
Abstract
Diabetic retinopathy (DR) is a hyperglycemia (HG)-mediated microvascular complication. In DR, the loss of pericytes and subsequently endothelial cells leads to pathologic angiogenesis in retina. Adipose-derived stromal cells (ASC) are a promising source of therapeutic cells to replace lost pericytes in DR. To date, knowledge of the influence of HG on the bioenergetics and pericytic function of ASC is negligible. Human ASC were cultured in normoglycemia medium (5 mM
Introduction
T
In this study, we investigated the influence of HG on the mitochondrial function and metabolic activity of ASC and on their pericytic function in vitro. This information may help to delineate both their capabilities and limitations with respect to their potential clinical translation.
Material and Methods
Cell isolation and culture
Human subcutaneous adipose tissue samples from healthy human subjects with body mass index <30 were obtained after liposuction surgery (Bergman Clinics). All donors provided informed consent and all procedures were performed in accordance to national and institutional guidelines as well as with the ethical rules for human experimentation stated in the Declaration of Helsinki.
For ASC isolation, lipoaspirates were enzymatically digested with 0.1% collagenase A (Roche Diagnostic) in phosphate-buffered saline (PBS), containing 1% bovine serum albumin (Sigma-Aldrich) at 37°C for 90 mins. Centrifugation (300 g, 4°C, 10 mins) was used to separate adipocytes and lipid content from the stromal cell fraction. The stromal cell fraction was subjected to Lymphoprep (Axis-Shield PoC) density gradient centrifugation. The cells from the interface were seeded in culture flasks at 10,000 cells/cm2. The culture medium was normoglycemic RPMI-1640 (5 mM D-glucose, NG-RPMI) (Lonza Biowhittaker Verviers) supplemented with 10% fetal bovine serum (FBS) (Thermo Scientific), 100 U/mL penicillin, 100 μg/mL streptomycin (Gibco, Life Technologies), and 2 mM L-glutamine (Lonza Biowhittaker Verviers). It was sterilized by filtration (0.22 μm). Optimal culture conditions of 37°C, 5% CO2, and 95% humidity were maintained throughout the culture period. ASC in passage 1–3 were harvested with Trypsin-EDTA in 0.9% NaCl (Sigma-Aldrich) and routinely checked for both mesenchymal cell surface and pericyte markers, including CD31−/CD44+/CD45−/CD29+/CD105+/CD144−/NG2+/PDGFRβ+ [19]. The cells were passaged at 70%–80% confluence until passage3 (P3) with three changes of medium per week. The cells from P3–6 were cultured in normoglycemic and hyperglycemic (30 mM
Immunofluorescence analysis of cocultured ASC and HUVEC
ASC were plated in 24-well cell culture plates (Corning) at 10,000 cells/cm2 in NG-RPMI medium for 5 days. HUVEC were seeded on top of ASC monolayers or as the control on gelatin-coated wells at 10,000 cells/cm2 in NG-ECM or HG-ECM for 7 more days after which vascular networks had formed. Cells were washed with PBS and fixed in 2% paraformaldehyde in PBS at RT for 20 min. Cells were permeabilized with 0.5% Triton X-100 in PBS (Sigma-Aldrich) for 15 min. Subsequently, samples were incubated with goat-anti-human-RGS5 antibody (1:100; Santa Cruz)/rabbit anti-human-SM22α antibody (1:100; Abcam) and mouse-anti-human-CD31 antibody (1:100; Dako) for 90 min. Samples were washed with PBS and incubated with a cyanine3-conjugated-rabbit-antibody to goat-IgG for RGS5/cyanine3-conjugated-donkey-antibody to rabbit-IgG for SM22a (1:300; Life Technologies) and fluorescein-conjugated-donkey-antibody to mouse-IgG (for CD31, 1:300; Life Technologies) for 45 min. Stained samples were mounted with PBS. Imaging was performed with a high-end fully motorized Zeiss AxioObserver Z1 epifluorescent microscope (TissueGnosticsTissue Faxs). The coculture experiment was repeated with HUVEC that were lentivirally tagged to express EGFP (green) and ASC lentivirally tagged with dTomato (red) to monitor the origin cells irrespective of phenotypes changes during coculture.
