
Abstract
Graphene represents one of the most interesting additions to the tissue engineering toolbox. Novel graphene-based composites are required to improve the beneficial graphene properties in terms of tridimensional polymeric structure, conferring a higher mechanical strength and favoring the differentiation of human mesenchymal stem cells. Here, we have demonstrated in a wide range of composite combinations, the successful use of graphene and silk-fibroin constructs for future bioengineering applications in the field of clinical regenerative dentistry using human periodontal ligament stem cells. Our results provide exciting new data for the development of suitable scaffolds that allow good cell engrafting, preservation of cell viability and proliferation, promotion of spontaneous osteoblastic differentiation, and importantly, stimulation of a higher cementum physiological synthesis than using other different available biomaterials.
Introduction
I
SF is a well-known biocompatible biomaterial with a beta-sheet molecular structure, conferring an excellent combination especially with film format graphene. On the contrary, human dental stem cells, including dental pulp stem cells, stem cells from human exfoliated deciduous teeth, periodontal ligament stem cells (hPDLSCs), dental follicle progenitor stem cells, and stem cells from apical papilla, have been reported to have a high phenotype plasticity demonstrating their pluripotent potential, allowing their use in cell therapy and regenerative medicine. Importantly, hPDLSCs represent a novel stem cell population able to differentiate into both cementoblast cells and collagen-forming cells, as in vitro and preclinical studies have previously evidenced. Their phenotypic characterization have demonstrated high expression levels of the mesenchymal stem cell (MSC) markers, CD105, CD73, and CD90, but a lack of expression of the hematopoietic markers, CD45, CD34, CD14, CD19, or HLA class II. Recent data have demonstrated that the use of nanotechnology is a promising issue for periodontal ligament bioengineering [20 –22].
The aim of this study was to assess the effect of different SF-GO nanoconfigurations as a scaffolding support for hPDLSCs. Accordingly, we have optimized the parameters of fabrication of these composite films, using different ratios of the two materials (SF and GO) and also varying the graphene oxidation status. Furthermore, the hPDLSC differentiation into osteo/cementoblast was explored for each of these SF-GO composite films.
Materials and Methods
SF processing and film preparation
Cocoons of Bombyx mori were obtained from silkworms reared in the sericulture facilities of the IMIDA (Murcia, Spain), and GO water dispersion (4 mg/mL) was provided by Graphenea (San Sebastian, Spain). Forty-eight-well tissue culture plates (Nunclon™; Thermo Fisher Scientific, Waltham, MA) were used to prepare different films composed of GO, reduced GO (rGO), SF, SF plus GO, or rGO at different ratios and configurations, as described below. After handling, all tissue culture plates were sterilized by immersion in 70% v/v ethanol for 10 min and washed twice with phosphate-buffered saline (PBS) before cell seeding.
To make GO films, a GO water dispersion (250 μL/well) at 4 mg/mL was evaporated at room temperature into 48-well tissue culture plates, leading to the formation of brown films composed of 1 mg of GO, covering the bottom of each well.
For films, including SF in their compositions, cocoons of silkworms were chopped in four or five pieces and boiled in 0.02 M Na2CO3 for 30 min to remove the glue-like sericin proteins. Then, raw SF was rinsed thoroughly with water and dried at room temperature for 3 days. The extracted SF was dissolved in 9.3 M LiBr (Acros Organics, Geel, Belgium) for 3 h at 60°C to generate a 20% w/v solution that was dialyzed against distilled water for 3 days (SnakeSkin Dialysis Tubing 3.5 kDa MWCO; Thermo Fisher Scientific) with eight total water changes. The resultant 6% w/v SF dissolution was recovered, filtered, and stored at 4°C for no longer than 30 days.
Pure SF films were obtained by casting 200 μL of 1.5% w/v SF aqueous dissolution per well (3 mg/well). To manufacture the composite films of SF and GO, different amounts of SF solution (1.5% w/v) and GO water dispersion (4 mg/mL) were blended and evaporated at room temperature inside each well of the tissue culture plates. These films were named as GO:SF (ratio expressed in dry weight). GO:SF (3:1) films were produced using a mixture of 750 μL of GO water dispersion (4 mg/mL) and 66.7 μL of SF water solution (1.5% w/v) leading to the formation of films composed of 3 mg of GO and 1 mg of SF. GO:SF (1:1) films were made by means of evaporation of 500 μL of GO water dispersion (4 mg/mL) mixed with 133.3 μL of SF water dissolution (1.5% w/v), being the final composition of each film, 2 mg of GO and 2 mg of SF. A blend of 250 μL of GO water dispersion (4 mg/mL) and 200 μL of SF water solution (1.5% w/v) was evaporated to produce GO:SF (1:3) films, whose composition was 1 mg of GO and 3 mg of SF per film. Once dried at room temperature, these materials, including SF in their composition, were annealed within a water filled desiccator in vacuum conditions for 24 h to produce water insoluble films.
Two different layer by layer composite films were produced during the same experiments, to test the effects of GO and SF in direct contact with the cells and the potential use of these materials in biomedicine, in which the presence of the SF would be the key to their appropriate handling. This way, one of the layer by layer composite films, named as GO/SF, was produced by means of evaporation of 200 μL of 1.5% w/v SF aqueous dissolution (3 mg/well) with a subsequent step of water annealing (as described previously) and a second step consisting of casting 250 μL of GO water dispersion at 4 mg/mL, leading to the formation of a continuous second layer of GO (1 mg) that constituted the contact surface for the cells. The second layer by layer composite film, named as SF/GO, was produced in the same way but with its layers in inverted positions. In this case, the SF surface was the layer in contact with the cells.
All these composite films described before were used for the cell culture experiments, but some of them were immersed in an ascorbic acid solution (20 mM) for 3 h at 70°C, following the previous method described by Fernández-Merino et al. to reduce the GO present in their composition, leading to the formation of black films of rGO [16]. Some pure SF films were also treated with the ascorbic acid solution to test the influence of this step on the properties of them as biomaterials. To differentiate from films without ascorbic acid treatment, these materials were named as rGO, rSF, rGO:rSF (3:1), rGO:rSF (1:1), rGO:rSF (1:3), rGO/rSF, and rSF/rGO films, respectively, and were also used for cell culture experiments.
