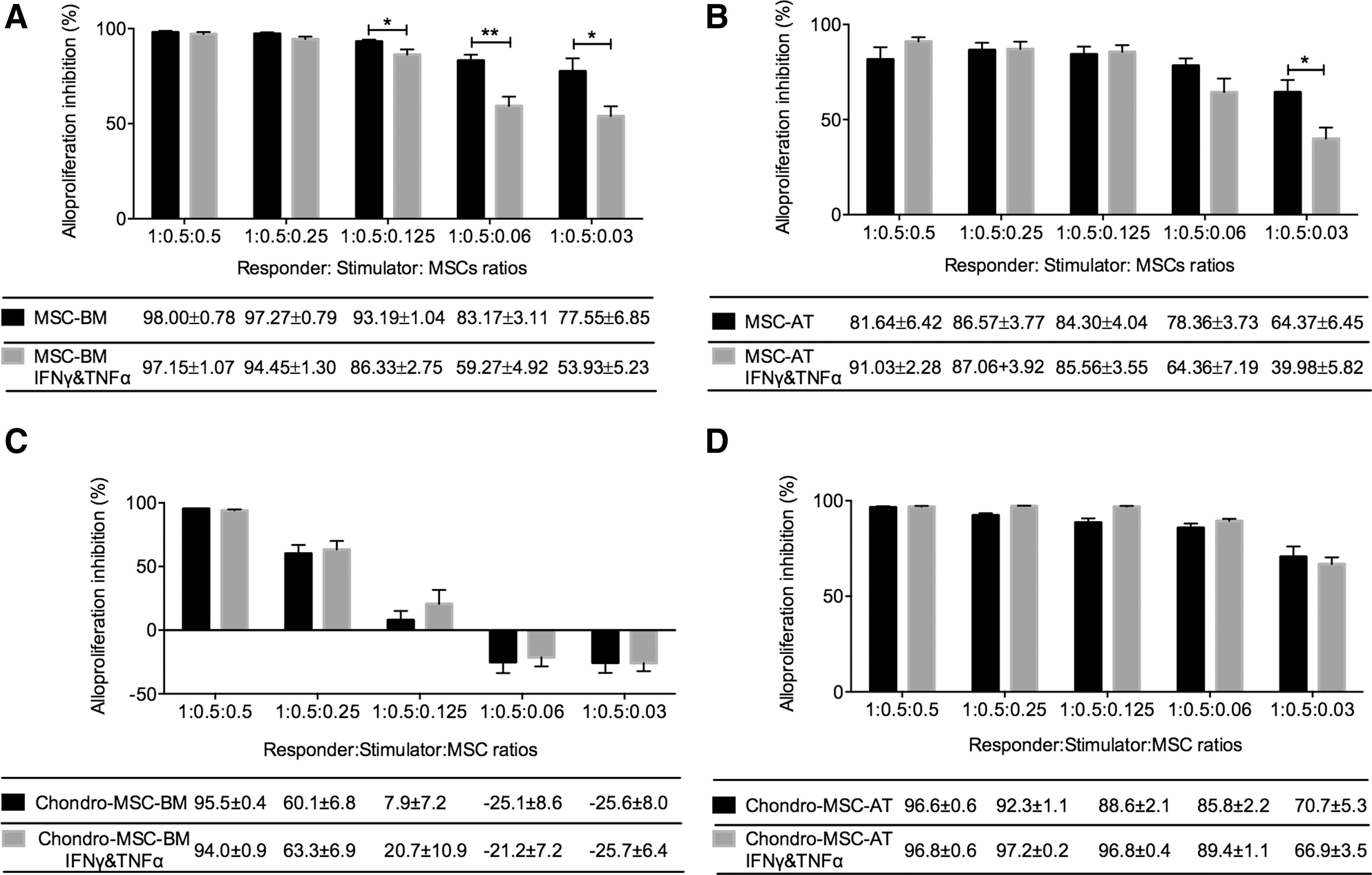
MSC-BM/-AT and chondro-MSC-BM/-AT exerted immunosuppression on HLA-mismatched PBMC in a dose-dependent manner. Irradiated LCL (stimulator) was used toward PBMC (responding cells) to induce potent T-cell alloproliferation, considered as 100% alloproliferation. As third-party cells, MSC-BM/-AT
To further investigate the effect of MSC-derived chondrocytes on NK cell function, cytotoxicity assays were performed in which MSC-derived chondrocytes served as third-party cells in NK cell (effector cell)-mediated cytolysis against K562 target cells (Fig. 5). Results showed that MSC-derived chondrocytes inhibited the cytolytic function of different donor-derived NK cells at all concentrations tested (1.5 × 104, 7.5 × 103, 3.75 × 103, 1.75 × 103, and 0.9 × 103). When considering the K562 lysis by NK cells as 100% specific cytolysis, the inhibition exerted by MSC-derived chondrocytes was between 30% and 75% according to the ratio used. As previously shown by Selmani et al. [16], MSC-BM display strong inhibition on NK cell function. Our data confirmed that MSC-derived chondrocytes also retained potent suppression on NK cell-mediated cytotoxicity.