Apoptosis and proliferation
To estimate the influence of HG on cell survival, apoptosis was evaluated by fluorescence-activated cell sorting (FACS) analysis using an Apoptotic/Necrotic Cells Detection Kit (PromoKine). Briefly, ASC were treated with NG-RPMI, HG-RPMI, and NG+Ma-RPMI medium for 7 days and harvested by Trypsin-EDTA in 0.9% NaCl right after treatment. Cells were washed with PBS once and resuspended in 500 μL kit binding buffer (100,000 cells/500 μL) and stained in the dark with 2.5 μL FITC-AnnexinV (marker for apoptosis) and 2.5 μL of Ethidium Homodimer lll (marker for necrosis) at RT for 15 min. Samples were analyzed using a FACS Calibur flow-cytometer (BD biosciences) within 1 h after staining and the results were analyzed with Kaluza 1.2 software. For measurement of proliferation, ASC from P3-6 were plated on 24-well cell culture plates (Corning) at 10,000 cells/cm2 in NG-RPMI medium with 10% FBS for 24 h. The medium was changed for NG-RPMI, HG-RPMI, or osmotic control, all with 5% FBS for 7 days. Medium was changed daily. Cells were fixed and permeabilized as described above. Samples were incubated with rabbit-anti-human-ki67 (Monosan PSX1028) 1:250 in DAPI (5 mg/mL; Sigma-Aldrich) for 90 min. Samples were washed and subsequently stained by incubation with a cyanine3-conjugated-donkey-antibody to rabbit-IgG (1:100; Life Technologies) for 30 min. Stained samples were mounted with PBS. Imaging was performed with a high-end fully motorized Zeiss AxioObserver Z1 epifluorescent microscope.
Detection of ROS level
Cellular ROS production was determined using the dye 2′,7′-dichlorofluorescein diacetate (DCFH-DA; Sigma-Aldrich), a cell permeable nonfluorescent probe, which is de-esterified intracellularly by means of esterases (DCFH) and turns to highly fluorescent 2′,7′-dichlorofluorescein (DCF) on oxidation [21]. Cells were harvested after 7 days culturing in NG, HG, and NG+Ma-RPMI medium and suspended in 1 mL of medium followed by incubation with 20 μM DCFH-DA in the dark at 37°C for 15 min. Samples were analyzed directly without washing using a FACS Calibur flow cytometer (BD Biosciences) within 15 min after the staining and the results were analyzed with Kaluza 1.2 software.
Assessment of mitochondrial membrane potential and mitochondrial morphology analysis
To specify the influence of HG on mitochondrial morphology and membrane potential, ASC cultured in NG-RPMI or HG-RPMI medium for 7 days were incubated at 37°C in a 5% CO2 humidified chamber with 120 nM membrane potential–independent dye MitoTracker Green FM (MTG, Green, Life Technologies) and 10 nM tetramethylrhodamine, ethyl ester, perchlorate (TMRE, Red-orange; Life Technologies), a membrane potential-dependent fluorescent dye for 45 min, washed three times, and incubated in medium containing TMRE for 15 min before imaging to allow adequate equilibration of the membrane potential-sensitive TMRE dye within the mitochondria. TMRE was kept in the medium while imaging. All experiments were performed in triplicate with at least three technical replicates and normalized by the number of cells per sample.
Confocal microscopy
Cells were imaged live by confocal microscope (Leica TCS SP8 Confocal Microscope) with a 63 × oil immersion objective. The cells were kept at 37°C in a 5% CO2 humidified microscope stage chamber. MTG was subjected to 490 nm argon laser excitation and 516 nm filter emission. TMRE was recorded through a band-pass 573–607 nm filter. To observe individual mitochondria Z-stack images were acquired in series of six slices per cell ranging in thickness from 0.5 to 0.8 μm per slice [22].
Analysis of mitochondrial length and circularity
Mitochondrial circularity of ASC was measured as described previously [23]. Mitochondrial circularity is a measure of “roundness” of mitochondria with 0 referring to a straight line and 1 as a perfect circle. Cells containing a majority of long interconnected mitochondrial networks were classified as cells with tubular mitochondria. Cells with a majority of short mitochondria were classified as fragmented and cells with mostly sparse small round mitochondria were classified as very fragmented [24]. Morphological aspects were measured by ImageJ software for individual mitochondria.
Oxygen consumption rate and extracellular acidification rate
The oxygen consumption rate (OCR) and extracellular acidification rate (ECAR) of ASC grown in NG-RPMI or HG-RPMI medium were measured by a bioenergetic assay (XF24; Seahorse Bioscience). ASC were plated and grown on V7-PS plate (Seahorse Bioscience, Inc.) in NG-RPMI or HG-RPMI medium to reach a confluent monolayer for 7 days. Assays were initiated by removing growth medium, replacing with unbuffered RPMI-1640 medium and incubating at 37°C for 60 min in a CO2-free incubator to allow temperature equilibrium and CO2 degassing from plate. The microplate was then assayed (XF24 Extracellular Flux Analyzer; Seahorse Bioscience) to measure extracellular flux changes of oxygen and pH in the medium immediately surrounding the adherent cells. After steady state measurement, oxygen consumption and ECARs were obtained. Oligomycin (2 μM), which inhibits ATP synthase, and the proton ionophore FCCP [carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone; 5 μM], which uncouples mitochondria, were injected sequentially through reagent delivery chambers for each well in the microplate, to obtain maximum OCRs. Finally, a mixture containing 2 μM rotenone (an inhibitor of mitochondrial complex I) and 2 μM antimycin A (an electron transport blocker) was injected to confirm that respiration changes were mainly due to mitochondrial respiration. The values of oxygen consumption and extracellular acidification were normalized to total cellular proteins in each sample well.
