
Abstract
Dipeptidyl peptidase 4 (DPP4)/CD26 truncates certain proteins, and this posttranslational modification can influence their activity. Truncated (T) colony-stimulating factors (CSFs) are decreased in potency for stimulating proliferation of hematopoietic progenitor cells (HPCs). T-CXCL12, a modified chemokine, is inactive as an HPC chemotactic, survival, and enhancing factor for replating or ex-vivo expansion of HPCs. Moreover, T-CSFs and T-CXCL12 specifically downmodulates the positively acting effects of their own full-length molecule. Other chemokines have DPP4 truncation sites. In the present study, we evaluated effects of DPP4 inhibition (by Diprotin A) or gene deletion of HPC on chemokine inhibition of multicytokine-stimulated HPC, and on chemokine-enhancing effects on single CSF-stimulated HPC proliferation, as well as effects of DPP4 treatment of a number of chemokines. Myelosuppressive effects of chemokines with, but not without, a DPP4 truncation site were greatly enhanced in inhibitory potency by pretreating target bone marrow (BM) cells with Diprotin A, or by assaying their activity on dpp4/cd26 −/− BM cells. DPP4 treatment of myelosuppressive chemokines containing a DPP4 truncation site produced a nonmyelosuppressive molecule, but one which had the capacity to block suppression by that unmodified chemokine both in vitro and in vivo. Additionally, DPP4 treatment ablated the single cytokine-stimulated HPC-enhancing activity of CCL3/MIP-1α and CCL4/MIP-1β, and blocked the enhancing activity of each unmodified molecule, in vitro and in vivo. These results highlight the functional posttranslational modulating effects of DPP4 on chemokine activities, and information offering additional biological insight into chemokine regulation of hematopoiesis.
Introduction
H
Studies of how posttranslational modification of proteins by enzymes influences the functional activity of cytokines and chemokines are under reported. Dipeptidyl peptidase 4 (DPP4) [3,4] is present as CD26 on the surface of many cells, including hematopoietic stem cells (HSCs), HPCs, and mature cells, and is found within cells, plasma, and serum [5 –7]. DPP4 cleaves dipeptides from the N-terminus usually after a penultimate proline or an alanine and with less efficiency after other penultimate amino acids such as serine. We previously demonstrated that DPP4 treatment of stromal-derived factor-1 (SDF-1; CXCL12) produced truncated CXCL12 that was inactive as a chemotactic agent, and that could block chemotaxis induced by full-length (FL) CXCL12[5]. Inhibition of DPP4 activity on target cells by small peptides such as Diprotin A (ILE-PRO-ILE) or VAL-PYR, or functional deletion of dpp4 in cells from cd26 −/− mice prevented dpp4-induced truncation of CXCL12 and allowed for enhanced chemotactic and homing activities of CXCL12 [6,7], as well as for enhanced engrafting capacity of limiting the number of mouse bone marrow (BM) cells into lethally irradiated congenic mice in competitive and noncompetitive short- and long-term HSC assays in vivo [6], and of engraftment of low numbers of human cord blood (CB) CD34+ cells injected into sublethally irradiated immunodeficient mice [8 –10].
CXCL12 also has survival-enhancing activity for HSC and HPC, enhances growth factor-induced replating of HSC in vitro, a measure of the limited self-renewal capacity of HPC, and enhances ex-vivo expansion of CD34+ CB cells [7,11 –13]. Preventing DPP4 truncation of CXCL12 significantly increased these CXCL12 activities [5,7]. Colony-stimulating factors (CSFs), including human and murine granulocyte-macrophage colony-stimulating factor (GM-CSF), granulocyte colony-stimulating factor (G-CSF), interleukin (IL)-3, and EPO, have DPP4 truncation sites, and inhibiting DPP4 truncation with Diprotin A greatly enhanced the stimulating activities of these CSFs in vitro and in vivo [7]. In addition, in vivo inhibition of DPP4 in mice by injecting Diprotin A into mice, or by oral administration of the FDA-approved small molecular weight DPP4 inhibitor, sitagliptin, greatly enhanced the engrafting capabilities of mouse BM HSC, as well as the recovery of phenotypically defined HSC and HPC, and functionally defined HPC after nonlethal doses of radiation, or after chemotherapeutic agents such as 5-flurouracil and cyclophosphamide [6,7].
Hence, inhibition of DPP4 or deletion of cd26/dpp4 increases the positively acting effects of a chemokine, CXCL12, and of selected CSFs [5,7]. It is now apparent that a large number of cytokines and chemokines have putative DPP4 truncation sites [7,14,15]. The fact that DPP4 actually truncates chemokines with DPP4 truncation sites has been reported [16 –18]. Since a number of chemokines, including CCL3/MIP-1α, CXCL4/PF-4, CXCL5/ENA-74, CXCL6/GCP-2, CXCL8/IL-8, CXCL9/MIG, and CXCL10/IP-10 with DPP4 truncation sites [14], have colony-inhibiting effects on multi-cytokine-stimulated colony formation in vitro, and myelosuppressive effects in vivo in mice [19 –42], and the chemokines CCL3/MIP1α and CCL4/MIP-1β (also with a DPP4 truncation site) have enhancing effects on single CSF-stimulated colony formation in vitro [21,36,43], we questioned what effects DPP4 truncation would have on the in vitro and in vivo functions of these chemokines. We now report that inhibition of DPP4 on target cells in vitro and deletion of cd26/dpp4 in mice enhances the activities of these chemokines, and that DPP4 posttranslational modification of selected chemokines by truncation modifies their functional activities.
Materials and Methods
Mice and cells
BM cells were from male and female C57Bl/6 and cd26−/−/dpp4 mice (on a C57Bl/6 mouse strain background) at 6–10 weeks old, as previously described [6,7]. The Indiana University Committee on Use and Care of Animals approved all the animal studies (IACUC Study No: 10985 MD/R/E/AR). Cells were unseparated and either left untreated or were preincubated with Diprotin A to inhibit DPP4 enzymatic activity, and washed before the addition of CSFs−/+ SCF,−/+ chemokines [3 –5]. CCR5−/− and CCR5+/+ mice were purchased from Jackson Laboratories (Bar Harbor, ME).