Isolation and culture of hPDLSCs
Human periodontal ligaments (hPDL) were obtained from impacted third molars from 10 healthy subjects. Donors provided written informed consent according to the guidelines of the Ethics Committee of our Institution. hPDL was scraped from the middle third region of the root surface. After their extraction, hPDL was washed with Ca2+/Mg2+-free Hank's balance salt solution (Gibco, Gaithersburg, MD), and subjected to a collagenase-A digestion (3 mg/mL; Sigma-Aldrich, St. Louis, MO) for 1 h at 37°C. Afterward, cells obtained after this enzyme digestion were seeded into 25-cm2 plastic culture flasks (BD Biosciences, San Diego, CA) in low-glucose Dulbecco's modified Eagle's medium supplemented with penicillin/streptomycin (PAA Laboratories, Pasching, Austria),
Scanning electron microscopy
Scanning electron microscopy (SEM) was used to analyze cell adhesion, spreading, and proliferation on the surface of each composite. The different films were placed at the bottom of wall-removable four-well plates. Then, hPDLSCs were seeded on each composite film at a density of 7 × 103 cells/cm2. After incubation for 10 days, the culture medium was removed and hPDLSCs were washed with PBS and fixed with 3% glutaraldehyde in 0.1 M cacodylate buffer for 1.5 h at 4°C. Subsequently, they were rinsed again and postfixed with osmium tetroxide for 1 h before being dehydrated in a series of graded ethanol solutions (30%, 50%, 70%, and 90% v/v). Final drying was performed by the critical-point method (CPDO2 Balzers Union). Finally, gold-coated specimens were examined by SEM.
Proliferation assay
Cell proliferation on the different biomaterials was analyzed using the MTT assay (MTT Cell Growth Kit; Chemicon, Rosemont, IL). This assay is based on the ability of mitochondrial dehydrogenase enzymes to convert the yellow water-soluble tetrazolium salt 3-(4,5-dimethyl-thiazol)-2,5-diphenyl-tetrazolium bromide into colored compounds of formazan, whose absorbance is proportional to the amount of proliferating cells. The proliferation rate of hPDLSCs was analyzed after 4, 7, and 10 days of culture. hPDLSCs were resuspended in the complete medium at a density of 7 × 103 cells/cm2 and seeded into 48-well culture plates previously coated with the described fibroin, graphene, or fibroin/graphene composite films. Other hPDLSCs were seeded on uncoated plastic wells to be used as a control. Afterward, wells were washed twice with PBS and incubated with the MTT solution (1 mg/mL) for 4 h. Then, wells were extensively washed with PBS, and 100 μL of dimethyl sulfoxide (DMSO) was added to extract and solubilize the formazan. Absorbance at 570 nm (Abs570) was measured in an automatic microplate reader (ELx800; Bio-Tek Instruments, Winooski, VT), using the Abs690 as the reference wavelength. Each experimental condition of the assay was analyzed in quintuplicate. Afterward, data were imported to Excel software (Office 2007; Microsoft Corporation, Redmond, WA) and subjected to statistical analysis.
Analysis of expression of MSC surface markers by flow cytometry
The expression of MSC surface molecules was analyzed on cultures of hPDLSCs in passage 2–4 by flow cytometry. Briefly, cells were seeded at a density of 3.0 × 104 cells/cm2 in 48-well plates previously coated with fibroin, graphene, or fibroin/graphene composite films and cultured in the complete medium for 10 days at 37°C. Then, cells were detached using a 0.25% w/v trypsin-EDTA solution, rinsed with PBS, and incubated in the dark at 4°C for 30 min with fluorescence-conjugated specific monoclonal antibodies for CD73, CD90, and CD105 (Miltenyi Biotec, Bergisch Gladbach, Germany), which have been recommended by the International Society of Cellular Therapy (ISCT) as essential to confirm the mesenchymal phenotype of the cells [23,24]. Lack of expression of the hematopoietic markers CD14, CD20, CD34, and CD45 was also analyzed. Nonspecific fluorescence was measured using specific isotype monoclonal antibodies. Afterward, cells were acquired using a BD FACSCanto flow cytometer (BD Biosciences) and later analyzed with Kaluza analysis software (Beckman Coulter, Inc., Brea, CA).
Determination of cellular viability (Annexin V/7-AAD staining)
hPDLSCs were cultured on the different biomaterials up to 10 days, followed by double staining with PE-conjugated Annexin-V and 7-AAD (BD Biosciences). Briefly, cells were cultured, detached, washed as described above, and incubated with PE-conjugated Annexin-V and 7-AAD in Annexin-V-binding buffer at room temperature for 15 min. The percentage of live (Annexin-V−/7-AAD−), early apoptotic (Annexin+/7-AAD−), or late apoptotic and necrotic cells (Annexin-V+/7-AAD+ and Annexin-V−/7-AAD+) was analyzed in a BD FACSCanto flow cytometer. Subsequently, percentages of each population were calculated. All determinations were performed in triplicate.
RNA isolation, reverse transcription polymerase chain reaction, and quantitative polymerase chain reaction
hPDLSCs were seeded in 48-well plates coated with different fibroin and graphene composite films and cultured in the complete medium for 10 days. Importantly, a negative control (plastic) and a positive control for osteogenic differentiation (OsteoDiff®; Miltenyi Biotec) were carried out. Total messenger RNA (mRNA) was isolated by a standardized phenol–chloroform protocol using the TRIzol® Reagent (Life Technologies, Carlsbad, CA) and miRNeasy Mini Kit (QIAGEN, Limburg, Netherlands) according to the manufacturer's protocol. In brief, hPDLSCs were directly solubilized and homogenized using prechilled TRIzol Reagent and scrapper procedure. Whole cell lysates were previously centrifuged at 12,000g at 4°C for 10 min to remove insoluble material, including remaining substrate preparation and extracellular matrix (ECM) calcium deposits if any. Afterward, mRNA amount and purity were commonly quantified by NanoDrop (Thermo Fisher Scientific).