Assessment of glucose uptake by 2-deoxy D-glucose
To determine the influence of HG on glucose uptake in ASC, cells from P3–6 were cultured in NG or HG for 7 days. After that cells were washed twice with PBS and serum deprived for 4 h, in either NG or HG medium supplemented with 100 U/mL penicillin, 100 μg/mL streptomycin, and 2 mM
Statistics
All the data are presented as a mean ± SEM and were analyzed by GraphPad Prism (GraphPad Software, Inc.). Statistical significance was determined using one-way ANOVA and unpaired t-test analysis. Values of p < 0.05 were considered statistically significant.
Results
HG reduces viability of ASC, but does not influence proliferation
After 7 days culture in HG medium ∼25% ± 1.1% of the ASC were apoptotic and necrotic, while this was reduced in NG (16% ± 0.8%, P < 0.0001, Fig. 1A, B). Interestingly, the level of proliferation, as determined by the fraction of Ki-67-expressing ASC, did not differ between cells cultured in NG medium versus HG medium (Fig. 1C) or osmotic controls (data not shown) after 7 days culture.
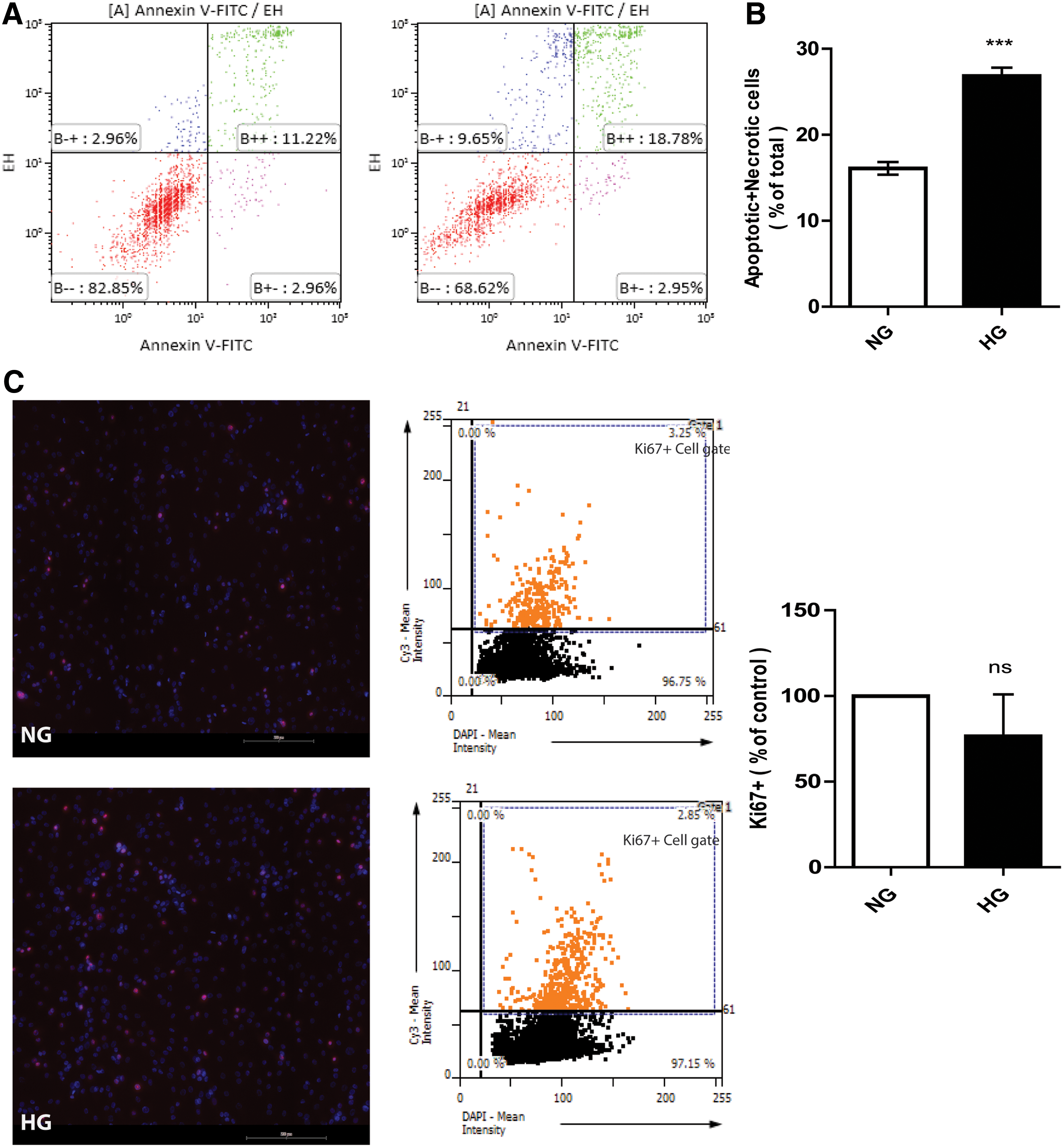
ASC undergo apoptosis after exposure to HG for 7 days. Apoptosis and necrosis were quantified by FACS after FITC-Annexin V and Ethidium Homodimer lll labeling.