Chemokines and cytokines
All recombinant human chemokines and mouse cytokines were purchased from R&D Systems (Minneapolis, MN), and had previously been tested for activity [21,24,25,29 –43].
DPP4 pretreatment of chemokines
This was done as previously described [7]. Before their use, Diprotin A was added to chemokine preparations to inactivate the DPP4 before the chemokine was assayed for activity, as noted in text and figure legends.
In vitro assays
Multi-cytokine-stimulated and single CSF-stimulated BM colony formation, which respectively represent immature and more mature HPC, was performed as previously reported [7,21].
In vivo assays
These were done as previously reported [22,30,33,40], and are described within the text and figure legends.
Statistical analysis
Significant differences were determined by two-tailed Student's t-test.
Results
Inhibition of dpp4 enzymatic activity or deletion of dpp4 genes in mouse BM cells enhances selected chemokine-mediated myelosuppression of HPC proliferation in vitro
Chemokine structures and receptors have been well described [44 –48]. Some, but not all, chemokines suppress HPC proliferation in vitro and in vivo [49,50]. Of the myelosuppressive chemokines [49,50], a number have DPP4 truncation sites [14,16 –18]. Since chemokine inhibition is manifest on target BM cell populations that express CD26, a cell surface DPP4, we reasoned that this DPP4 could possibly modify the actions of myelosuppressive chemokines. We chose representative myelosuppressive chemokines with DPP4 truncation sites (Table 1) and assessed their suppressive activity on multi-cytokine [recombinant murine (rm)-GM-CSF and rm-stem cell factor]-stimulated granulocyte macrophage progenitor cell (CFU-GM) colony formation of C57Bl/6 mouse BM cells that were either preincubated with control (phosphate-buffered saline) medium or with Diprotin A (5 mM; ILE-PRO-ILE; a specific DPP4 inhibitor) for 1 h before washing, or with untreated BM cells from cd26 −/− mice (on a Bl/6 mouse strain background) (Fig. 1A; results are expressed as percent inhibition of CFU-GM colony formation). Multi-cytokine-stimulated HPC are more immature subsets of HPC than those stimulated by a single CSF [1]. The five chemokines (CCL3, CXCL6, CXCL9, CXCL10, and CXCL4) tested suppressed colony formation of C57Bl/6 BM cells pretreated with control medium, consistent with results in previous studies using untreated BM. Maximal suppression (∼50% inhibition), previously shown to relate to percent of progenitor cells in S-phase of the cell cycle at the time the chemokines are added, was detected at 100 ng/mL chemokine, with greatly reduced, but still significant suppression at 10 ng/mL chemokine. In contrast, suppressive activity was detected at 10- to 100-fold lower concentrations (0.01–1.0 ng/mL) when target BM population were first pulsed with Diprotin A. Enhanced suppressive activity at low concentrations of chemokine was confirmed against BM cells from cd26 −/−- mice.
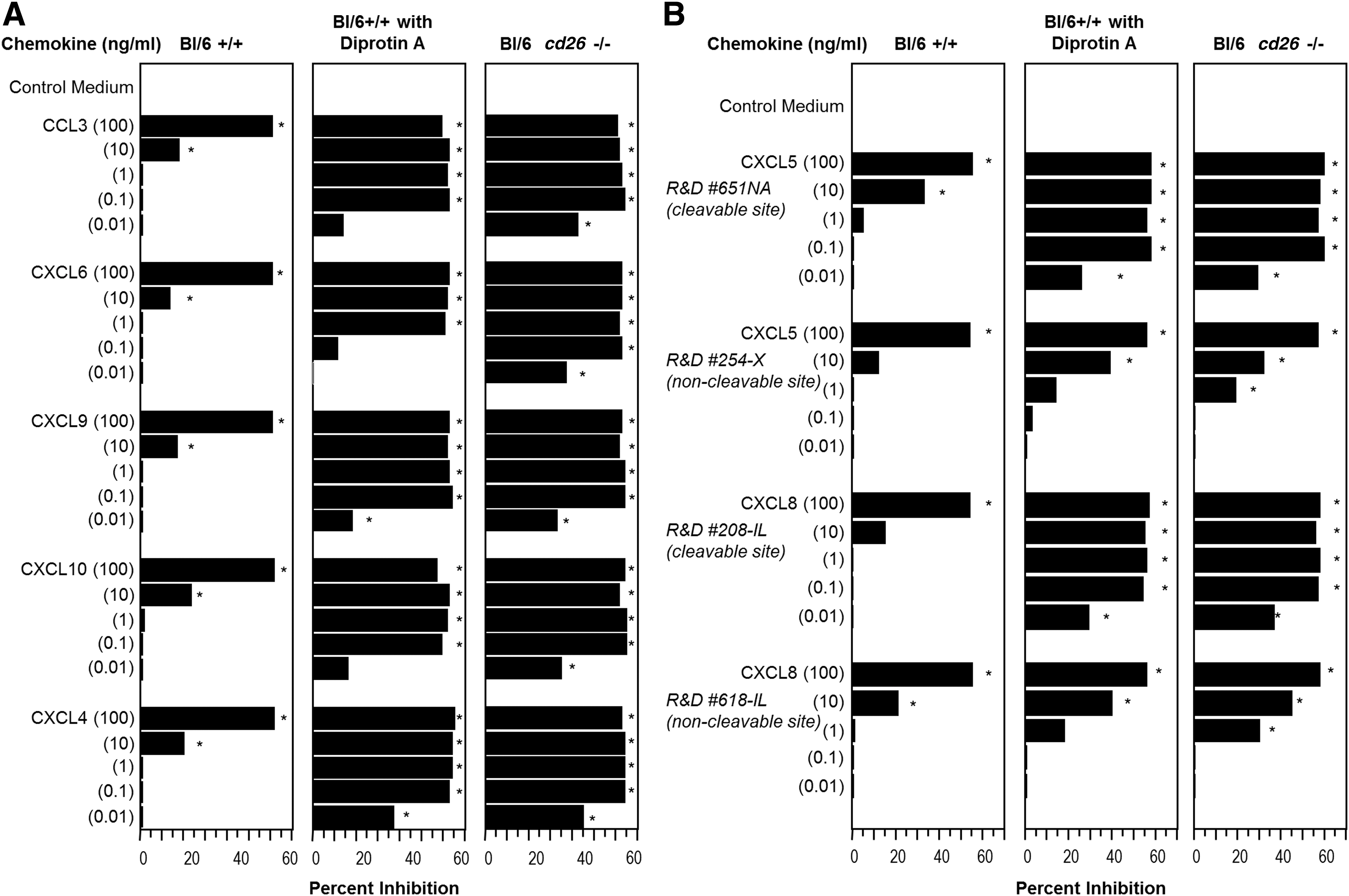
Influence of DPP4 inhibition or cd26/dpp4 deletion on chemokine suppression of mouse BM CFU-GM colony formation in vitro. BM cells from control mice (Bl/6) preincubated with control medium for 1 h followed by washing of cells, control mouse cells preincubated with 5 mM Diprotin A for 1 h and washed, or cd26/dpp4
−/− cells were plated at 5 × 104 cells/mL in the presence of rmGM-CSF (10 ng/mL) and rmSCF (50 ng/mL) minus and plus various concentrations of chemokines as indicated. Results are expressed as percent inhibition compared to control medium. For
Bold letters denote the DPP4-truncation site.
