
Abstract
Preterm white matter injury (WMI) is an important cause for long-term disability. Stem cell transplantation has been proposed as a novel therapeutic approach. However, intracerebral transplantation is not feasible for clinical purpose in newborns. Intranasal delivery of cells to the brain might be a promising, noninvasive therapeutic approach to restore the damaged brain. Therefore, our goal is to study the remyelinating potential of human Wharton's jelly mesenchymal stem cells (hWJ-MSCs) after intranasal delivery. Wistar rat pups, previously brain-damaged by a combined hypoxic-ischemic and inflammatory insult, received hWJ-MSC (150,000 cells in 3 μL) that were intranasally delivered twice to each nostril (600,000 cells total). WMI was assessed by immunohistochemistry and western blot for myelination, astrogliosis, and microgliosis. The expression of preoligodendrocyte markers, and neurotrophic factors, was analyzed by real-time polymerase chain reaction. Animals treated with intranasally delivered hWJ-MSC showed increased myelination and decreased gliosis compared to untreated animals. hWJ-MSC may, therefore, modulate the activation of microglia and astrocytes, resulting in a change of the brain microenvironment, which facilitates the maturation of oligodendrocyte lineage cells. This is the first study to show that intranasal delivery of hWJ-MSC in rats prevented hypomyelination and microgliosis in a model of WMI in the premature rat brain. Further studies should address the dose and frequency of administration.
Introduction
A
The major underlying causes for WMI are hypoxia-ischemia and inflammatory changes including proinflammatory cytokine release in brain parenchyma as a consequence of intrauterine infection in preterm birth [1,2]. A combination of hypoxia-ischemia and inflammation strongly increases the risk of developing WMI [5,6]. Studies identified preoligodendrocytes (preOLs) as the most vulnerable cells to hypoxia-ischemia and inflammatory damage. The peak period of WMI occurrence (24–32 weeks of gestation) coincides with the highest presence of preOLs in the white matter of the brain. Thus, it has been hypothesized that the hypomyelination in WMI is due to a depletion of preOLs or an inability of those to later differentiate into mature myelinating oligodendrocytes [7].
Human Wharton's jelly mesenchymal stem cell (hWJ-MSC) have been proposed as a therapeutic agent in a wide range of diseases. Importantly, it has been shown that they are able to elicit neuronal repair, have immunomodulatory properties, are multipotent, and secrete several trophic factors important for neuroregeneration [8,9].
While previous preclinical studies showed the neuroregenerative potential of stem cell transplantation for perinatal brain damage, cellular transplantation in these models usually involved intracerebral or intrathecal administration [10]. If this treatment shall be developed for wider clinical use in human newborns, a less invasive method is needed.
Several animal models of WMI have been established [11,12]. We have developed a combined model of hypoxic-ischemic and inflammatory (HII) preterm brain damage in postnatal day 2 rats, when the rat's brain development corresponds to 23–32 weeks of human gestational age [13].
Our aim was to determine the neuroregenerative potential of hWJ-MSC using the noninvasive intranasal administration route.
Materials and Methods
Human Wharton's jelly mesenchymal stem cell
After informed consent, umbilical cords from healthy term deliveries were collected. The study was approved by the institutional review boards of the University Hospital Bern and the Canton of Bern.
The hWJ-MSC were isolated from the Wharton's jelly as described previously [14]. The cells were expanded in Dulbecco's modified Eagle's medium (DMEM)/F12 supplemented with 10% fetal calf serum (FCS), 2 mM glutamax, 100 U/mL penicillin, and 100 mg/mL streptomycin (Thermo Fisher Scientific, Waltham, MA). hWJ-MSC were positive for CD105, CD73, and CD90 and negative for myeloid and hematopoietic cell lineage-specific antigens and were able to differentiate into osteocytes, chondrocytes, and adipocytes [14].
Induction of WMI and intranasal delivery of hWJ-MSC
All animal procedures were approved by the Veterinary Department of the Canton of Bern, Switzerland. Wistar rat pups out of two litters were randomly assigned to three different experimental groups: control (n = 5), HII (n = 6), and HII+hWJ-MSC (n = 8). Lipopolysaccharide (LPS, 0.1 mg/kg BW; Sigma-Aldrich, St. Louis, MO) was intraperitoneally administrated in 2 days old Wistar rat pups. On the following day, animals were anesthetized with 4% isoflurane and the left common carotid artery was cauterized followed by hypoxia (3 L/min, 8% O2/92% N2) for 65 min. Control rat pups received 0.9% NaCl and were sham operated.
Two days after damage, 3 μL drops containing hWJ-MSC (50,000 cells/μL, passage 6) or DMEM/F12 + 10% FCS (HII group) were delivered intranasally twice to each nostril making a total of 12 μL (600,000 cells). Thirty minutes before cell delivery, 3 μL of hyaluronidase (100 U in phosphate-buffered saline; Sigma-Aldrich) were administrated once to each nostril to increase nasal mucosa permeability. Brains were harvested 7 days after treatment.
RNA and protein isolation
Proteins from frozen brain tissue were isolated using the QIAschredder and the AllPrep DNA/RNA/Protein Mini Kit. RNA from paraformaldehyde-fixed paraffin-embedded brain slices was isolated with the RNeasy FFPE Kit according to manufacturer's instructions (Qiagen, Hilden, Germany). The concentration of RNA was measured by Nanodrop spectrometry (Thermo Fisher Scientific). RNA quality was determined with the OD 260/280 ratio, which was between 1.8 and 2.1. Up to 5 μg of RNA were reverse transcribed using the SuperScript III First-Strand Synthesis System (Thermo Fisher Scientific). The total protein concentration of the brain tissue was determined with the bicinchoninic acid protein assay kit (Sigma-Aldrich).