HG induces ROS in ASC
ASC cultured in NG or control medium produced virtually no ROS, as determined by FACS analysis of converted DCFH-DA. However, 7 days culture of ASC in HG medium increased intracellular levels of ROS by more than threefold (P < 0.0001, Fig. 2A). In the positive control cells which were treated with 1 mM hydrogen peroxide (Merk) as a potent ROS inducer, ROS induced well beyond the levels induced by HG (Fig. 2B).
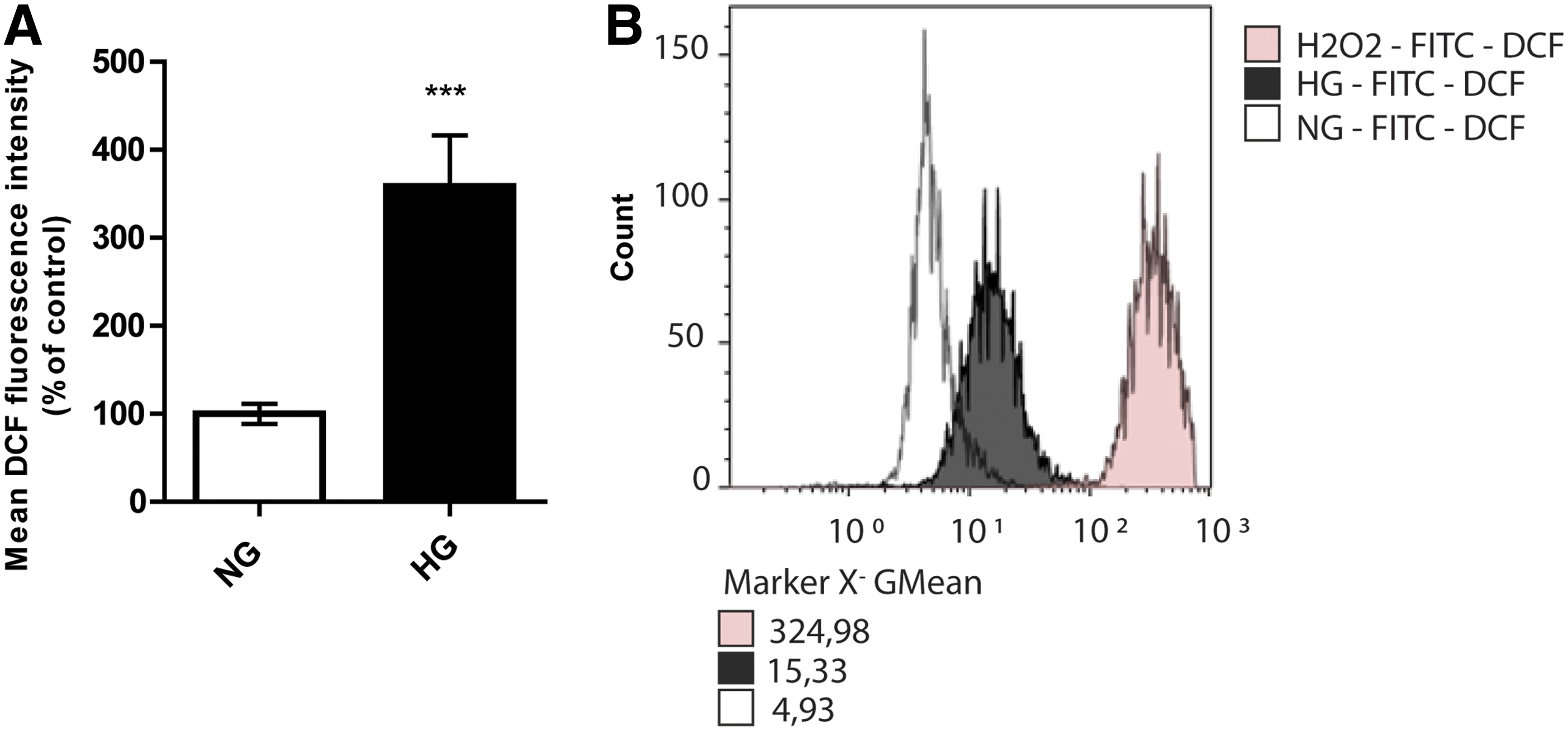
HG induces production of total cellular ROS in ASC. ROS production was measured by conversion of DCF in ASC after exposure to HG for 7 days.