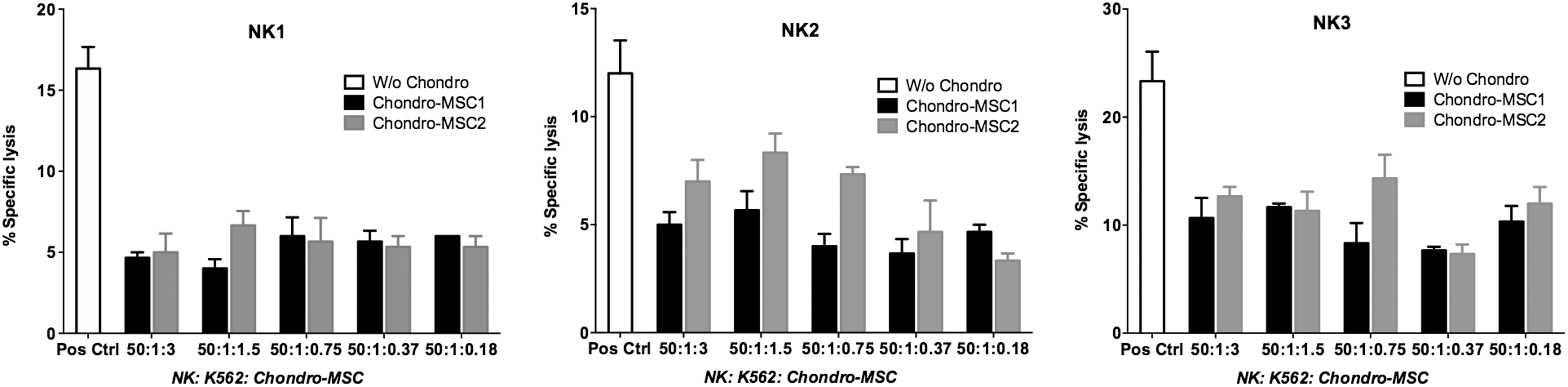
MSC-derived chondrocytes strongly inhibited the NK cell-mediated cytotoxicity. To define the immunosuppressive property of MSC-derived chondrocytes on NK cell-mediated cytolysis, cytotoxicity assay was performed. The cytolysis activity of peripheral blood NK cells against the K562 target cells was assessed in 4-h 51Cr-release assay. NK cells were obtained from three distinct healthy donors, and MSC-derived chondrocytes from two distinct BM samples were used. “W/o chondro” indicated that NK effector cells were used against K562 target cells in absence of MSC-derived chondrocytes. Data are presented as mean ± SEM; each sample was performed in triplicate. NK, natural killer.