Total mRNA (0.1–0.25 μg) was reverse-transcribed into complementary DNA (cDNA; miScript II RT Kit; QIAGEN) in a thermal cycler for 1 h at 37°C (Techne TC-412; Bibby Scientific, Stone, Staffordshire, UK). Downstream quantitative polymerase chain reaction (qPCR; miScript SYBR Green PCR Kit; QIAGEN) was performed using QuantiTect® Primer Assay primers (QIAGEN) in a Light Cycler LC480 (Roche Diagnostic®, Basel, Switzerland) according to the manufacturer's recommendations. We assayed different clusters of genes coding for osteo/chondroblast-specific transcription factors (RUNX2, SOX9, and SP7/OSX), osteoblast-related proteins (BMP2, ALP, COL1A1, and BGLAP), as well as cementum-related genes (PTPLA/CAP and CEMP1). GAPDH was used as housekeeping gene. Technical triplicates were included for each sample and gene assayed. Fold-change gene expression was assessed by the 2−ΔΔCT method. All comparisons were related to plastic negative control.
Statistical analysis
Continuous variables were tested for normal distribution by the Kolmogorov–Smirnov test and compared by using t-tests, including the unpaired t-Student test or the one-way analysis of variance. The Kruskal–Wallis test and Wilcoxon/Mann–Whitney U test were used when appropriate. Statistical significance was accepted for P value <0.05. SPSS 19.0 for Windows (SPSS, Inc., Chicago, IL) was the statistical software used.
Results and Discussion
Culture of hPDLSCs
After collagenase-A digestion, the single hPDL cell suspensions were plated in 25-cm2 culture flasks and cultured at 37°C. Two days later, cells attached to the plastic bottom of the flasks displayed a typical spindle-shape fibroblast-like morphology in standard culture medium and showed single-cell-derived colony forming ability. hPDLSC cultures initially showed a heterogeneous cell morphology and turned to homogenous following subsequent serial subcultures, especially after two passages.
Analysis of the different graphene- and fibroin-based composite film topography
Graphene can be transferred into any flat or irregular-shaped surfaces, and later these graphene-coated, flexible, and supporting substrates can be easily bent into any shape required [25]. By contrast, GO is a difficult to handle weak material. To overcome this problem of handling, SF could provide many advantages to GO since it confers robustness [18] and a better handling to GO (data not shown).
Different degrees of roughness, discontinuities, and recesses were observed on the different fibroin–graphene composite surfaces by SEM (Fig. 1). Nevertheless, no topographic differences were detected between oxidized and reduced versions. In pure configurations, GO appears as a slightly cracked film with folds in the discontinuities, while fibroin had the appearance of a substantially smooth surface, accordingly observed in previous studies [26]. As expected, surface irregularities increased progressively with the amount of fibroin in different ratios of graphene–fibroin mixtures. A surface with scarce folds and discontinuities was observed in the 3:1 ratio. In the 1:1 ratio, a higher roughness and few folds were found. On the contrary, the 1:3 ratio displayed the image of several discontinuities and folded films in different depths, as previously reported [26]. Finally, bilayer configurations with fibroin in the surface presented the same aspect of pure fibroin, although the inverted configuration displayed no similarities with pure GO composites. Specifically, the surface of the bilayer GO/SF and rGO/rSF showed a high degree of roughness, with many discontinuities and folds at different levels and depths, conferring to these composites similar features to 3D scaffolds.

Scanning electron microscopy characterization of graphene- and fibroin-based composites. Composite topography. Graphene- and fibroin-based composites, both oxidized
Analysis of hPDLSC morphology and attachment on the different graphene- and fibroin-based composite films
Several authors have confirmed a relationship between substrate topography and morphology and adhesion of human MSCs (hMSCs) [27 –33]. Accordingly, we previously found that a lower cellular spreading of hPDLSCs cultured on SF composites was associated to decreased levels of F-actin filaments as well as with a slightly low but significant adherence compared to GO and GO-SF [26]. Now, we have evidenced that the flat configuration of composites with the cells in direct contact to fibroin (pure composites or bilayers with fibroin on top) significantly affects hPDLSC morphology after 10 days after cell seeding. Accordingly, cells showed a lower spreading, thus suggesting a defective adhesion mechanism to these substrates (Fig. 1). Park et al. also reported a lower cell spreading after treating MSCs seeded on collagen-coated culture dishes with anti-β1 integrin blocking antibody [34]. These findings suggest that lower spreading of hPDLSCs on fibroin composites may be closely associated to a defective or delayed β1 integrin-dependent adhesion. By contrast, hPDLSCs displayed a spindle-shaped morphology in the remaining composites containing graphene. Similarly, Nayak et al. reported that graphene did not influence the shape of hMSCs but polyethylene terephthalate and polydimethylsiloxane non-graphene-coated substrates showed cells with rounded or irregular morphology [14].
In addition, hPDLSC density of the cellularized composites was also analyzed by SEM (Fig. 1). Mainly, both oxidized and reduced versions of GO:SF (1:3) configurations displayed an extensive cellular colonization of the composite, allowing the formation of a cell monolayer covering the entire surface of the biomaterial. Not only oxidized and reduced 3:1 ratios but also configurations of the composite films displaying a lower cell spreading (SF, rSF, SF/GO, rSF/rGO) showed a very low density.
Effect of different configurations of graphene and fibroin composites on hPDLSC proliferation
hPDLSCs seeded on the different graphene and fibroin composites during a time course of 10 days after the initial cell seeding were explored for their engrafting capacity and proliferation. Accordingly, MTT proliferation rates of hPDLSCs on the different materials were measured after 4, 7, and 10 days of culture (Fig. 2).