Real-time polymerase chain reaction
Gene expression of chondroitin sulfate proteoglycan 4 [Cspg4, TaqMan gene expression (GE) assay ID: Rn00578849_m1], platelet-derived growth factor receptor alpha (Pdgfra, TaqMan GE assay ID: Rn01417534_g1), S100 calcium-binding protein B (S100b, GE assay ID: Rn04219408_m1), brain-derived neurotrophic factor (Bdnf, GE assay ID: Rn02531967_s1), insulin-like growth factor 1 (Igf1, GE assay ID: Rn00710306_m1), and vascular endothelial growth factor A (Vegfa, GE assay ID: Rn01511601_m1; all Thermo Fisher Scientific) were assessed in coronal brain sections at the level of the middorsal hippocampus.
The polymerase chain reaction (PCR) was run with the following program on a 7300 Real-Time PCR System (Thermo Fisher Scientific): 2 min at 50°C, 10 min at 95°C, followed by 45 cycles of 15 s at 95°C and 1 min at 60°C. The transcripts were normalized to the reference gene glyceraldehyde-3-phosphate dehydrogenase. Data were analyzed using the 7300 System Software (Thermo Fisher Scientific) and expressed as fold change relative to rNPC cultured alone for the in vitro and relative to total rat brain RNA (Amsbio, Abingdon, United Kingdom) for the in vivo experiments.
Western blotting
Proteins were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) on a 15% gel for myelin basic protein (MBP) detection and on a 4%–20% gradient gel (Bio-Rad, Hercules, CA) for glial fibrillary acidic protein (GFAP) detection, transferred onto a PDVF membrane, blocked with 5% milk, and analyzed with antibodies against MBP (1:400; Millipore, Billerica, MA) or GFAP (1:1,000; Millipore). Horseradish peroxidase (HRP)-coupled donkey anti-rabbit or sheep anti-mouse (1:1,000; GE Healthcare Life Sciences, Piscataway, NJ) were used as secondary antibodies. Binding was detected by chemiluminescence using Amersham ECL Prime Western blotting reagent (GE Healthcare Life Sciences).
ImageJ software (NIH, Bethesda, MD) was used for pixel summation of individual bands, and pixel intensities were corrected for background. MBP and GFAP intensities were standardized to the corresponding β-actin bands. MBP and GFAP expression in total brain lysates from brain slices collected at the level of the middorsal hippocampus were compared to the HII group.
Immunohistochemistry
Coronal paraffin sections (6 μm) were incubated with anti-MBP (1:200; Millipore), anti-GFAP (1:500; Millipore), anti-caspase3 (CASP3; 1:50; Abcam, Cambridge, United Kingdom), and anti-ionized calcium-binding adaptor molecule 1 (IBA1; 1:100; Abcam) antibodies followed by a peroxidase-labeled polymer antibody (Dako, Glostrup, Denmark). Binding was visualized with the EnVision+System-HRP (Dako) and diaminobenzamidine. Finally, slides were counterstained with cresyl violet (MBP and IBA1 staining) and luxol fast blue (GFAP staining). To detect hWJ-MSC, an anti-human nuclei antibody (hnuclei, 1:100; Millipore) was used followed by an IgM Alexa Fluor 594 antibody (1:200; Thermo Fisher Scientific). For fluorescent double staining, anti-MBP (1:200; Millipore) and anti-SMI31 (1:1,000; Covance, Princeton, NJ) antibodies were combined. The detection was made with an IgG Alexa Fluor 488 antibody and an IgM Alexa Fluor 594 antibody (1:200; Thermo Fisher Scientific). To detect proliferating cells, an anti-Ki67 (1:20; BD Biosciences, Franklin Lakes, NJ) antibody was used and the detection was made with an IgM Alexa Fluor 594 antibody (1:200; Thermo Fisher Scientific).
The area of MBP and GFAP staining was used as a measure of the quantification of myelin or astrogliosis in three serial sections at the level of the middorsal hippocampus in a region of interest from the contralateral and ipsilateral hemispheres. This region encompassed the supracallosal radiation, the underlying corpus callosum and the rhinal sulcus. The area of MBP- and GFAP-positive staining was quantified using ImageJ [15]. The difference between contralateral and ipsilateral hemispheres was then determined. IBA1 immunoreactivity was expressed by the percentage of the whole image area containing IBA1 immunostaining. CASP3- and Ki67-positive cells were counted and averaged from three visual fields in the white matter. All images were obtained with a DM6000 B microscope (Leica Microsystems, Wetzlar, Germany).
Statistical analysis
All the values were calculated with a confidence interval of 95% and the comparison of the results was performed using GraphPad Prism (version 6.00d; GraphPad Software, Inc., San Diego, CA). The statistical analyses of the in vivo data were performed by one-way analysis of variance (ANOVA) followed by Dunnett's multiple comparison test. Each group was compared to the HII group. Outliers were detected using the Grubbs test (GraphPad Software, Inc.). A probability level of P < 0.05 was set as statistically significant. All data are expressed as mean ± standard error of the mean.