Alg/HA hydrogel scaffold did not affect both hypoimmunogenic and immunosuppressive capacity of MSC-BM/-AT
As it mimics the natural extracellular matrix, Alg/HA hydrogel scaffold is a promising biomaterial for tissue engineering [22]. However, it is necessary to fully investigate the properties of Alg/HA hydrogel scaffold whether it is antigenic or toxic in terms of triggering the immunological responses. In this regard, we combined MSC-BM/-AT with hydrogel scaffold as one unity to be used as stimulating cells or third-party cells in MLR. MSC-BM/-AT without hydrogel were used as control. PBMC obtained from three healthy donors were used. Each PBMC was performed in triplicate in allogenic settings with or without hydrogel. Our results showed that both MSC-BM (Fig. 6A) and MSC-AT (Fig. 6B), combined with hydrogel or not, did not elicit T-cell alloproliferation. One representative experiment is shown (Fig. 6). Pretreatment with IFNγ and TNFα significantly enhanced the hypoimmunogenic properties of MSC-BM and MSC-AT regardless of the presence of hydrogel (P < 0.001, Table 3). Moreover, addition of hydrogel scaffold did not change the immunosuppressive potential of MSC-BM (Fig. 6C) and MSC-AT (Fig. 6D) in allogenic settings. Noticeably, IFNγ- and TNFα-primed MSC-BM/-AT showed significantly enhanced immunosuppressive capacities even though at lower cell dosages (***P < 0.001, **P < 0.01,*P < 0.05, n = 3; Fig. 6C, D). These results differ from the data in Figure 4A and B. These varying effects of cytokines were also seen in our previous studies [20,23], leading us to propose that the immune regulation mediated by MSC may be PBMC donor dependent.
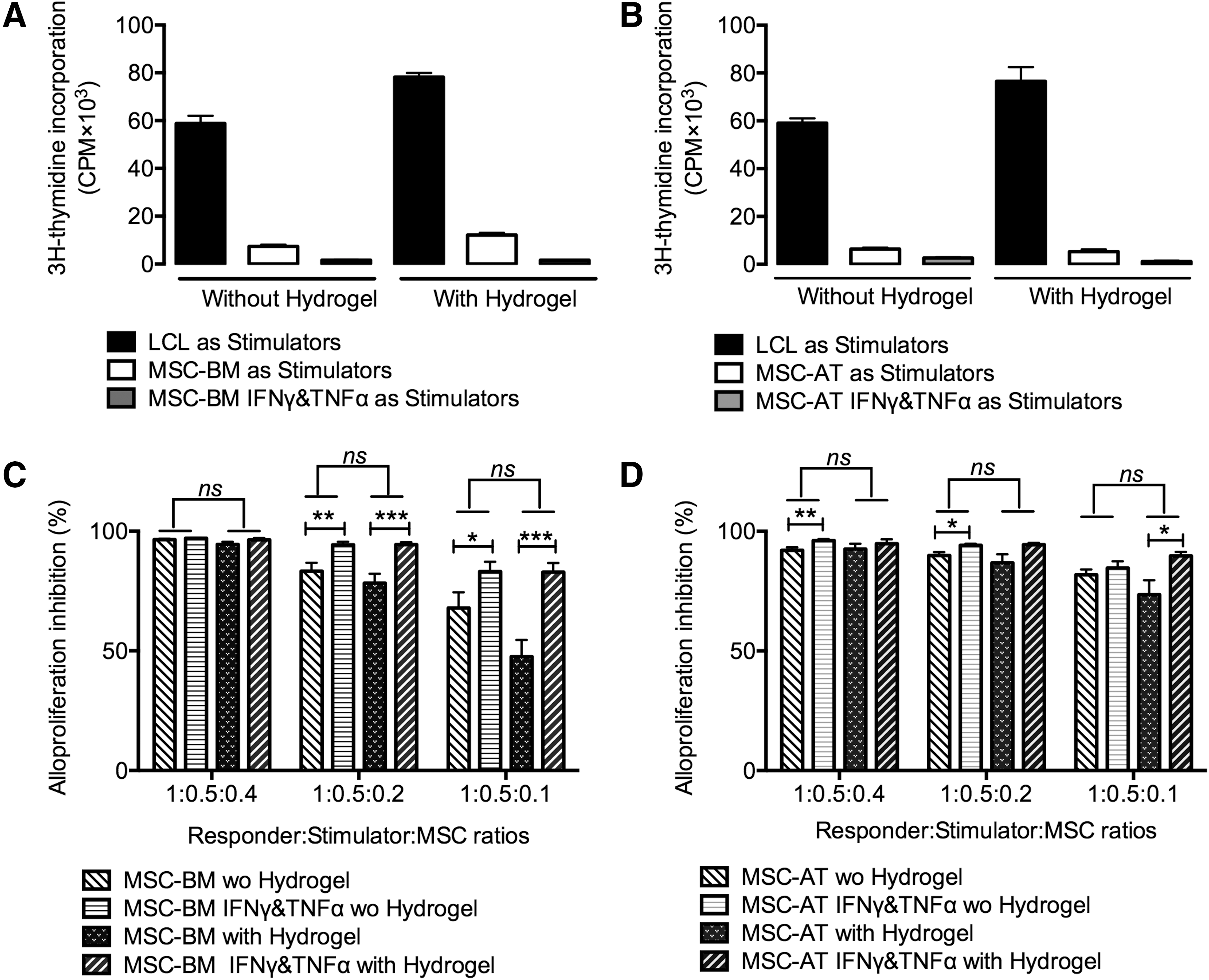