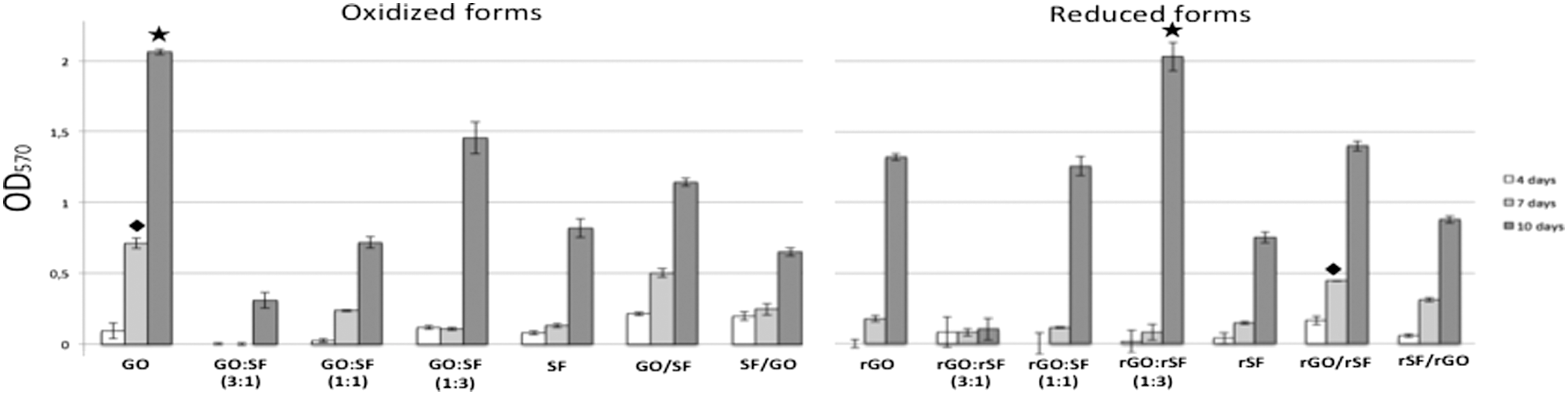
MTT proliferation assay. Data are expressed as the mean values of optical density (Abs570) ±standard deviations (n = 5). hPDLSC proliferation rate on different graphene- and fibroin-based composites at 4, 7, and 10 days after seeding. Symbols represent the highest proliferation with significant differences (P < 0.05) in composites with the same oxidation state at 7 days (diamond) and 10 days (star). Abs570, absorbance at 570 nm.
Most published works have studied how affect different ceramics or hydroxyapatite-based preparations on proliferation. Some materials such as diopside ceramics, Ca7Si2P2O16 bioceramics, zein, or multiwalled carbon nanotube/hydroxyapatite nanoparticles have demonstrated ability for supporting hPDLSC growth [35 –38]. Other previous works only compared cell proliferation rates in a small group of biomaterials, obtaining best results on substrates containing hydroxyapatite or grapheme [39 –41]. In this study, we have assessed different fibroin–graphene composites to find the best configuration regarding cell proliferation and differentiation.
To date, most of the experiments illustrating cell behavior on graphene surfaces have been conducted in glass or plastic surfaces covered by graphene monolayers, or papers made by GO flakes recovered by filtration of an aqueous solution. Fibroin is a well-known natural and implantable biomaterial and its β-sheet molecular structure confers an excellent combination, especially with film format grapheme [18]. In the present study, up to 14 different configurations of graphene- and fibroin-based biomaterials were evaluated. Three of them were used in a prior study previously reported by our group [26], obtaining similar results in the same configurations.
Importantly, the proliferation rate of hPDLSCs on the GO composite obtained after 7 days was found to be consistently higher than that found for the remaining composites, including the rGO configurations (P < 0.01). No significant differences were found in hPDLSC proliferation when compared to GO/SF with rGO/rSF bilayer, but both proliferation rates were significantly lower than those obtained on GO and higher when compared with the remaining composites (P < 0.05; Fig. 3A).
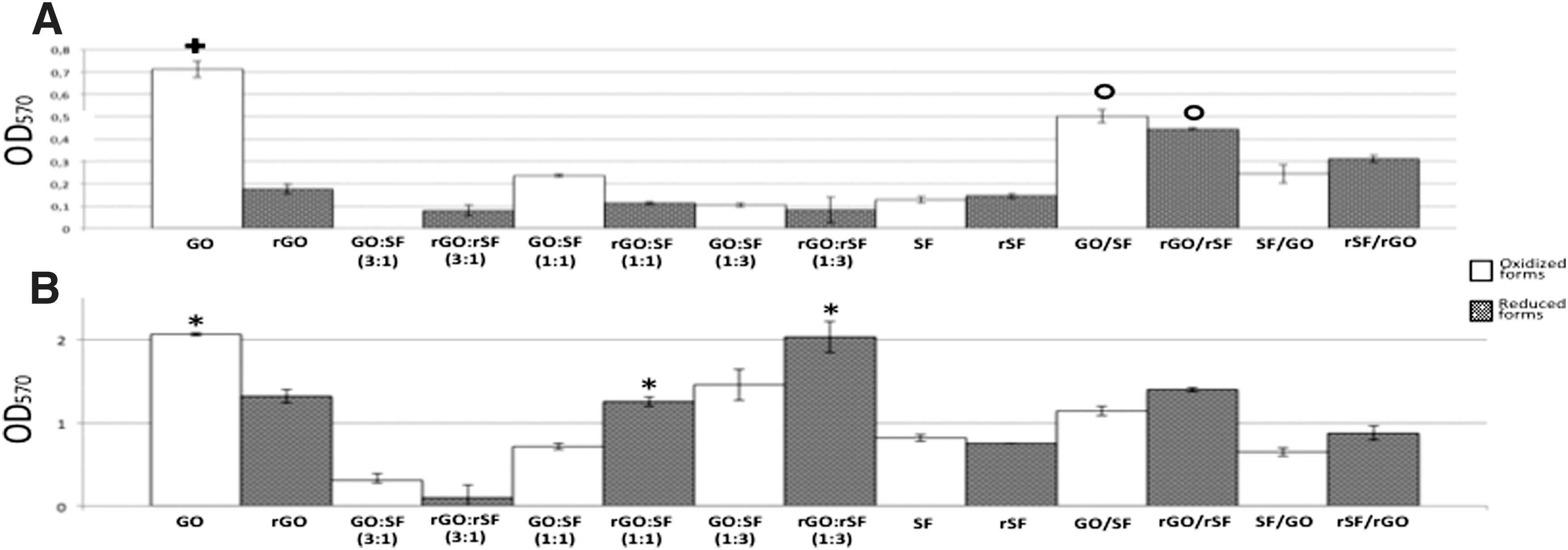
MTT proliferation assay. Data are expressed as the mean values of optical density (Abs570) ±standard deviations (n = 5).