Three chemokines that have a DPP4 truncation site, but are not inhibitory (CXCL12, CCL4, and CCL5) had no myelosuppressive activity when assayed at 100, 10, 1.0, 0.1, or 0.01 ng/mL on either control BM, BM cells preincubated with Diprotin A, or cd26 −/− BM (0–10% inhibition, P > 0.05; data not shown). We compared myelosuppressive effects of CXCL5 (a known suppressive chemokine with a DPP4 truncation site) with that of CXCL5 produced without a DPP4 cleavage site (Table 1), and also of CXCL8 (another known suppressive chemokine with a DPP4 truncation site) to that of CXCL8 produced without a DPP4 cleavage site (Table 1). As seen in Fig. 1B, CXCL5 and CXCL8 were equally inhibitory within the range previously reported for these chemokines (significant suppression to 10 ng/mL) on control BM cells. In contrast to FL forms of CXCL5 and CXCL8, which manifested significant suppressive activity on Bl/6 BM cells pretreated with Diprotin A or on cd26 −/− BM cells at concentrations as low as 0.01 ng/mL, CXCL5 and CXCL8 without DPP4 truncation sites, were only slightly or not at all enhanced in suppressive activity on DPP4-inhibited or cd26 −/− BM cells compared to control BM cells. XCL1, a myelosuppressive chemokine without a DPP4 truncation site, was not enhanced in activity at lower chemokine concentrations when target BM cells were cd26 −/− (Fig. 2). Thus, myelosuppressive chemokines with, but not without, a DPP4 truncation site are significantly enhanced in myelosuppressive activity at lower chemokine concentrations when DPP4 is either inhibited or deficient in target BM cells.
Influence of DPP4 treatment of chemokines on their suppressive activity on control (Bl/6) and cd26 −/− mouse BM CFU-GM colony formation in vitro and their effects on the suppressive activity of their own FL chemokine molecule. BM cells were stimulated as in the legend to Fig. 1, and results expressed as mean colonies ±1SD per 5 × 104 cells plated. *P < 0.05 compared to control. T, DPP4-treated molecule after preincubation with soluble DPP4; NT, molecule without DPP4 truncation site that was pretreated with soluble DPP4. FL, full length.
DPP4-treated chemokines are inactive in vitro as myelosuppressive proteins, and block the inhibitory activity of the FL form of that specific chemokine
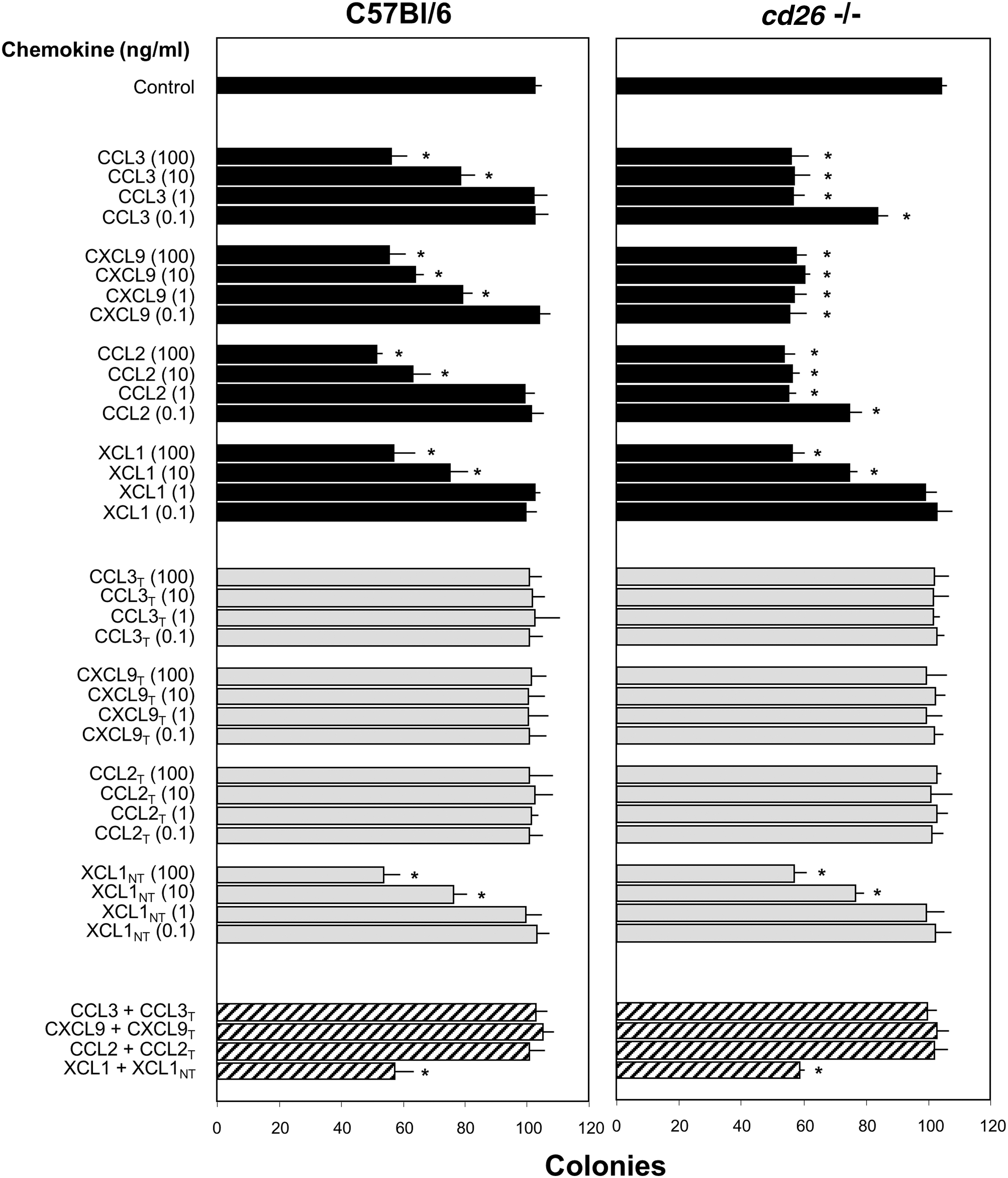