Alg/HA hydrogel scaffold did not affect the hypoimmunogenic and immunosuppressive capacity of MSC-BM/-AT. Combined with Alg/HA hydrogel, MSC-BM/-AT
PBMC from healthy individuals were used as responder cells toward MSC-BM/-AT, primed with IFNγ and TNFα or not, as stimulating cells at 1:0.2 responder:stimulator ratio.
Data are mean ± SEM of alloproliferation percentage obtained with three distinct donors. This percentage was calculated considering PBMC stimulated by irradiated LCL as 100% alloproliferation.
MSC-BM/-AT and chondro-MSC-BM/-AT express HLA-G
To evaluate the expression level of HLA-G on MSC, Passages P3 and P6 of MSC-BM/-AT were tested by flow cytometry. Our data show that the expression of HLA-G on MSC-BM/-AT decreased as cells repeatedly passaged (Fig. 7A). Then, MSC pretreated with IFNγ and TNFα or not were studied to investigate the effect of cytokines on HLA-G expression. We observed a significant increase of HLA-G expression on MSC-BM/-AT after stimulating with inflammatory cytokines (Fig. 7B). To study the effect of chondrocyte differentiation process on HLA-G expression, chondro-MSC-BM/-AT slides were stained with the anti-HLA-G antibodies 4H84 and 2A12. Confocal data revealed that a significantly increased expression of HLA-G was seen in chondro-MSC-BM/-AT in contrast to undifferentiated MSC-BM/-AT (Fig. 7C). However, the expression of HLA-G on chondro-MSC-BM/-AT decreased after incubation in normoxia for 3 days, and this expression did not change under cytokine stimulation compared with the undifferentiated MSC-BM/-AT (Fig. 7D).

HLA-G was expressed by MSC-BM/-AT and chondro-MSC-BM/-AT, and its expression level varied with different conditions.