Oxidized versus reduced composites
After 10 days of culture, we also reported significant differences on the proliferation rates obtained on some oxidized or reduced forms with the same configurations (Fig. 3B). Pure fibroin configurations (SF and rSF) were similarly prepared, although they were differently treated with ascorbic acid. No significant differences were found on their proliferation rates (Fig. 3B; P = 0.653), thus suggesting that ascorbic acid treatment may be innocuous for proliferation in our cell model. The hPDLSC proliferation rate after 10 days of growth on GO remained significantly higher than on rGO (P < 0.05). When both oxidized and reduced forms of the different ratios were analyzed, the proliferation rate in reduced configurations of 1:1 (P < 0.01) and 1:3 ratios (P < 0.05) was found to be significantly higher than the oxidized. The 3:1 ratio, both oxidized and reduced, displayed the lowest proliferation rate with statistical significance (P < 0.05). When GO and SF were together in the different ratios or bilayer configurations, cell proliferation was generally higher on reduced composites than on oxidized (Fig. 3). Taken together, these results suggested that the best biomaterials in terms of proliferation of hPDLSCs were GO and rGO:rSF (1:3).
Graphene–fibroin synergism
Previous data have shown that hPDLSCs bioengineered on 3D graphene scaffold preparations are associated with higher proliferation rates than on 2D ones [41]. Other graphene preparations based on hydroxyapatite-derived 3D structures have not shown significant differences in hMSC proliferation rates [25]. In this study, we used fibroin to confer 3D characteristics to the GO or rGO as well as to improve its handling. On one hand, GO and rGO are breakable materials when they are handled. On the other hand, direct contact with SF or rSF, as pure as in bilayer configurations, affects spindle hPDLSC morphology resulting in a lower cell spreading (Fig. 1). The hPDLSC proliferation rate on rGO:rSF (1:3) composites was the highest and statistically significant compared to the other mixtures (P < 0.05; Fig. 3B). Importantly, hPDLSC proliferation rates in mixture configurations went up as the SF dose increased and GO amounts decreased, both oxidized and reduced. Results obtained in the four proposed bilayers showed a moderate proliferation. Accordingly, no significant differences were found between GO:SF (1:1), GO/SF, and SF/GO, neither the same reduced forms (P = 0.067; Fig. 3B). Our data evidenced that GO and SF together, as oxidized as reduced, improves the limitations shown by pure biomaterials in terms of handling and proliferation, therefore using fibroin upgrades its manufacture for future clinical translational purposes.
Cytotoxicity of graphene- and fibroin-based composites
The biocompatibility of biomaterials is a prerequisite for the optimal generation of cellularized scaffolds that allow their further eligibility and use as therapeutic tools for specific clinical settings. For this purpose and for evaluating the possible cytotoxicity of the different configurations of fibroin- and graphene-based composite films, we investigated the viability of hPDLSCs cultured on the different biomaterials by measuring the binding of Annexin-V and 7-AAD, two-color flow cytometry analysis usually used to determine the cell apoptosis stage. This method allows differentiating three cell populations: live (Annexin-V− 7-AAD−), early apoptotic (Annexin-V+ 7-AAD−), and both late and necrotic cells (Annexin-V+ 7-AAD+ and Annexin-V− 7-AAD+). In a previous study, we determined that hPDLSCs cultured on SF, GO, or GO/SF (1:3) displayed only a slight initial cell death, with more than 85% of viable cells after 7 days of culture on all biomaterials [26]. Similarly to these results, the present configurations of SF and graphene combined biomaterials, both oxidized and reduced, displayed hPDLSC viability higher than 95% after 10 days of culture (Fig. 4). Thus, graphene- and fibroin-based biomaterials described here could be considered as promising biocompatible scaffolds, which used in the suitable configuration may preserve the hPDLSC proliferation, viability, and function.