Results
Intranasally delivered hWJ-MSC reach the brain
To confirm the migratory capacity of hWJ-MSC after intranasal application, rat pups were sacrificed 1 day after transplantation. We detected human cells in the olfactory bulb, and in regions more distant from the application site, mainly the hippocampus (Fig. 1).
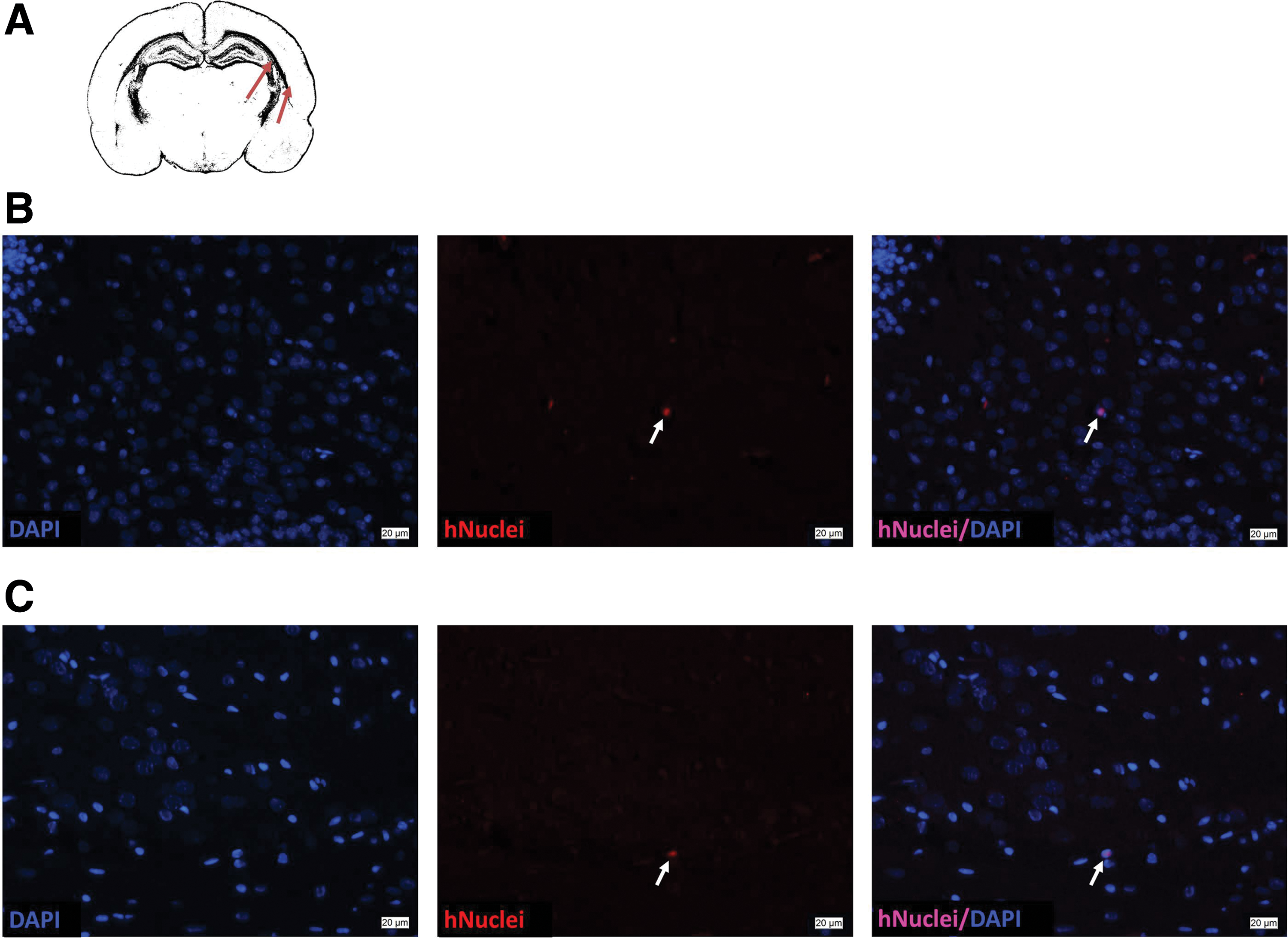
Human nuclei positive hWJ-MSC detected in the subcortical white matter at the level of the middorsal hippocampus. Coronal sections (6 μm) were incubated with a mouse anti-human nuclei antibody followed by an IgM Alexa Fluor 594 (middle). Nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI) (left). Few hWJ-MSC (white arrows) were detected in the subcortical white matter of rats treated with hWJ-MSC (right).
Effects of HII insult on myelination
HII resulted in a hypomyelination of the ipsilateral side when compared to contralateral side or control animals (P < 0.05). Myelin sheaths in damaged animals showed an impaired development and disrupted structure. Damaged animals had significantly higher levels (P < 0.01) of MBP expression after treatment than the untreated HII animals and myelination with a normal appearance similar to undamaged control animals (Fig. 2A). These results were further confirmed by western blot (Fig. 2B).