To test effects of DPP4 treatment on myelosuppressive activities of chemokines, we studied CCL3, CXCL9, and CCL2/MCP-1, each with a DPP4 truncation site, and XCL1 without a DPP4 truncation site (Table 1) for inhibition of colony formation by Bl/6 or cd26 −/− BM cells (Fig. 2; data expressed as colonies per 5 × 104 cells plated). Lower concentrations (0.1–1.0 ng/mL) of CCL3, CXCL9, and CCL2, but not of XCL1 (no DPP4 truncation site), were myelosuppressive on cd26 −/− compared to control Bl/6 BM (see black bars). DPP4 treatment of chemokines before assay, followed by addition of Diprotin A to inactivate the DPP4 before use in the assay resulted in loss of myelosuppressive activity for CCL3, CXCL9, and CCL2, but not for XCL1, on control and cd26 −/− BM cells (see gray bars). DPP4-treated CCL3, CXCL9, and CCL2 at a 1:1 ratio of 100 ng DPP4-treated:100 ng untreated chemokine blocked myelosuppression of their respective FL chemokines. DPP4-treated XCL1, which lacks a DPP4 truncation site, did not block activity of the FL form of XCL1 at a 1:1 ratio of 100 ng:100 ng (see crosshatched bars toward bottom of Fig. 2). Thus, DPP4-treated chemokines with DPP4 truncation sites do not manifest myelosuppressive activity, but they block inhibition of their FL form.
We next used cd26 −/− mouse BM cells to determine what ratio of DPP4-treated to FL chemokine was required for DPP4-treated chemokine to block myelosuppression by FL chemokine, and if DPP4-treated chemokine of one type could block the FL form of another type of chemokine (Fig. 3A, B; with results expressed as colony numbers per 5 × 104 cells/mL). As shown in Fig. 3A, DPP4-treated CCL3, CXCL9, and CCL2, but not DPP4-treated XCL1, were inactive as suppressor molecules (gray bars). DPP4-treated CCL3, CXCL9, and CCL2 were able at a ratio of 1:1 (100 ng:100 ng), but not at a 100 ng:10 ng or 1 ng ratio of DPP4-treated to FL molecule to block suppression of FL chemokine of that type (crosshatched bars). DPP4-treated CXCL9 and CCL2, or XCL1 were not able at 1:1 (100 ng:100 ng) ratios to block the suppressive activity of FL CCL3; DPP4-treated CCL3 and CCL2, and XCL1 were not able to block suppression by FL CXCL9; DPP4-treated CCL3, CXCL9, and XCL1 were not able to block the suppressive activity of FL CCL2, and neither DPP4-treated XCL1 nor the DPP4-treated CCL3, CXCL9, or CCL2 were able to block the activity of XCL1 (see crosshatched bars, Fig. 3). Thus, DPP4-treated forms of one chemokine were not able to block myelosuppressive activities of another chemokine molecule. Lack of cross-blocking activity is not unexpected as these chemokines bind different chemokine receptors. As noted in our previous studies, blocking effects of truncated GM-CSF on the activity of FL GM-CSF were mediated at the receptor level [7].