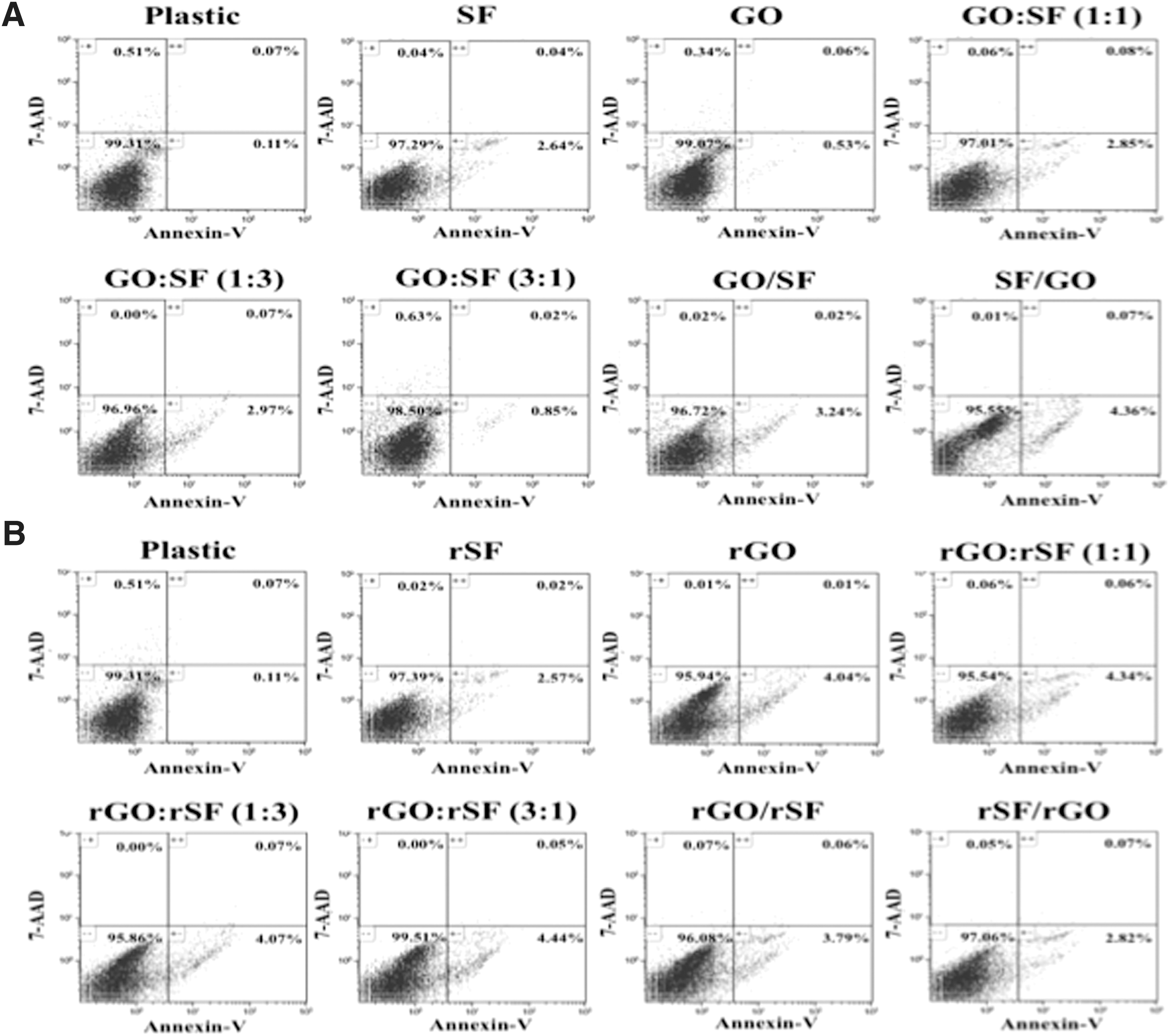
Effect of graphene- and fibroin-based composite in hPDLSC viability. hPDLSCs were cultured on oxidized
Expression of MSC surface molecules on hPDLSCs cultured on graphene- and fibroin-based composites
To confirm the mesenchymal phenotype of isolated hPDLSCs from primary hPDL cultures and to determine possible phenotypic changes after their culture on the different graphene and fibroin configurations, flow cytometry studies were carried out. In all the biomaterials tested, the MSC surface molecules CD73, CD90, and CD105 were expressed to levels >99.5%, whereas expression of the hematopoietic markers CD14, CD20, CD34, and CD45 was <5% (Fig. 5). Although the turnover of MSC markers could be slightly up- or downregulated in response to cell confluence and cell cycle, hPDLSCs cultured on graphene- and fibroin-based biomaterials for 10 days displayed mostly a significant downregulation of the expression of CD73, CD90, and CD105, mainly when the composite contains reduced graphene in its composition. Some authors have previously reported that the expression level of mesenchymal markers progressively declined in MSCs during their multilineage differentiation, including the differentiation toward osteoblast-like cells [42 –44]. These results suggest that graphene and fibroin composite films could be suitable biomaterials to be used as spontaneous inductors of osteogenic differentiation of hPDLSCs.
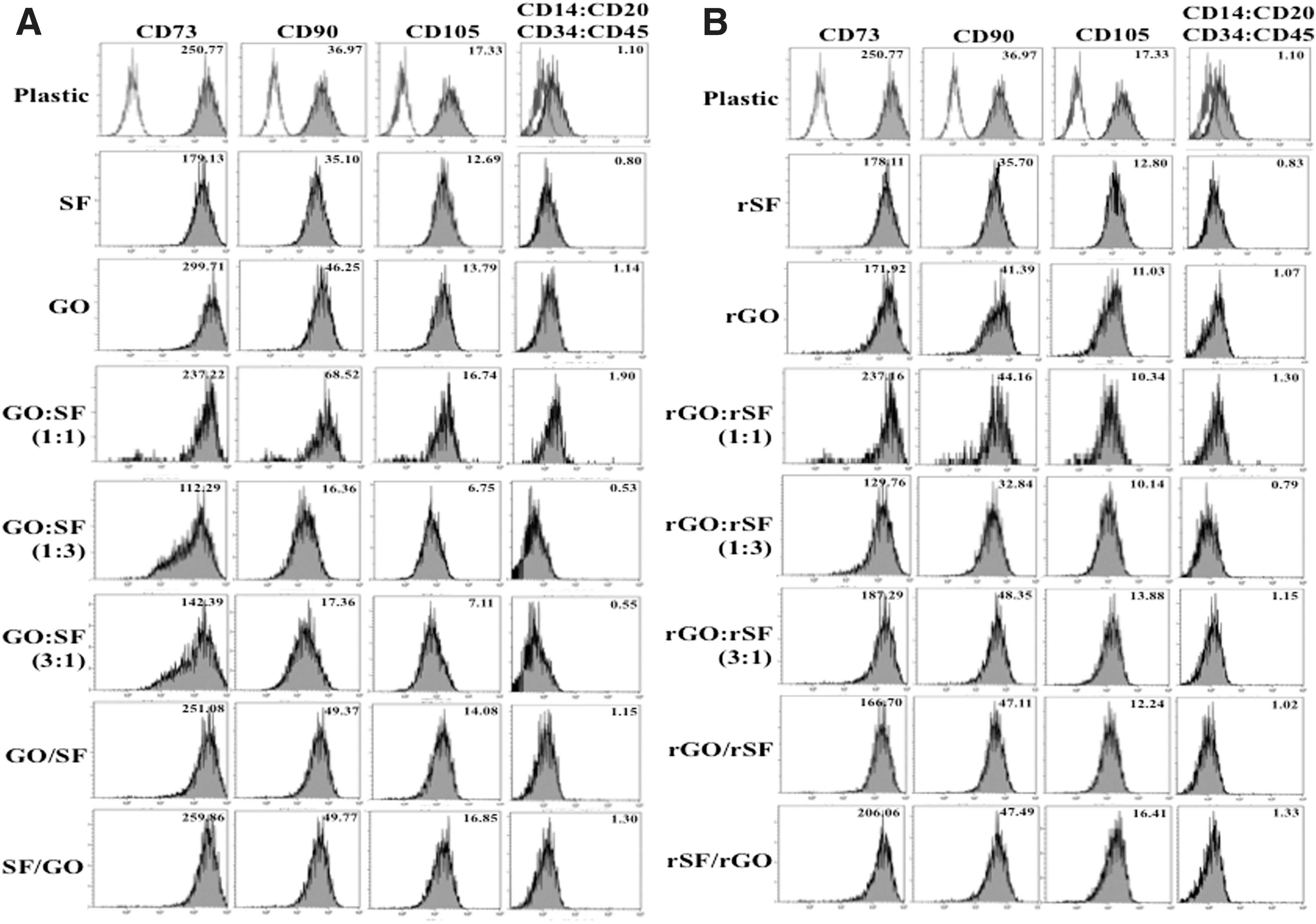
Mesenchymal phenotype analysis of hPDLSCs after culture on graphene- and fibroin-based composites by flow cytometry. hPDLSCs were cultured on oxidized
Coexpression of bone- and cementum-related factors during hPDLSC differentiation
Graphene seems to act on the differentiation of stem cells such as neural stem cells differentiating to neurons [15,45,46], MSCs to osteoblasts [14,25,47 –49], PDLSCs to osteoblasts [41], and induced pluripotent stem cells to different cellular lineages [50]. Cementum is a heterogeneously calcified tissue rendering a rigid tooth-anchoring structure and resulting from cementoblasts. Cementoblasts