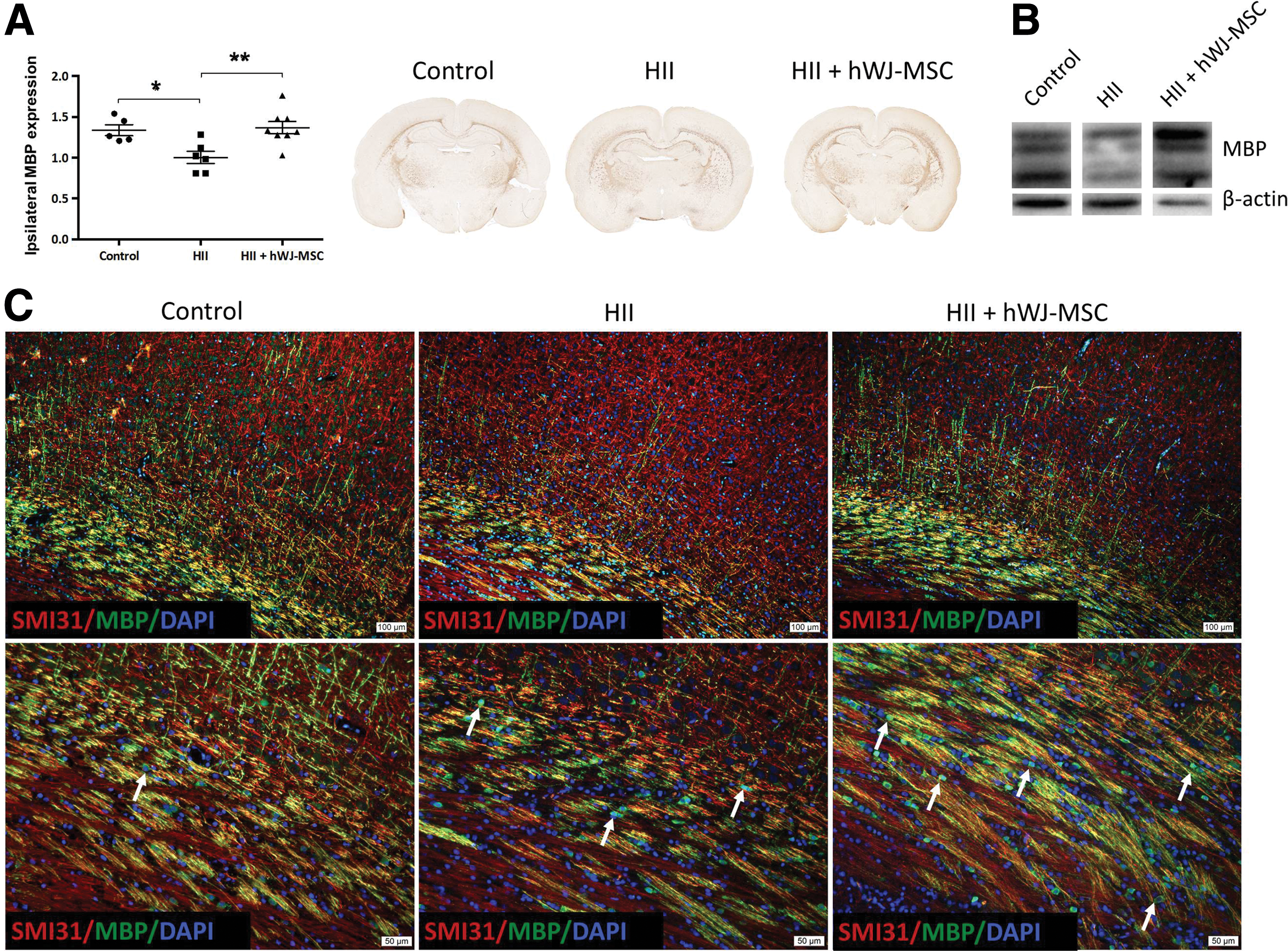
Effect of hWJ-MSC on myelination.
To verify whether axons were less myelinated in HII exposed animals, we stained myelinated axons with SMI31 and MBP. We detected a marked reduction of axonal myelination in damaged animals without axonal degeneration (Fig. 2C, Supplementary Fig. 1). Further, after injury, we observed MBP-positive cells in a linear and sequential arrangement within the white matter tracts that displayed a strong fluorescent signal mainly in and around the perikaryon, but thin and weekly positive cell processes. In control animals, these cells were only very rarely detected (Fig. 2C). In treated pups, observations similar to the injured ones could be made, but with cell processes that were thicker, more ramified, and strongly positive for MBP. Finally, numerous Ki67-positive cells were observed in the white matter tracts of the HII exposed and hWJ-MSC treated animals. The Ki67-positive cells, though, did not co-stain with MBP-positive cells (Fig. 3).