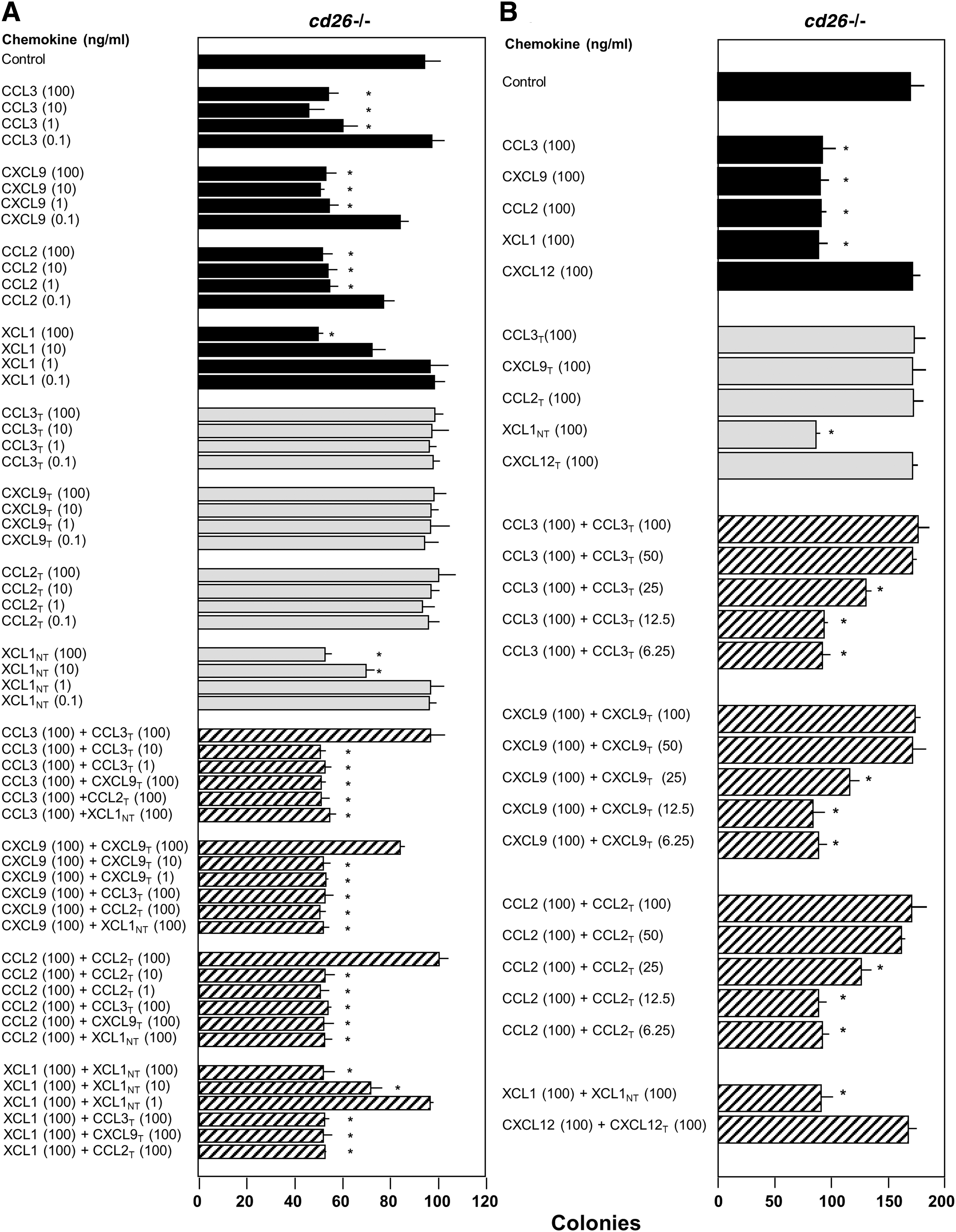
Influence of DPP4 treatment of chemokines on the suppressive activity of cd26
−/− mouse BM CFU-GM colony formation in vitro. (
Myelosuppressive effects of certain chemokines are mediated through specific receptors (eg, for CXCL8/IL-8 [30] and CCL2/MCP-1 [32,35]). However, the receptor-mediating myelosuppression by CCL3/MIP-1α is not known. CCL3/MIP-1α has two main receptors: CCR1 and CCR5 [44
–48]. Inhibitory effects of CCL3/MIP-1α are not mediated through ccr1 [36]. Therefore, we used ccr5
−/− mice to see if CCL3/MIP-1α was active as a myelosuppressive agent on these cells in vitro and in vivo. CCL3/MIP-1α was as inhibitory on ccr5
−/− BM cells in vitro (Supplementary Fig. S1; Supplementary Data are available online at
To determine the concentration of DPP4-treated chemokine necessary to block the inhibitory activity of their own FL chemokine, we used amounts of DPP4-treated molecules that were 1/2, 1/4, 1/8, or 1/16 that of the FL molecule. As shown in Fig. 3B (crosshatched bars), 50 ng/mL of DPP4-treated CCL3, CXCL9, and CCL2, respectively, completely blocked the suppressive activity of 100 ng/mL of their own FL chemokine; 25 ng/mL of each DPP4-treated chemokine partially blocked suppression of 100 ng/mL of their own chemokine. Thus, DPP4-treated chemokine could at one-fourth to one-half the concentration of its own FL chemokine partially or fully block myelosuppression. This is consistent with our previous report that lower amounts of truncated GM-CSF outcompeted higher amounts of FL GM-CSF for receptor binding [7]. As an additional control, CXCL12, a nonmyelosuppressive chemokine, which has a DPP4 truncation site, was not myelosuppressive in either FL or DPP4-treated form, and the combination of FL and DPP4-treated CXCL12 was not myelosuppressive.
DPP4-treated chemokines are inactive in vivo as myelosuppressive proteins, and block the in vivo inhibition of multicytokine-responsive HPC numbers and cycling by the FL form of that specific chemokine
To determine if effects noted in vitro for FL and DPP4-treated chemokines (Figs. 1 –3) are also manifested in vivo. FL or DPP4-treated CCL3 (Fig. 4A), CXCL8 (Fig. 4B), and CXCL9 (Fig. 4C) were injected subcutaneously (s.c.) into Bl/6 mice each alone, or in combination (FL and DPP4-treated molecules were injected s.c. separately at different sites in the mice). Mice were sacrificed and analyzed 24 h later, a time shown by us previously to detect significant inhibition of numbers and cycling status of HPC [22,33]. FL chemokines suppressed numbers and cycling of HPC, DPP4-treated chemokines were not suppressive, and each DPP4-treated chemokine blocked the activity of their own FL chemokine (Fig. 4). Thus, in vivo effects of FL and DPP4-treated chemokines mimic their in vitro actions.