HLA-G5 participated in MSC- and chondro-MSC-mediated immunosuppression, and its expression level was boosted in immune-active environment
To study the role of HLA-G played in MSC-mediated immunosuppression, we collected the MSC-BM/-AT that participated in MLR or not, made cell slides, and stained them with the anti-HLA-G5 2A12 mAb to evaluate HLA-G expression (Fig. 8A). Interestingly, compared with the unstimulated MSC-BM/-AT, we found an upregulation of HLA-G expression in MSC-BM/-AT that have been involved in MLR. In addition, our confocal microscopy analysis revealed that not all the MSC-BM/-AT expressed HLA-G, indicating the existence of different functional MSC subpopulations. Due to the cell mixture of MSC and PBMC after MLR, to precisely determine the expression of HLA-G on MSC, we harvested the cells, double-stained them with anti-HLA-G and anti-CD14 antibodies, and then analyzed them by flow cytometry. Based on the cell size forward scatter (FSC), we excluded lymphocyte populations (smaller size, smaller FSC). Then, we gated the MSC population that was CD14negative (to exclude the monocyte population) and analyzed their HLA-G expression. First, we stained cells with MEMG9 mAb (recognizing membrane-bound HLA-G proteins) without carrying out permeabilization with saponin. Data revealed that almost no MEMG9-positive cells could be seen (Fig. 8B), indicating that membrane-bound HLA-G might not participate in MSC-mediated immunosuppression. Then, we used saponin to destroy the cell membrane and labeled cells with the anti-HLA-G5 2A12 mAb. Surprisingly, a boost of HLA-G was observed in immune-activated MSC-BM/-AT (Fig. 8C, D), suggesting that HLA-G5 was the main HLA-G isoform that took part in MSC-mediated immunomodulation. Interestingly, the expression of HLA-G was varying in the three donor-derived PBMC settings. Also, not all the MSC expressed HLA-G, which was consistent with our confocal data. Our results indicated that MSC displayed heterogeneity consisting of different functional subpopulations. Similar results were found with chondro-MSC-BM/-AT (Fig. 8E, F). Besides that, we primed chondro-MSC-BM with cytokines before MLR. Interestingly, there was a slight raise of HLA-G (from 64.3% to 72.0%) in chondro-MSC-BM compared to unprimed chondro-MSC-BM (Fig. 8F), indicating that the regulatory capacity of chondro-MSC-BM could be enhanced by pretreatment with inflammatory cytokines.
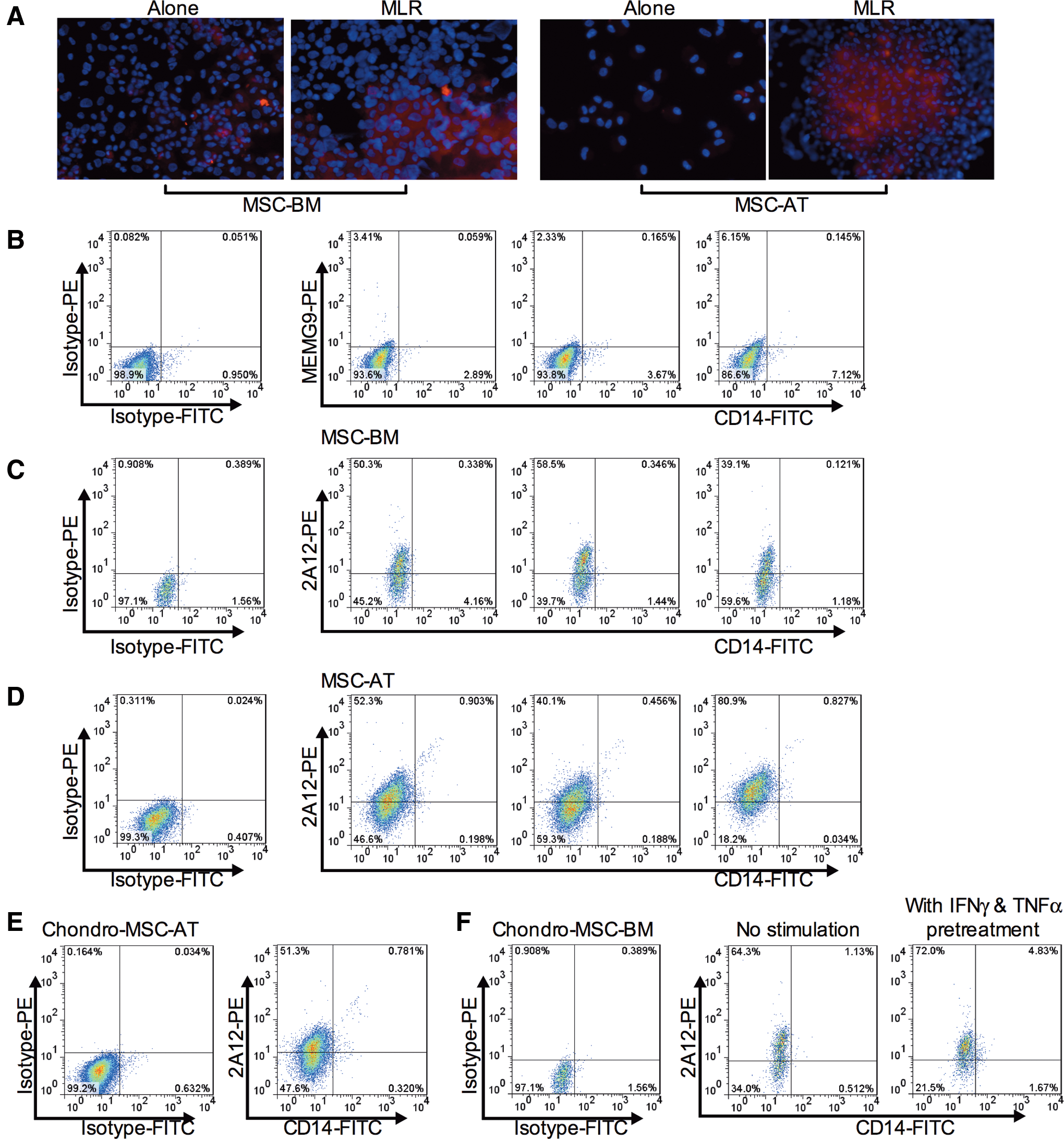