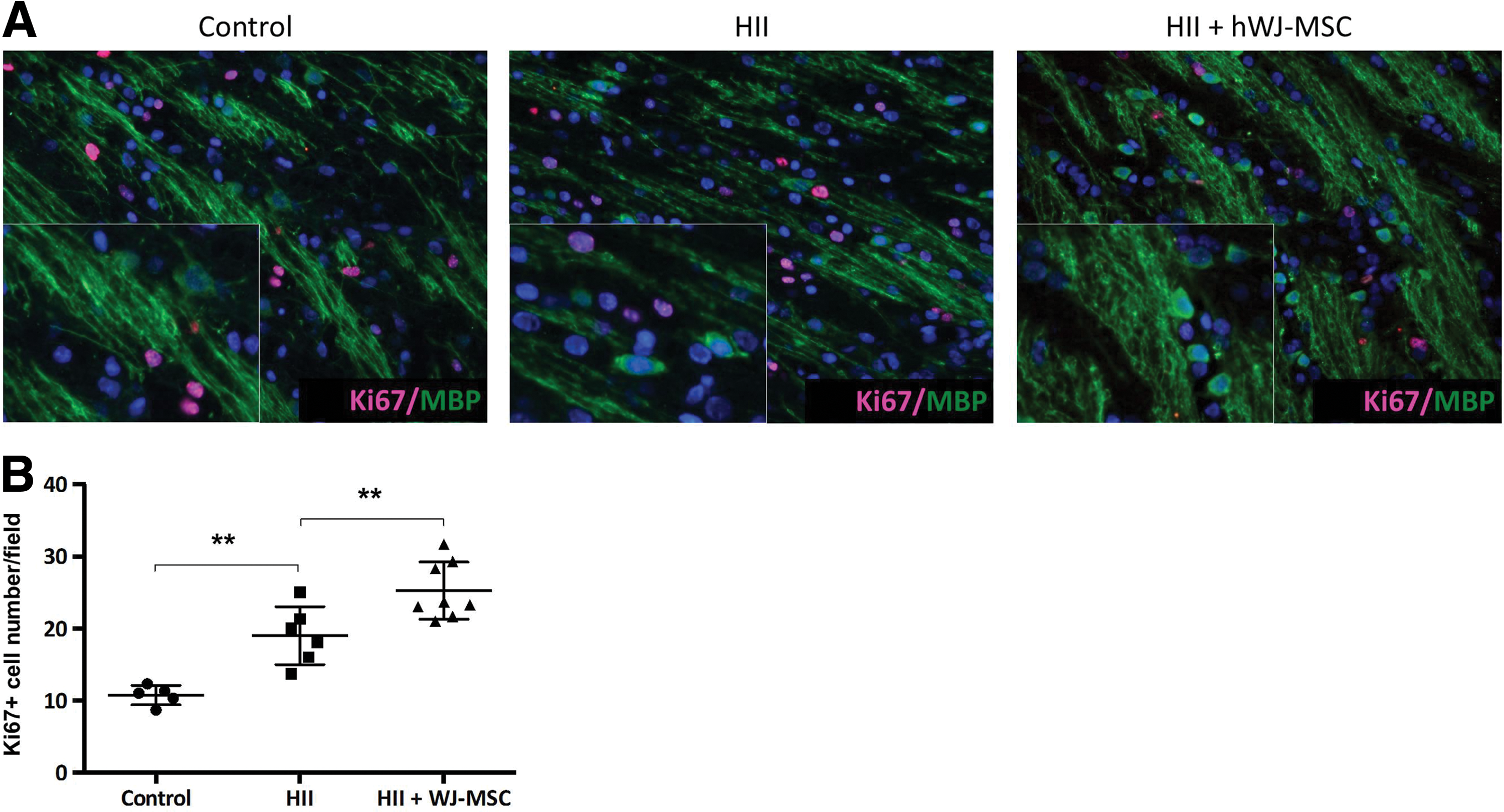
Morphology of oligodendrocytes after HII and HII+WJ-MSC.
hWJ-MSC decrease astrogliosis and microgliosis
Given the importance of glial response after brain injury, we stained for GFAP (astrocytes) and IBA1 (microglia). The expression of GFAP was significantly higher in the HII group (P < 0.05) than in the control group (Fig. 4A). Astrocytes had larger cell bodies with thicker processes in the damaged animals (Fig. 4C). The group receiving the hWJ-MSC showed lower expression levels. Western blots of GFAP displayed equal results (Fig. 4B).
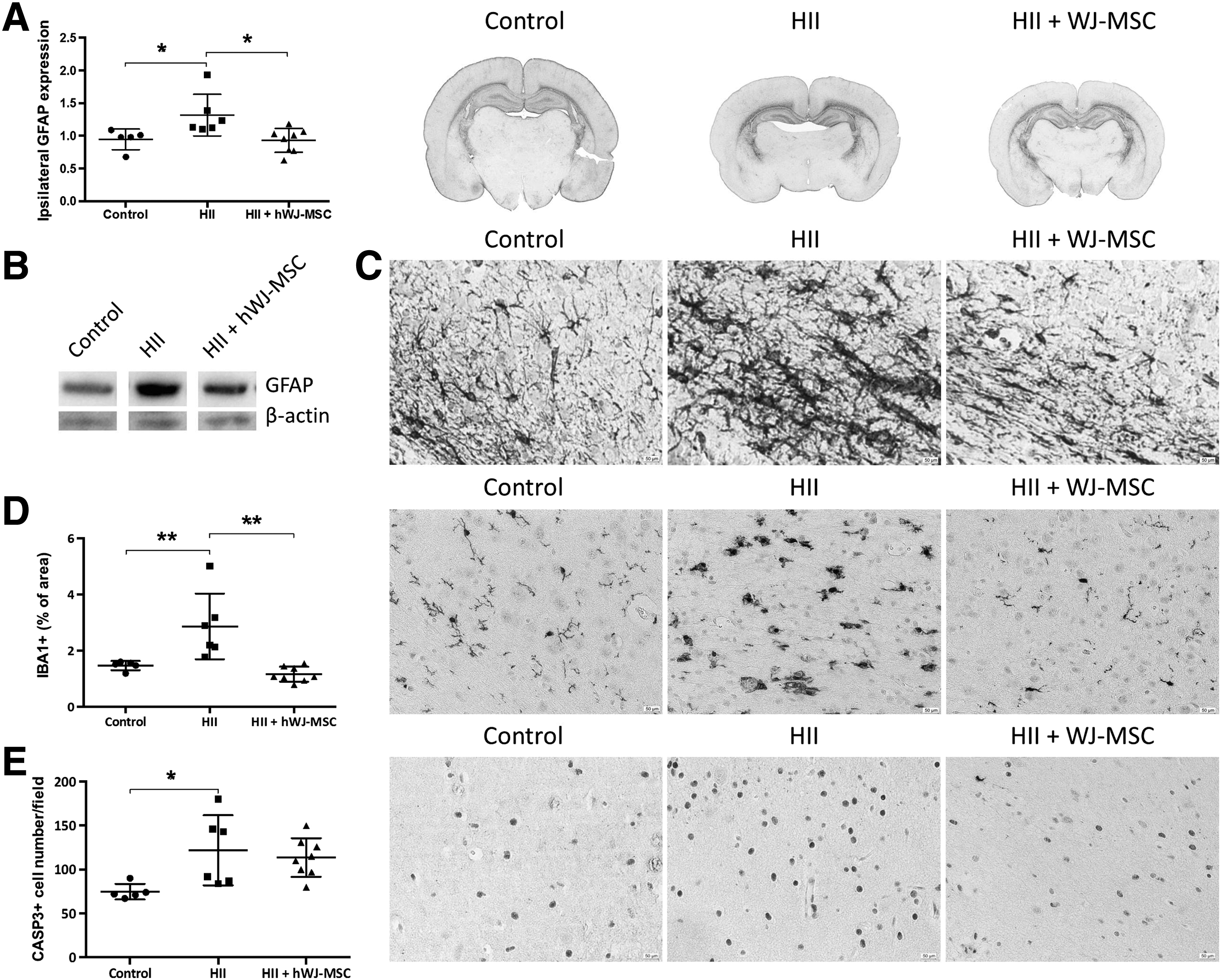
Attenuated astrogliosis and microgliosis.
We detected higher percentages of IBA1 immunostaining, and higher amounts of IBA1-positive cells with round cell bodies and reduced processes after injury (Fig. 4D). Transplantation of hWJ-MSC resulted in a significant (P < 0.05) reduction of the microglial response, as only a few activated microglia could be found in treated animals (Fig. 4D).