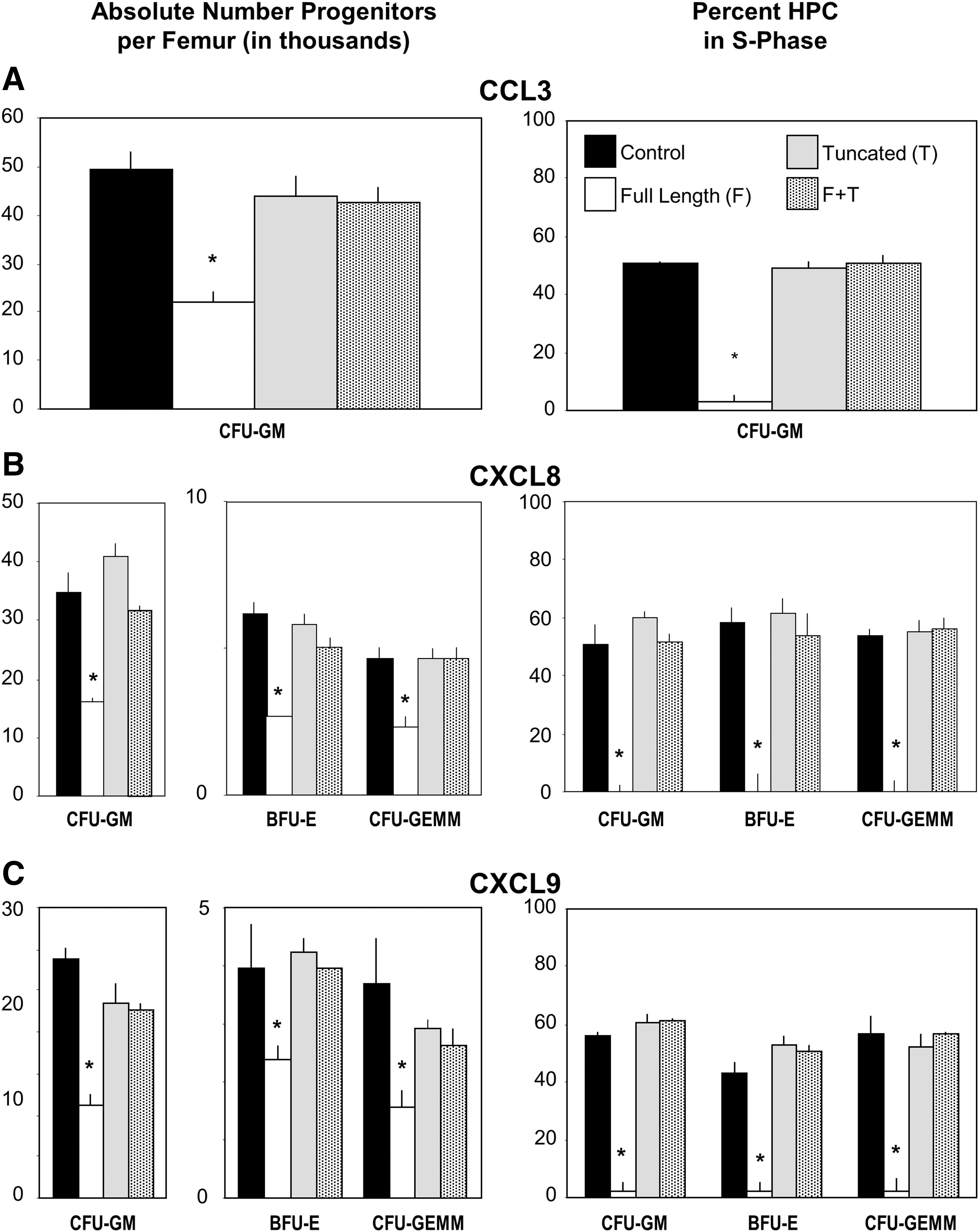
Influence of s.c. injection of FL, DPP4-treated, and the combination of DPP4-treated plus FL chemokine (each given at a different site) to C57Bl/6 mice on absolute numbers and cycling status of BM hematopoietic progenitor cells. Results are shown as mean numbers ±1SEM for three mice per group with each mouse assessed individually. Experiments for groups
DPP4-treated CCL3 and CCL4 are inactive as enhancing proteins for proliferation of single cytokine-responsive HPC in vitro, and block the enhancing activity of their FL proteins
CCL3 and CCL4/MIP-1β enhance colony formation of CFU-GM and CFU-macrophage (M) stimulated, respectively by GM-CSF alone and M-CSF alone in vitro [21,36,43]. CCL3, but not CCL4, acts as a suppressive agent for proliferation of multicytokine-stimulated colony formation of HPC, and since DPP4-treatment of CCL3 changes it to noninhibitory molecule against multicytokine-stimulated proliferation of HPC, but does block this suppression by FL CCL3, we tested whether DPP4 treatment of CCL3 and CCL4 would influence their enhancing activity for proliferation of single cytokine-stimulated HPC. Single cytokine-responsive HPC are a more mature progenitor cell population than multicytokine-responsive HPC [1]. As shown in Fig. 5A and B for GM-CSF-stimulated CFU-GM and Fig. 5C and D for M-CSF-stimulated CFU-M on BM cells not pretreated with Diprotin A (black bars in Fig. 5A–D), DPP4-treated CCL3 and CCL4 were inactive as enhancing agents for GM-CSF-responsive CFU-GM and for M-CSF-responsive CFU-M. Moreover, DPP4-treated CCL3 and CCL4, respectively, blocked enhancement of their FL counterparts. Since CCL3 and CCL4 mediate their enhancing activities through a common chemokine receptor, CCR1, we evaluated the capacity of DPP4-treated CCL3 and CCL4 to reciprocally block the enhancing activity of the other FL chemokine. DPP4-treated CCL3 blocked the enhancing effects of FL CCL4, and DPP4-treated CCL4 blocked the enhancing effects of FL CCL3 (Fig. 5A, C).