HG alters the mitochondrial membrane potential and changes the mitochondrial phenotype of ASC
The continuous culture of ASC in HG medium decreased their mitochondrial membrane potential by ∼31% ± 8% compared to NG medium as judged by TMRE fluorescence (P < 0.001, Fig. 3). This apparent sign of a decreased mitochondrial membrane potential was corroborated by disturbances in the mitochondrial network of ASC cultured in HG medium for 7 days and assessed with MitoTracker dye. This showed a significant decrease of 23% ± 9.6% compared to the NG control. ASC cultured in NG medium had an extensive network of mitochondria throughout the cell (P < 0.01, Fig. 4A). Another relevant feature to determine mitochondrial dysfunction is mitochondrial shape and circularity. We observed a mainly tubular mitochondrial morphology in the ASC cultured in NG medium, which changed to a fragmented morphology when cultured in HG medium for 7 days. The mitochondrial fragmentation increased to virtually small fragments in nonphysiologically high (50 mM) glucose concentrations (P < 0.0001, Fig. 4B).

HG effected mitochondrial membrane potential of ASC.
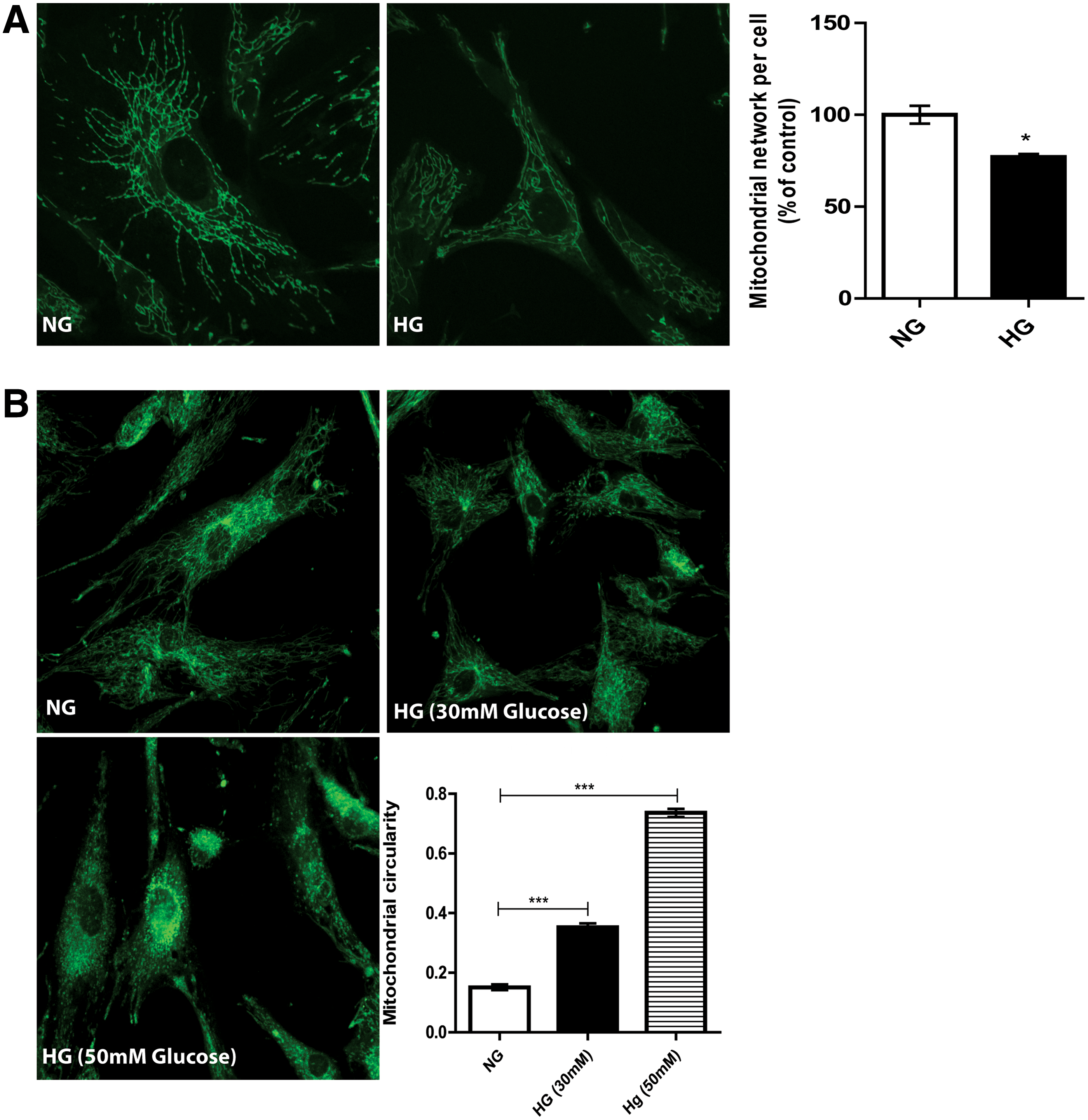
Mitochondrial morphology of ASC changes after exposure to HG.