Expression level of HLA-G5 in MSC and chondro-MSC was boosted in immune-active environment.
To further determine the role played by sHLA-G5 in MSC-BM/-AT- or chondro-MSC-BM/-AT-mediated immunosuppression on HLA-mismatched PBMC assays, sHLA-G5 was neutralized by using the 87G anti-HLA-G5 blocking antibody. Also, we analyzed MSC or MSC-derived chondrocytes pretreated with cytokines. Our results showed that MSC-BM- and MSC-AT-mediated inhibition of T-cell alloproliferation was significantly reversed by adding 87G to MLR (P < 0.05, Fig. 9A, B). However, when MSC were primed with IFNγ and TNFα, regardless of BM or AT derived, the restoration by 87G was not significant (P > 0.05, Fig. 9A, B). Also, MSC-derived chondrocyte-mediated immunosuppression of T-cell alloproliferation was only partially reversed by 87G (P > 0.05, Fig. 9C, D). These data show that sHLA-G5 is largely responsible for undifferentiated MSC-mediated immunosuppressive properties; however, its role in immune suppression mediated by MSC, once primed with cytokines or under chondrogenic differentiation, is not prominent.

Anti-HLA-G antibody partially restored T-cell alloproliferation by neutralizing the secreted soluble HLA-G5 from MSC-BM/-AT and chondro-MSC-BM/-AT. 20 μg/mL 87G (anti-HLA-G antibody) and isotype control antibody were, respectively, added to MLR, in which MSC-BM/-AT
Discussion
In the present study, we addressed four main points relying on whether immune regulatory properties of MSCs were modified by (1) MSC differentiation toward the chondrocytic cell lineage, (2) addition of natural scaffold (ie, Alg/HA hydrogel), (3) licensing with inflammatory cytokines IFNγ and TNFα, and (4) involvement of the tolerogenic HLA-G molecule. In this regard, we analyzed both the immunogenic and immunosuppressive properties of MSC from BM or AT in allogenic conditions, that is, facing HLA-mismatched PBMC. To identify low immunogenic MSC types, we studied their ability to be recognized as allogenic cells by HLA-mismatched PBMC in MLR using MSC as stimulating cells and PBMC from various healthy donors as responder cells. To examine their immunosuppressive properties, we studied their ability to modulate T-cell alloproliferation as third-party cells in a classical MLR. All the functional experiments were performed by considering the differentiation status of MSC either immature or chondro-differentiated, seeded onto biomaterial or not, and pretreated by IFNγ and TNFα or not.