Effects of DPP4-treated CCL3 and CCL4 on in vitro colony formation of GM-CSF-stimulated CFU-GM
We comparatively evaluated the enhancing effects of CCL3 and CCL4 on colony formation with or without first pretreating them with Diprotin A to inhibit DPP4 (Fig. 5A, B). As reported previously, GM-CSF had greater CSF activity on Diprotin A-pretreated than control BM cells [7]. However, neither FL CCL3 nor CCL4 demonstrated enhancing effects on GM-CSF stimulation of Diprotin A-pretreated cells, with colony formation of GM-CSF-stimulated Diprotin A-pretreated cells being comparable to that of GM-CSF-stimulated colony formation of control (C57Bl/6) BM cells plated in the presence of FL CCL3 or CCL4. Of interest, DPP4-treated CCL3 and CCL4, while having no enhancing activity on GM-CSF-stimulated control BM cells, had an inhibitory activity on GM-CSF-stimulated Diprotin A-pretreated BM cells. This highlights the complexity of cytokine regulation of hematopoiesis when the action of DPP4 and its effects on chemokine action are taken into consideration.
M-CSF does not have a DPP4 truncation site, and inhibition of CD26 on target cells does not enhance M-CSF-stimulated colony formation [7]. Thus, we did not pretreat target cells with Diprotin A for effects of CCL3 and CCL4 on M-CSF-stimulated colony formation (Fig. 5C, D), FL-CCL3, and CCL4-enhanced M-CSF-stimulated colonies, but DPP4-treated CCL3 and CCL4 were without enhancing activity, and blocked the effects of the FL CCL3 and CCL4.
Administration of CCL3 to mice in vivo results in enhanced proliferation of GM-CSF- and M-CSF-responsive HPC, whereas DPP4-treated CCL3 is not active in this action and counteracts the effect in vivo of administrated FL CCL3
We had not previously assessed the in vivo activity of a chemokine on numbers and proliferation of single cytokine-responsive HPC, which in context of CCL3 or CCL4-enhancing activity would allow us to compare their in vitro and in vivo effects. We now report effects of in vivo administration s.c. of CCL3, DPP4-treated CCL3, and the combination of CCL3 and DPP4-treated CCL3 (each injected s.c. into different sites) on absolute numbers and cycling status (% HPC in S-Phase of the cell cycle) of CFU-GM responsive to stimulation by GM-CSF, and of CFU-M responsive to stimulation by M-CSF. As noted in Fig. 6, CCL3 administration enhanced absolute numbers and cycling status of GM-CSF-responsive CFU-GM and M-CSF-responsive CFU-M. DPP4-treated CCL3 had no enhancing effects on absolute numbers or cycling status of GM-CSF-responsive CFU-GM and blocked the enhancing effects of non-DPP4-treated FL CCL3 on these actions. DPP4-treated CCL3 had no enhancing effect on absolute numbers of M-CSF-responsive CFU-M, and blocked the enhancing effect of non-DPP4-treated CCL3 on this effect. In contrast to effects of DPP4-treated CCL3 on cycling of GM-CSF-responsive CFU-GM, DPP4-treated CCL3 decreased cycling of M-CSF-responsive CFU-M in the absence and presence of administration of FL CCL3. Hence, in vitro enhancing effects of CCL3 and of DPP4-treated CCL3 on single cytokine-stimulated HPC are confirmed in vivo.
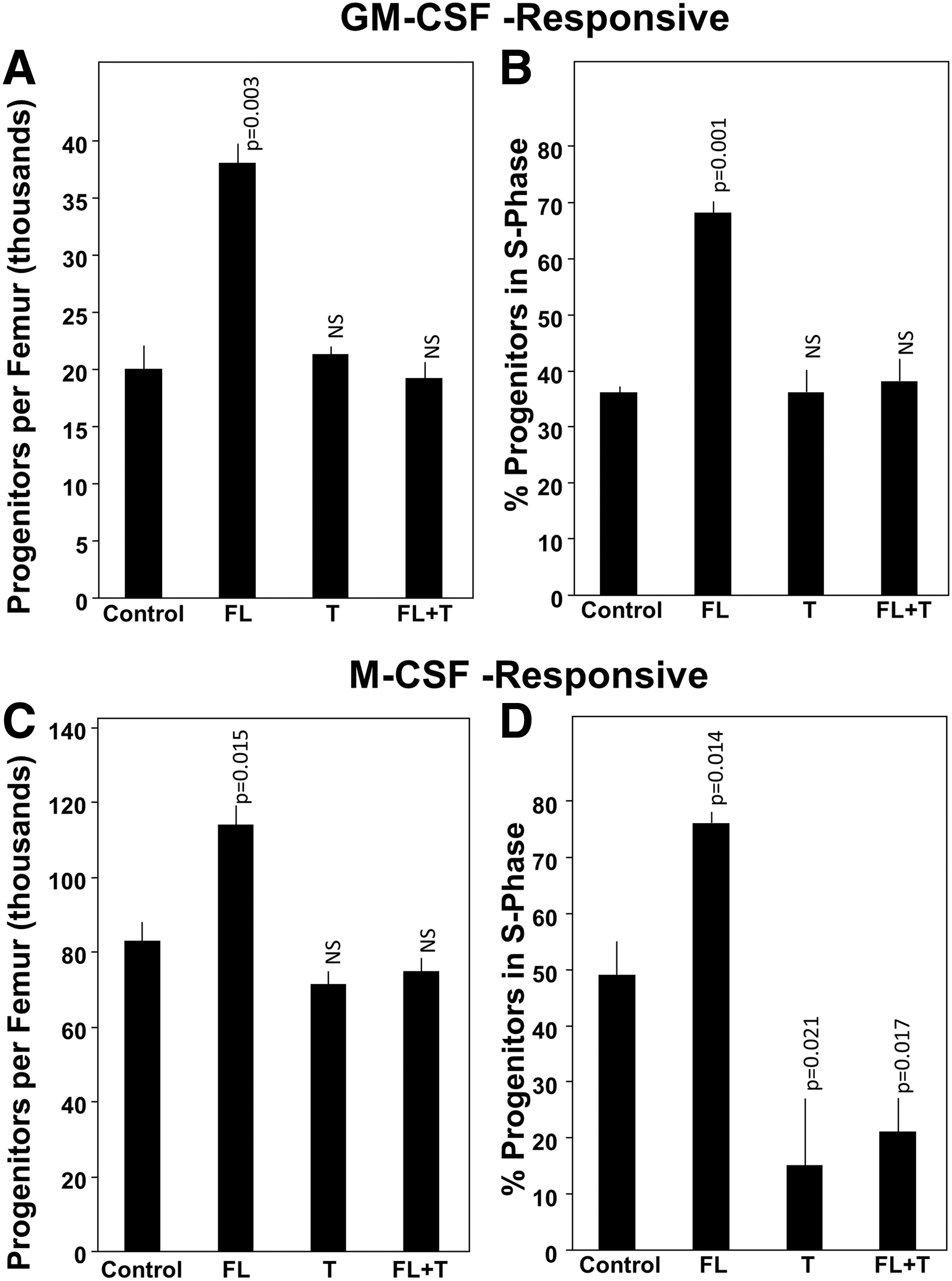
Effects of CCL3 in vivo in C57Bl/6 mice (not pretreated) on absolute numbers and cycling status of GM-CSF-responsive BM CFU-GM
Discussion
Chemokines are important members of the cytokine family of biological regulators [1,2,44 –48]. Posttranslational modification of cytokines may change their biological activities. DPP4 truncation of the CSFs: GM-CSF, G-CSF, IL-3, and EPO, and of SDF-1/CXCL12 results in a less efficient CSF for stimulation of colony formation, and an inactive SDF-1/CXCL12 for chemotaxis and survival of HPCs [5 –7]. Moreover, these truncated molecules downmodulate the activities of the FL forms of their molecules. Since regulation of hematopoiesis reflects on both positively and negatively acting cytokines and chemokines [1], and a number of chemokines that have been implicated in regulation of hematopoiesis in vitro and in vivo [48 –50] have DPP4 truncation sites [14], we assessed the biological effects in vitro and in vivo of DPP4-treated myelosuppressive chemokines (CCL2, CCL3, CXCL4, CXCL5, CXCL6, CXCL8, CXCL9, and CXCL10) on immature subsets of HPCs that in vitro respond to stimulation by multiple cytokines, and of DPP4-treated enhancing chemokines (CCL3 and CCL4) that act on single CSF (GM-CSF or M-CSF)-stimulated more mature subsets of HPCs.