HG decreases the maximum OCR and the ECAR in ASC
ASC cultured in NG or HG conditions were simultaneously measured with a bioenergetic assay (XF24; Seahorse Bioscience) to determine the rates of cellular oxygen consumption and extracellular acidification. Steady state of oxygen consumption and extracellular acidification were measured at three time points (Fig. 5). Oligomycin (Fig. 5A: injection vertical line A) was injected to inhibit ATP synthase, followed by the addition of FCCP (Fig. 5A: injection vertical line B) to uncouple mitochondria and obtain values for maximum oxygen consumption. Finally, mixture of rotenone and antimycin A were injected (Fig. 5A: injection vertical line C) to confirm that the respiration changes just could be attributed to mitochondrial respiration. ASC grown in HG showed a significant decrease (29.54% ± 3%) in maximum oxygen consumption compared to ASC grown in NG medium (P < 0.0001, Fig. 5B). ECARs were examined simultaneously. Changes in the ECAR may indicate changes in the rate of glycolysis in these cells. Under HG, ASC showed significantly decreased extracellular acidification (Basal: 10%, oligomycin-induced: 34%, and FCCP-induced: 44%) as compared with ASC grown in NG medium (P < 0.01, Fig. 5C).
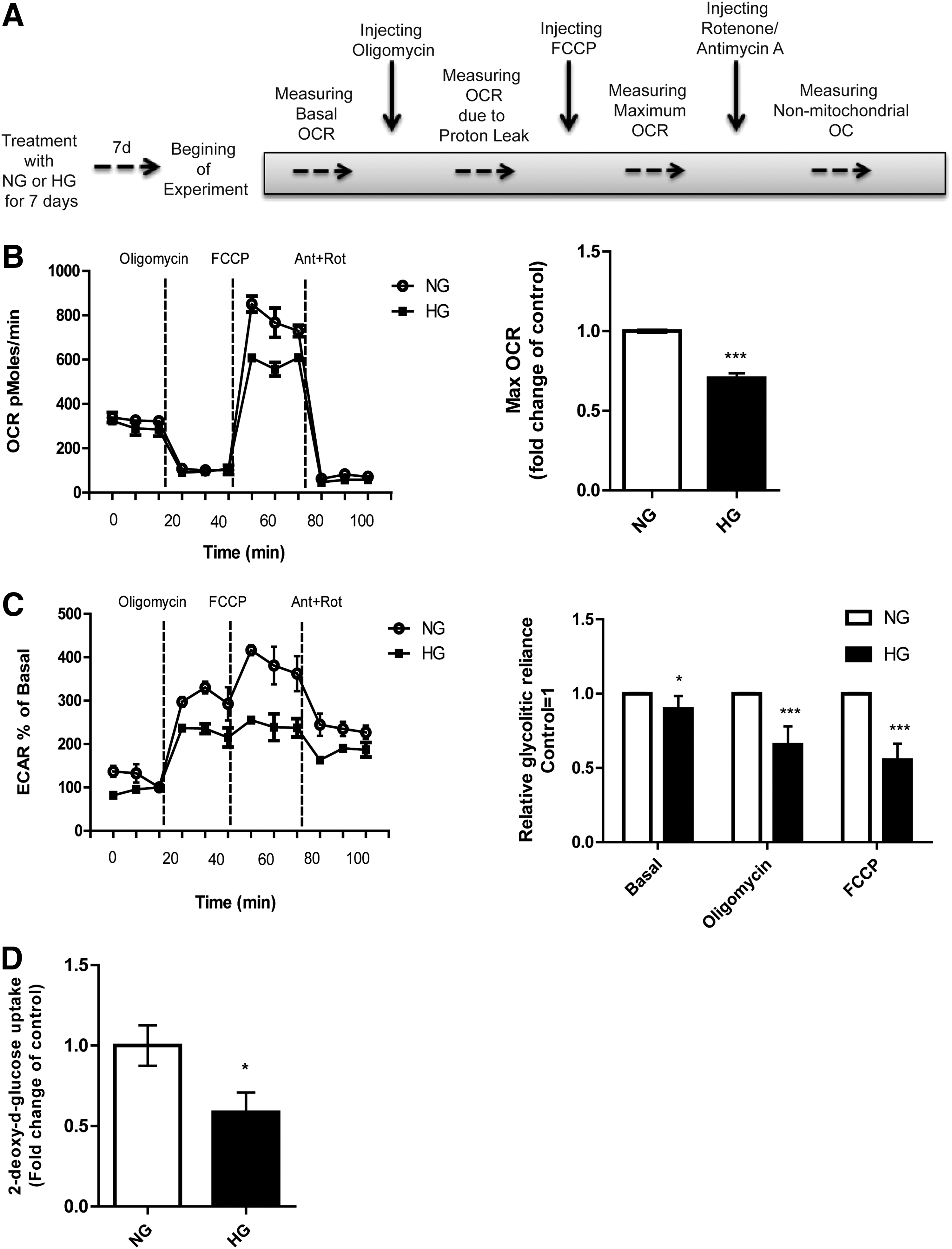
HG affects mitochondrial maximum oxygen consumption rate (OCR) and extracellular acidification in ASC.