Higher numbers of caspase3-positive cells could be detected (P < 0.05) in the HII exposed animals compared to control animals. The hWJ-MSC group did not show significantly lower caspase3 expression (Fig. 4E).
Damage diminishes preOLs population and neurotrophic factor expression
To further elucidate hWJ-MSC's protective mechanisms on oligodendrocyte survival, we tested mRNA expression of the most common genes representative for preOLs. The expression of preOLs marker genes (Cspg4, P < 0.05; Pdgfra, P < 0.01; S100b, P < 0.001) was significantly decreased after injury compared to controls. Treatment with hWJ-MSC could not prevent the loss of preOLs significantly, although trends of amelioration could be seen (Fig. 5A).
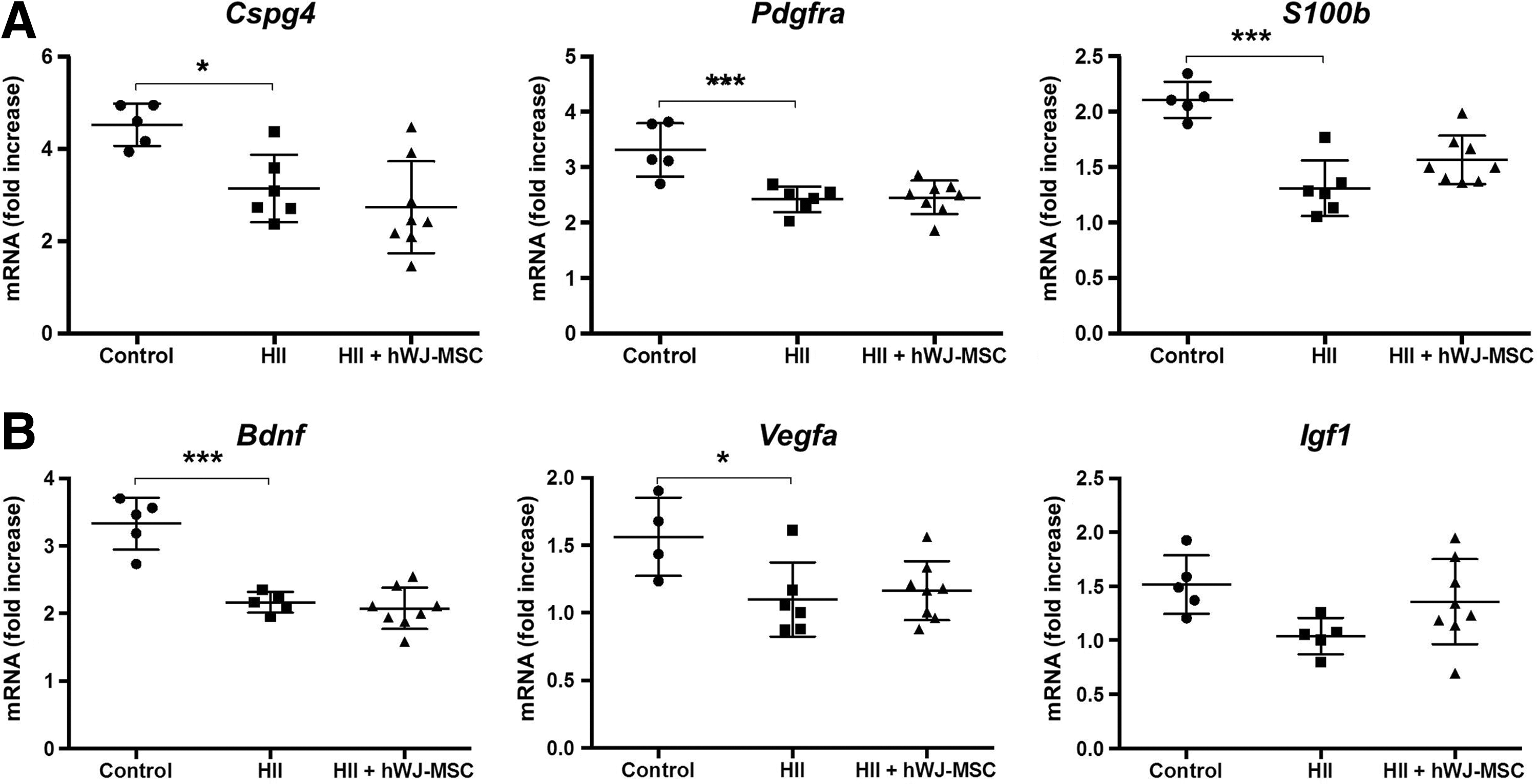
White matter injury reduces expression of preOLs markers and neurotrophic factors.