derive from undifferentiated MSCs residing in the connective tissue of the dental follicle. Moreover, hPDLSCs have been also demonstrated to be a source of cementoblasts or osteoblasts, thus forming cementum or alveolar bone, respectively [51 –53]. Importantly, cementum expresses certain unique molecules and factors that promote cell chemotaxis, migration, adhesion, recruitment, proliferation, and differentiation of some hPDLSCs [52,53].
Using hyperphosphatemic conditions by commercial formulations or a mixture of β-glycerophosphate, ascorbic acid and dexamethasone, it has been demonstrated the induction of hPDLSC mineralization and osteoblast differentiation. In this study, we used a commercially osteogenic differentiation medium to differentiate hPDLSCs into osteoblast-like cells as a positive control for the expression of the targeted genes. The majority of genes were not normally distributed, and therefore, they were plotted into box plot graphs (Fig. 6). Osteo/chondroblast- and cementum-related genes were assayed as described in the Materials and Methods section. hPDLSCs seeded on propylene plates and cultured in the OsteoDiff medium were found to concomitantly upregulate osteoblast (RUNX2, SP7/OSX, BMP2, BGLAP, COL1A1, ALP), chondroblast (SOX9), and cementoblast phenotype markers (PTPLA/CAP and CEMP1) compared to expression observed in hPDLSCs cultured in complete medium (Fig. 7B). Importantly, these results were in concordance with previous published results [51]. Noteworthy, after 10 days under osteogenic induction, no expression of DSPP was found, as expected for hPDLSCs (data not shown) [54]. We also analyzed the functional mineralization of these hPDLSCs differentiating into osteo/cementoblast-like cells by light microscopy, appearing calcium deposits as dark precipitates on the cell monolayer (Fig. 7A). Those results together indicated that hPDLSCs have the phenotypic plasticity and capability to differentiate into functional cementoblasts or osteoblasts through chondroblast intermediates, thus suggesting their potential use to preferably achieve a cementoblast phenotype.