HG affects the glucose metabolism of ASC
Measuring uptake of 2DG is the gold standard to assess glucose uptake. ASC cultured in HG showed a 41% ± 0.17% decrease in glucose uptake compared to NG medium cultured ASC (P < 0.01, Fig. 5D), which is in line with the results from the extracellular acidification measurements. In mature adipocytes insulin stimulates the uptake of glucose into these cells. ASC can be regarded as the precursors of adipocytes. Interestingly, in our experiments insulin did not affect the glucose uptake into ASC. This shows that under these conditions ASC are not sensitive to insulin in terms of increased glucose uptake. Yet, GLUT4, the insulin-sensitive glucose transporter, was normally expressed (data not shown).
The stimulation by ASC of vascular networks of HUVEC in vitro is affected by HG
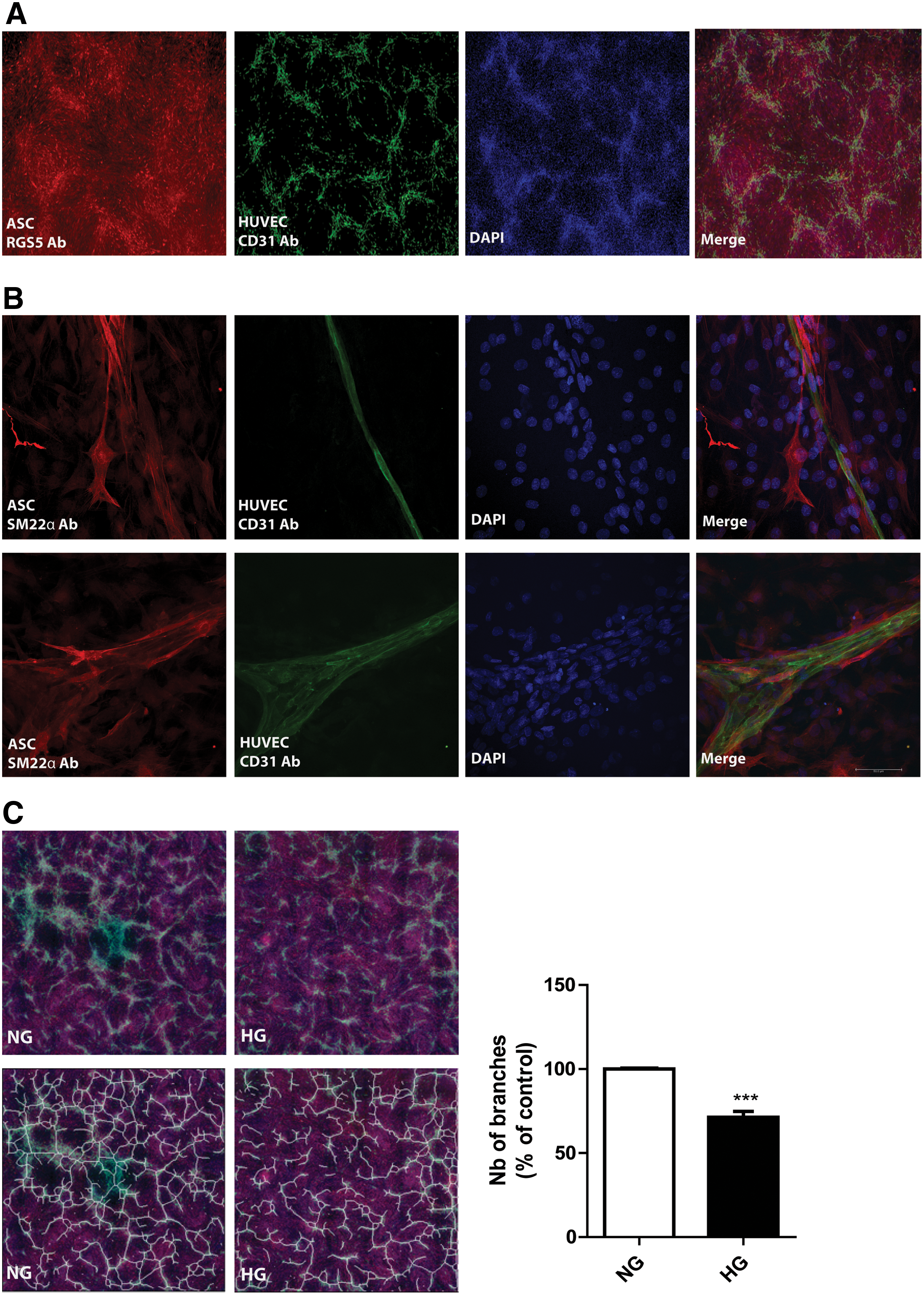
In NG medium, preformed monolayers of ASC promoted the formation of vascular network-like structures by seeded HUVEC during 7 days coculture (Fig. 6A). These networks were absent in ASC or HUVEC controls (data not shown). The vascular-like networks comprise of interconnected multicellular tubes of endothelial cells with a lumen ranging from capillary size to multicellular structures to which ASC attach in a pericytic fashion as shown by their pericytic position and expression of SM22α (Fig. 6B, Supplementary Video S1). This is corroborated by the absence of SM22α expression by the ASC that are not involved in vascular stabilization that is, which remained bound to the tissue culture plate itself (Fig. 6B). In HG medium, the ASC still supported the formation of vascular networks by HUVEC, although at a 29% ± 3.5% reduced scale (P < 0.0001, Fig. 6C).