The expression of important neurotrophic factors involved in myelination, remyelination and oligodendrocyte proliferation, survival, and differentiation were assessed to better understand how hWJ-MSC might support the regenerative potential of the brain. Bdnf (P < 0.001) and Vegfa (P < 0.05) were significantly downregulated in damaged pups compared to controls. There was a tendency of increased expression in the rats that received treatment. Although not statistically significant, Igf1 expression was upregulated in the hWJ-MSC group and downregulated in the HII group (Fig. 5B).
Discussion
Our study shows that the combination of hypoxia-ischemia and inflammation in premature rat pups results in diffuse WMI with decreased myelination and numbers of preOLs, mimicking closely today's clinical cases of preterm WMI. We further demonstrate that intranasal delivery to the preterm brain is feasible and that already a low quantity of cells can reduce hypomyelination, even though the loss of preOLs cannot be prevented.
Oligodendrocytes and preOLs are the cells most susceptible to HII damage. Several studies have shown that especially preOLs are very sensitive to this type of injury, whereas more mature and very early progenitors are more resistant [16,17]. Moreover, it is thought that the environment resulting from HII injury hinders damaged and newly generated oligodendrocytes to fully regain their functions and consequently to efficiently remyelinate the brain [4,18]. Seven days after HII damage at postnatal day 12, our rat pups displayed similar pathophysiological changes as described in the literature, given that they had low levels of preOLs compared to control animals and an overall hypomyelination. The arrested or less-myelinating mature oligodendrocytes, characterized by their bright MBP signal in the perikaryon and weakly positive extensions, detectable all along the subcortical white matter tracts (see Figs. 2C white arrows and 3A inset) were almost or completely absent in control animals. Further, they did not co-express Ki67, meaning that these oligodendrocytes were not proliferating. Treated animals had many of these MBP-positive Ki67-negative cells, but in contrast to the arrested oligodendrocytes seen in the HII animals, these cells showed bright, thick, and extensive MBP-positive cell processes, pointing toward myelinating oligodendrocytes (Figs. 2C and 3A).
Given that treated animals showed significantly higher levels of MBP as assessed by IHC and WB and their oligodendrocytes appeared to be no longer arrested (at least in part) when compared to HII animals, we hypothesize that hWJ-MSC might support maturation of oligodendrocytes and attenuate the negative signals hindering oligodendrocytes to remyelinate the injured white matter tracts.
Segovia et al. [7,19] observed that after a transient short-time depletion of preOLs caused by hypoxia-ischemia, there was a strong proliferative response of the remaining preOLs as a way to repopulate the lost oligodendrocyte lineage cells. Nonetheless, the authors further realized that those cells were not able to mature, which led to the arrested maturation hypothesis mentioned above. By changing the environment hindering the maturation of the preOLs, these cells would be the source of newly generated myelinating oligodendrocytes.
We could not observe an increase of the preOLs' gene expression in the HII damaged animals (with or without hWJ-MSC treatment) compared to control but rather a down-regulation at 1 week after induction of damage. This discrepancy of results may be due to a number of experimental differences like the induction of damage (with or without LPS, O2 concentration, duration of hypoxia), the brain area analyzed (midseptal nuclei compared to middorsal hippocampus, whole hemisphere vs. specific region), gene expression compared to protein expression and the time point of brain collection.
Despite the fact that we could not observe a proliferation of preOLs that may potentially develop into mature oligodendrocytes, other oligodendrocyte lineage cells that are more resistant to HII damage could also serve as a pool for myelinating oligodendrocytes. For instance, we did not see any significant difference in galactocerebroside (Galc) expression, a marker of immature oligodendrocytes, between the groups (data not shown). So even though the treatment with hWJ-MSC could not recover the amounts of preOLs, they still might have changed the microenvironment of the brain allowing other oligodendrocyte lineage cells (e.g., immature oligodendrocytes) to continue their maturation and finally support the remyelination capacities of the brain.