It is clear that DPP4 treatment of these members of the CC and CXC family of chemokines modify their biological activities. Myelosuppressive chemokines lose their suppressive activities when treated with DPP4, and these DPP4-treated chemokines compete with their FL forms to blunt the inhibitor activity manifested by the FL forms. Enhancing activity of the two members of the CC family, CCL3 and CCL4, that manifest this activity is lost upon DPP4 treatment of these chemokines and DPP4-treated CCL3 and CCL4 reciprocally block enhancement by their FL forms, likely through their receptor-binding capacity to the same receptor, CCR1, which is involved in the enhancing activities of these chemokines [32,36]. The in vitro activities of both the myelosuppressive and enhancing chemokines studied in this report, was confirmed in vivo in mice, suggesting that DPP4 truncation of these chemokines may be of physiological relevance. Of interest, is that CCL3, but not CCL4, can both suppress (on multi-CSF-stimulated immature subsets of HPC) and enhance (on single CSF stimulation of more mature subsets of CFU-GM) colony formation in vitro and progenitor cell proliferation in vivo.
While the enhancement of CCL3 is manifested through CCR1, as is that of CCL4, it is not yet clear what receptor mediates the myelosuppressive activity of CCL3. CCR1 does not mediate suppression through CCL3 [36] and it is now clear that CCR5, another major receptor for CCL3, does not mediate suppression by CCL3 in vitro or in vivo. Thus, a search is still on for the receptor mediating the myelosuppressive effects of CCL3.
We still have much to learn about cytokine- and chemokine-mediated regulation of hematopoiesis, and whether posttranslational modifications of these proteins is likely playing a role, perhaps even more than a blocking activity on the FL forms of these proteins. Future studies may define whether or not DPP4-truncated molecules have qualitative, in addition to quantitative, signaling and biological effects different from their FL forms. We find it intriguing that DPP4-treated CCL3 and CCL4 not only did not have the enhancing activity on GM-CSF-stimulated colony formation when the target cells were pretreated with Diprotin A to inhibit cellular DPP4 activity, but also in this situation, the DPP4-treated chemokines manifested suppressive activity on more mature subsets of CFU-GM. We have not previously found any of the FL chemokines to be suppressive on single cytokine-stimulated colony formation by CFU-GM [49,50].
It may be that DPP4 truncation of cytokines and chemokines is most relevant in a stressed situation, when the hematopoietic system is recovering from radiation or toxic drugs, or perhaps during disease. DPP4 does not seem to be as obviously involved with hematopoiesis during nonstressed steady-state conditions, where hematopoiesis seems relatively similar in control mice versus cd26/dpp4 −/− mice or in mice receiving a DPP4 inhibitor such as Diprotin A or sitagliptin. Sitagliptin is currently in clinical trial to accelerate recovery of single CB transplantation for treatment of patients with leukemia and lymphomas [51,52]. DPP4 seems to be mainly active during recovery from radiation or chemotherapy drugs, such as 5-fluorouracil. This is where inhibition of DPP4 activity shows effects, such as accelerated hematopoietic recovery [7], although this does not rule out effects of DPP4 activity in steady-state conditions.
Further studies on the role of DPP4 in cytokine/chemokine-mediated hematopoietic, and perhaps other tissue regulatory events, may help to better define normal physiological cell and tissue activities, and abnormalities during stress and disease. It may even be possible to use truncated molecules or prevention of DPP4 truncation to treat various disease states.
Footnotes
Acknowledgments
These studies were supported by the US Health Service National Institutes of Health (NIH) grants R01 HL056416, R01 HL067284, R01 HL112669, and a Center of Excellence in Molecular Hematology grant P01 DK090948 (and U54 DK106846) to H.E.B. M.C. was supported as a post-doc on NIH T32 DK07549 to H.E.B. T.B.C., currently an assistant professor at UCSF, was supported as a pre-doc on NIH T32 DK07519 during some of the beginning studies reported in this article.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.