Discussion
DR is characterized by progressive alterations in the retinal microvasculature, increasing vascular permeability caused by apoptosis in endothelial cells, and pericyte loss that leads to abnormal neovascularization in the late stage of the disease [25,26]. Consequently, pericyte replacement would be an important step in the treatment or reversal of DR. Although, ASC are pericyte-like cells and they would be promising to stabilize the vessels in DR, more studies are necessary to show the mechanism of ASC in interaction with endothelial cells in DR in vivo. In this study we showed the influence of HG on these cells in vitro individually. HG has a remarkable influence on retinal pericyte and retinal endothelial cells including bioenergetics and metabolic dysfunction, reduced mitochondrial function, and induced mitochondrial fragmentation [22,27]. The detrimental effects of HG on mitochondrial function and cellular metabolism could play a role in the apoptosis associated with the retinal pericytes and endothelial cells in DR [28]. We showed ASC had increased intracellular ROS after a long-term (7 days) exposure to HG. Chronic HG caused mitochondrial dysfunction in ASC, which was observed both as a decreased membrane potential in HG and as structural changes as well. The increased ROS production correlated with increased apoptosis and necrosis of ASC. Interestingly, the proliferation capacity of ASC after exposure to HG for 7 days was not affected. This indicates that, with respect to function, ASC are largely refractory to the mitochondrial dysfunction that is induced by HG in vitro. It is well known that HG-induced overproduction of ROS can disrupt the mitochondrial membrane potential and damage appropriate functioning of the mitochondria [29]. This deviation in mitochondrial function is an early sign of apoptosis, which was confirmed by the apoptosis increased in treated cells [30,31]. We showed that ASC, similar to most mammalian cell types in NG, form long and tubular networks of mitochondria. These networks are critical for a normal function of the mitochondria by regulation of fusion and fission events that involve the formation or breaking of the mitochondria network, respectively [32]. A decrease in the rate of fusion and a simultaneous increase in the rate of fission cause fragmentation of the mitochondrial network, which results in shorter and rounder mitochondria [33]. Our results suggest that HG-induced mitochondrial fragmentation maybe involved in the increased ROS production in ASC. It remains unclear, however, why the proliferation rate of ASC was unaffected by ROS production under HG. While mitochondria are functional, they support cell proliferation. The fact that mitochondria under HG display similar basal respiration as NG, means that HG cells can deal with the stress condition. However, they are still suffering from HG according to the statement that maximal respiration is decreased and losing their capacity to make ATP. Our results indicate that HG caused a decrease of the maximal OCR and the rate of extracellular acidification. The basal level of extracellular acidification in ASC decreased during HG exposure, and also after incubation with ATP synthase inhibitor (Oligomycin) and mitochondria uncoupler (FCCP). Corroborating the data obtained from the flux analyses, HG caused a significant decline in glucose uptake in ASC. Interestingly, this diminished rate of glycolysis was unresponsive to insulin stimulation. Our results indicate that ASC cultured in HG compensate the hyperglycemic environment by changing the metabolic capacity, as displayed by decreased oxygen consumption and glycolysis.
In this study the question was addressed how the chronic diabetic microenvironment affects the ASC's therapeutic potential. This was investigated through continuous exposure of HG on ASC and their capacity to support the formation of vascular networks by HUVEC. Interestingly, the networks were still formed, although with a reduced number of branches. The ROS-induced mitochondrial dysfunction and apoptosis, only partly affected the pericytic function of ASC, because network formation by endothelial cells was only marginally affected. Our in vitro results corroborate published data that ASC could functionally engraft in the retinal vasculature of hyperglycemic mice [17,18], while providing part of the protective measurements taken by HG-exposed ASC. Taken together, our results suggest ASC are largely resistant to long-term exposure to HG, which would explain why ASC can acquire a pericytic function in the diabetic retinal vasculature in mouse models for DR. Our current research focuses on the paracrine and juxtacrine interactions between pericytic ASC and retinal microvascular endothelial cells under HG to understand how ASC protect microvascular endothelial cells and promote their vascular stabilization.
Footnotes
Acknowledgments
TissueGnostics tissue FAXS, Austria, which is sponsored by NWO-grants 40-00506-98-9021. Additional funding: Cochilco-Fondecyt 1100995 (AAE), IMII P09-016-F (AAE). FP7 program:EULAMDIMA (PIRSES-GA-2011-295185). Dutch Diabetes Foundation (2012.00.1537). This project has received funding from the Marie Curie International Research Staff Exchange Scheme with the 7th European Community Framework Program under grant agreement no. 295185 - EULAMDIMA. The skillful technical assistance of K.A. Sjollema is greatly acknowledged.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.