Although the cell pellet method is a common way to perform MSC chondrogenic differentiation in vitro, it remains difficult to harvest individual cells after cell differentiation. Thus, fewer studies investigated the individual character of MSC after chondrogenic differentiation. With promising hydrogel scaffold appearance for chondrocytic differentiation [22,24], MSC seeded into the crosslinked hydrogel were easily isolated [21] and individual chondro-MSC were available for further analysis. Alg/HA hydrogel scaffold is a hydrophilic polymer network with a porous microstructure character allowing nutrient diffusion and biological activities (proliferation, differentiation, cell–cell contact, and so on) of cells encapsulated. Reppel et al. reported that BM- and umbilical cord-derived MSC embedded into Alg/HA hydrogel were able to differentiate into chondrocytes in vitro by verifying their expression of specific cartilage-related genes at the mRNA and protein level [24]. Following this protocol, we also successfully induced MSC-BM and MSC-AT encapsulated in Alg/HA hydrogel into chondrocytes. Of note, Alg/HA hydrogel is not only an attractive biomaterial for CTE, but also a promising hydrogel candidate for bone [25] and myocardial tissue engineering [26]. Moreover, Alg/HA hydrogel has also been reported to accelerate the in vivo wound healing process on rat model of excised wound [27]. As supplement, our data revealed that Alg/HA hydrogel was hypoimmunogenic and did not initiate immune response in allogenic settings in vitro even under inflammatory conditions, indicating its biocompatible property that would support its clinical application.
We reported that MSC-BM- and MSC-AT-derived chondrocytes retained their low immunogenicity and immunosuppressive function (higher cell dosages) even primed with cytokines. Although the expressions of costimulatory molecules CD40, CD80, and CD86 on MSC-derived chondrocytes were upregulated in the process of chondrogenic differentiation, it did not functionally trigger T-lymphocyte activation even when they were pretreated with IFNγ and TNFα, probably due to the fact that the HLA class II molecules (HLA-DR) were not expressed. With comparison, a much higher expression level of costimulatory factors was found on monocyte-derived mature dendritic cells (mDC), which were generated in the presence of CTLA4-Ig, and those mDC were proved to be hypoimmunogenic since they were unable to challenge alloreactive T cells [28]. Besides, our results are different from the previous studies: Chen et al. reported that the chondrogenic differentiation would alter the immunosuppressive properties of mouse BM-MSC by upregulating the expression of the costimulation molecule B7 [29]. Technau et al. demonstrated that chondro-MSC-AT reserved immunosuppressive properties but developed immunogenicity [30]. Why MSC developed distinct immune outcomes during chondrogenic differentiation? We believe that cell species, tissue sources, induction medium, and culture conditions synergistically drive the fate of chondro-MSC. Among involved factors, both Chen and Technau studies used TGF-β to induce MSC chondrogenic differentiation. TGF-β was reported not only as a mediator of MSC chondrogenesis but also as increasing the immunogenicity of MSC via upregulation of the expression of HLA-DR [30] and B7 [29]. By contrast, here we adopted the Alg/HA hydrogel method to induce MSC chondrogenic differentiation without adding growth factor, especially TGF-β. Considering this difference, we propose that adjusting the induction medium may be an approach to get immune privileged chondrocytes for clinical application. In addition, we reported that MSC-derived chondrocytes maintained potent immunosuppressive capacities on activated PBMC as well as on NK effector cells, which probably related to their expression of HLA-G [16,31,32], indicating that allogeneic MSC-derived chondrocytes would be promising candidates for substitute of autologous chondrocytes in cartilage tissue regeneration. Meanwhile, our data show that chondro-MSC-AT but not chondro-MSC-BM inhibited strongly allostimulated PBMC even at lower dosages, suggesting that MSC derived from AT would be better than those derived from BM for cartilage repair.