We believe that HII injury induces a strong loss of preOLs (even if at a later time point they might proliferate) and creates an environment that negatively affects maturation of all stages of oligodendrocyte lineage development including mature nonmyelinating oligodendrocytes that finally leads to the well-characterized hypomyelination.
Trophic factors, along with extracellular matrix (ECM) proteins secreted by hWJ-MSC, have been discussed as possible mediators for the positive effects on myelination. From the literature, and from protein analyses done by the authors (unpublished data), we know that hWJ-MSC produce factors like BDNF, VEGFA, IGF1, interleukins (IL-6 and IL-10), and many others [20]. These factors have been reported to be involved in neuroregeneration and remyelination [21,22].
We, therefore, decided to study the expression of some of these neurotrophic factors. The results obtained from the PCR analysis indicated that IGF1 secretion by hWJ-MSC might also be involved in the reduction of the damage, given that Igf1 mRNA expression levels are higher in the hWJ-MSC treatment group compared to the untreated HII animals, although it did not reach statistical significance, likely due to the low number of animals. IGF1 is known to be responsible for oligodendrocyte survival, proliferation, and differentiation and to stimulate myelin synthesis in vitro and in vivo [23]. VEGFA and BDNF expressions remained unchanged after treatment.
We believe that the amount of cells delivered was not enough to significantly change the levels of neurotrophic factors in the brain and we have preliminary data showing that higher amounts of intranasally delivered hWJ-MSC indeed significantly increase the expression of these factors. Our data indicate that VEGFA and BDNF are not essential for damage mitigation per se, but, given their known beneficial effects [21,22], probably helpful for regeneration.
Moreover, we postulate that the secretion of ECM proteins like laminins by hWJ-MSC may create a niche that is beneficial for oligodendrogenesis [24,25]. Further, hyaluronic acid that is produced by reactive astrocytes and that accumulates after injury in the white matter and inhibits preOLs differentiation and myelination [26,27], could hypothetically be modulated by MSC changing its negative effects and thus facilitating preOLs maturation [28].
In a model of term hypoxic-ischemic brain injury, Donega et al. found that a dose of 500,000 cells was sufficient to prevent hypomyelination and that higher doses were not beneficial [15]. Based on that, we decided to treat our rat pups with a dose of 600,000 cells. We found low numbers (<0.005%) of hWJ-MSC mainly in the olfactory bulb and very few cells in more distant regions like the hippocampus. It appears that this low number of grafted cells in the brain is sufficient to prevent hypomyelination and massive microglial activation, supporting the hypothesis that they may act through secretion of trophic factors. Other studies with peripherally administered MSC have also shown that low amounts of grafted cells in the brain are already sufficient for neuroprotection [29].
Another possible way how hWJ-MSC may achieve their effects is through peripheral immunomodulation. After intranasal administration, not all the cells end up in the brain, but also in other organs like the spleen [30]. MSC have been shown to modulate peripheral inflammatory responses and consequently reduce the invasion of immune effector cells in the brain and thus helping to reduce brain damage [29,31]. So, even if only a small amount of cells reaches the brain, the ones staying in the periphery may contribute as well to the reduction of brain damage. We hypothesize that intranasally delivered hWJ-MSC have a dual immunomodulatory mode of action, targeting microglia in the CNS and immune effector cells in the periphery. This immunomodulatory action might be a major contributor of damage mitigation and brain microenvironment modulation.
Nonetheless, even if low amounts of cells might be sufficient to mitigate HII brain damage, we believe that by optimizing the numbers of delivered cells and the frequency and time points of administration, we might be able to obtain even better results regarding the engraftment and neuroprotective capacities of hWJ-MSC. Intranasal delivery itself should be improved to enhance cell engraftment. hWJ-MSC could be grown on chitosan scaffolds or on gels before delivery to increase nasal residence time of the cells due to the mucoadhesive properties of those substances [32,33]. Moreover, the application of a vasoconstrictive substance shortly before cell delivery might limit the uptake of the cells into the systemic circulation and consequently facilitate nose-to-brain transport [34]. Finally, we will also perform behavioral experiments to analyze the long-term effects of damage and treatment.
In conclusion, our clinically relevant animal model of preterm WMI with a combined HII insult not only results in decreased hypomyelination and reduced preOLs expression, but also in reduced expression of important neurotrophic factors. Intranasal delivery of hWJ-MSC is able to revert severe hypomyelination and reduce microgliosis. The reduction of the microglial response might be responsible for the amelioration after treatment.
Thus, intranasal administration of hWJ-MSC appears to be a very promising approach to treat WMI in preterm infants. Still, it should be taken into account that nose-to-brain transport in rodents is expected to be more pronounced than in humans due to the anatomical differences between species. For instance, the nasal olfactory mucosa (the entering gate for cells to reach the brain) covers ∼50% of the total nasal epithelium in rats compared to only 5% in humans [35,36]. Further studies should address the cell dose to achieve a maximal recovery and investigate whether multiple administrations are able to further improve recovery. The right quantity of cells is probably a decisive factor for this treatment approach to advance into clinical trials.
Footnotes
Acknowledgments
We are thankful to midwives and physicians of the Department of Obstetrics of the University Hospital Bern for their help in collecting umbilical cords. This work was supported by Cryo-Safe, Switzerland, Mobiliar Jubiläumsstiftung, Switzerland and the Eagle Foundation, Crans-près-Céligny, Switzerland.
Presentation in a conference: 35th Annual Meeting of the Society for Maternal and Fetal Medicine, San Diego, CA, 2015.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.