HLA-G has been reported to be expressed by MSC derived from various tissues, including BM [16], AT [20], umbilical cord [33], and placenta chorionic villi [34]. Moreover, HLA-G was expressed by osteo-differentiated MSC at the protein level [20] and chondro-differentiated MSC-AT at the mRNA level [30]. Our study further demonstrated its raised expression on chondro-MSC at protein level via immunostaining with the anti-HLA-G5 2A12 mAb and the pan HLA-G 4H84 mAb. However, the expression of HLA-G on chondro-MSC apparently decreased after several days in normoxia cultures, indicating that hypoxia was a critical factor that affected the HLA-G output on MSC, in agreement with Reppel's report [35]. Meanwhile, we detected an increased restoration of PBMC alloproliferation when anti-HLA-G antibody 87G was added to the allogenic settings. Based on these data, we hypothesized that a boost expression of HLA-G occurred on MSC during MLR. To verify this point, we collected and immunostained the MSC that participated in MLR with the anti-HLA-G5 2A12 mAb. As we have expected, cells were strongly stained with 2A12 under confocal microscopy analysis. Since the cells after MLR contained monocytes, lymphocytes, and MSC, we targeted HLA-G staining on MSC by excluding CD14+ monocytic cells in FACS analysis. Our data revealed that sHLA-G5 (2A12, saponin permeabilization) rather than membrane-bound HLA-G1 (MEMG-9, without saponin permeabilization) was significantly increased in immune active MSC or chondro-MSC. The secreted HLA-G5 from MSC/chondro-MSC induced immune inhibition on activated PMBC that was partially blocked by adding its specific neutralizing antibody 87G. These results pointed out that secretion of HLA-G5 was an important mechanism in MSC- or chondro-MSC-mediated immunosuppression, which was consistent with Selmani's study [16]. Interestingly, we also observed that MSC-mediated immunosuppression was PBMC dependent, and the raised expression of HLA-G on MSC varied when facing different PBMC donors. Of note, our confocal results show that the MSC population revealed functional heterogeneity regarding HLA-G expression, especially when facing activated PBMC. Thus, we hypothesized that HLA-G+ MSC subsets would be more favorable for clinical immunosuppressant biotherapy [17,36]. Besides that, a decreased expression of HLA-G on MSC with passages in vitro was observed during our study, and MSC-mediated immune inhibition weakened when MSC were at higher cell passages (data not shown) [17]. Based on our data, we proposed that HLA-G could be an indicator for MSC aging, which was in accordance with the study of Rizzo et al. [37], who demonstrated a decreased production of sHLA-G from MSC with in vitro passage. Besides, Rizzo et al. also confirmed that the loss of MSC inhibitory functions was closely related to MSC aging by using a longitudinal analysis.
Considering all these data, early passage MSC should be preselected for clinical individualized cell immunotherapy.
Although here we demonstrated that MSC-derived chondrocytes displayed immunosuppression on PBMC alloproliferation as well as on NK cell-mediated cytotoxicity in vitro, it will be necessary in future studies to analyze their in vivo fate, especially facing B lymphocytes and dendritic cells. In this regard, Huang et al. demonstrated that both systemic and local injection of allogeneic MSCs was effective to promote fracture healing; however, only less than 3% injected Luc-MSC remained after 5 weeks postinjection [38]. Similarly, van Velthoven et al. demonstrated that the majority of transplanted MSC did not survive up to 3 days after transplantation [39]; less than 1% allogeneic MSC survived up to 4 weeks after transplantation, no matter what the administration routs [40 –42], even though transplanted cells were able to be detected up to 6 weeks after transplantation [40,42]. Referring to these studies, we hypothesize that the transplanted MSC or MSC-derived chondrocytes may temporarily exist at the injury tissue/organ but will produce a significant therapeutic effect depending on their paracrine and differentiation capabilities.
In summary, we identified both low immunogenicity and strong immunosuppressive function of MSC-BM/-AT after chondrocyte differentiation. Moreover, we showed that HLA-G5 participated in MSC-derived chondrocyte-mediated inhibition. In addition, Alg/HA hydrogel scaffold was hypoimmunogenic and suitable for transplantation in vivo, even in combination with MSC. Based on our study, MSC seeded into biocompatible Alg/HA hydrogel scaffold may be a promising treatment for cartilage tissue repair.
Footnotes
Acknowledgments
We thank Roman Luinaud and Florent Montespan for their help on immunological assays and Marina Daouya and Qing-Lian Wu for their help on flow cytometry analysis. This work was supported by the Chinese Scholarship Council (CSC).
Author Disclosure Statement
No competing financial interests exist.