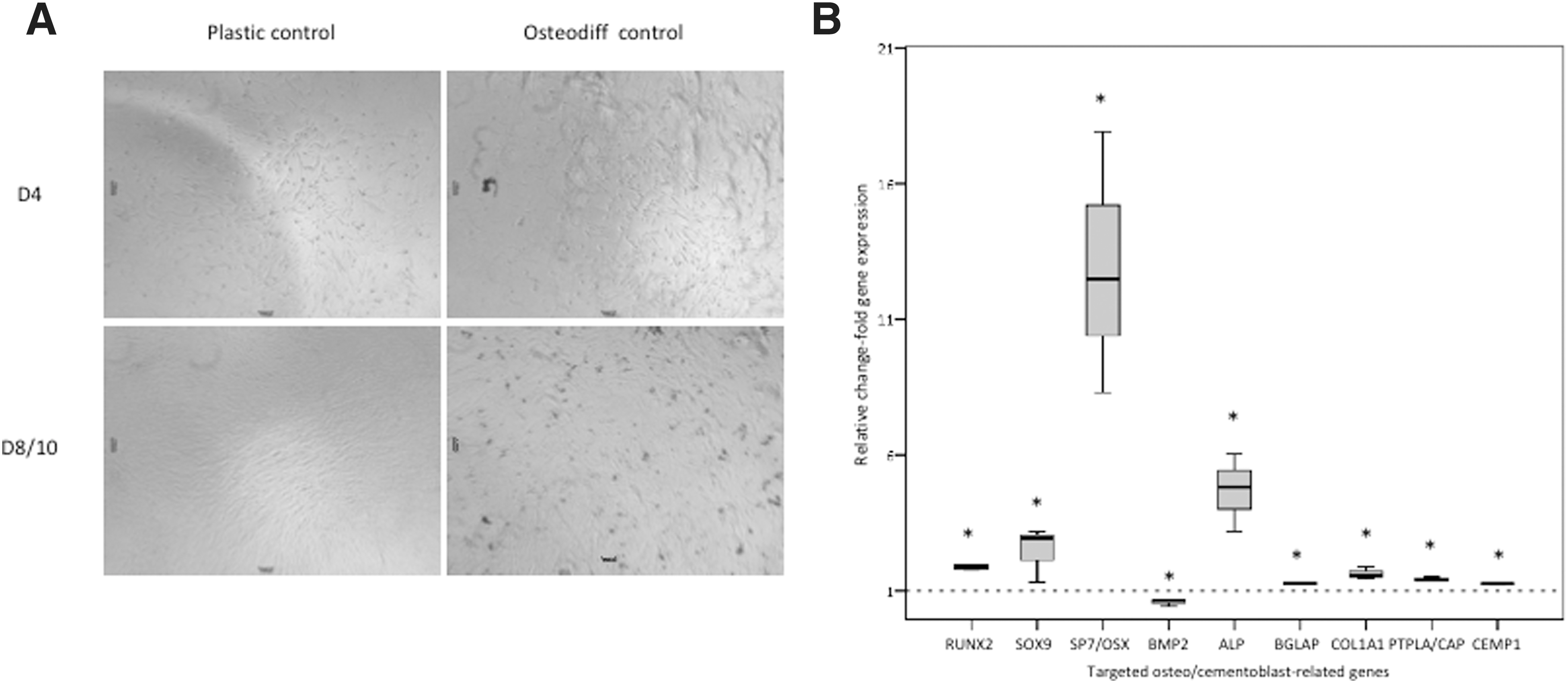
hPDLSC mineralization and differentiation into osteo/cementoblast-like cells. hPDLSCs were assessed for their phenotype plasticity and capability to differentiate into osteoblast-like cells. A commercial osteogenic differentiation medium (see Materials and Methods section) was used to promote osteoblast commitment.
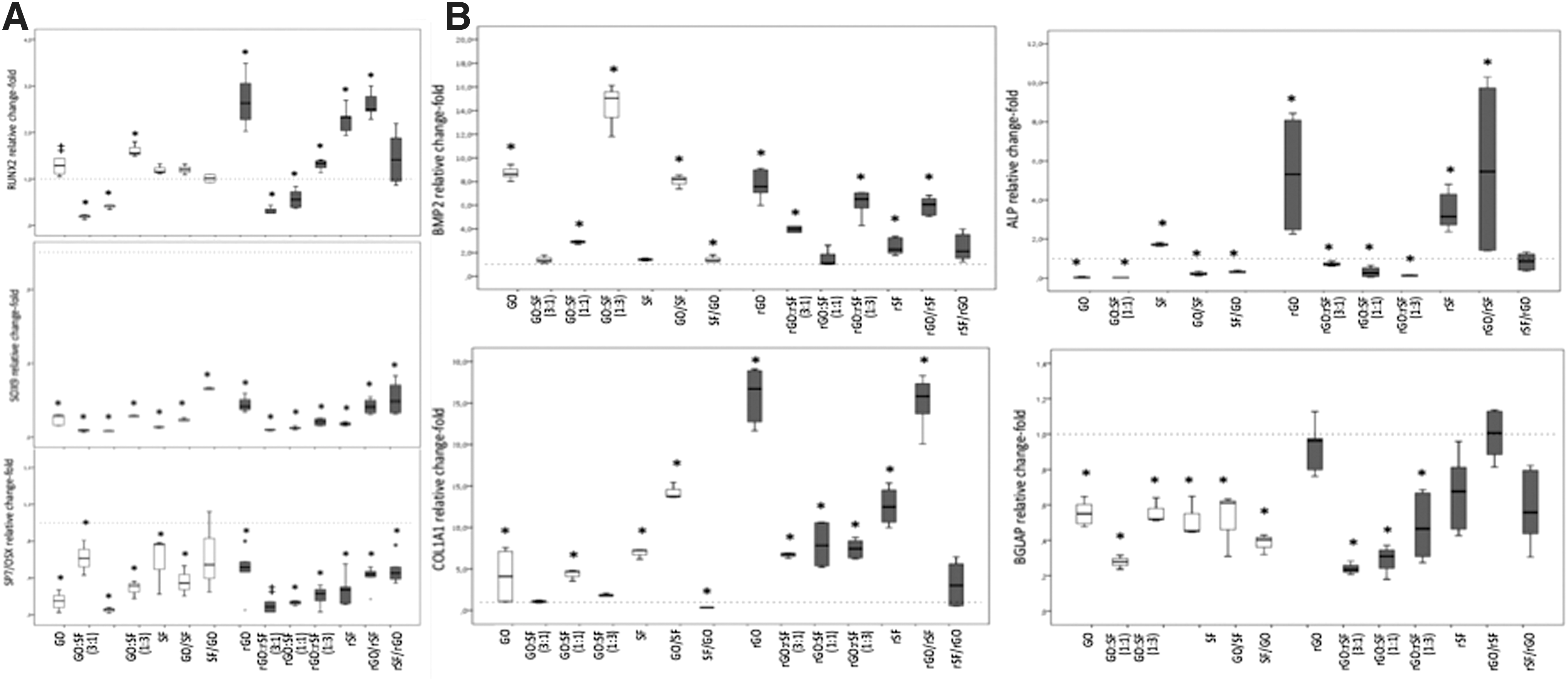
Graphene–fibroin substrates regulate the spontaneous expression of osteo/chondro-related genes in hPDLSCs after 10 days of culture.
Graphene–fibroin composites promote the overexpression of cementoblast-related genes
hPDLSC grafting/scaffolding on graphene biocompatible 3D preparations emerge as a valuable technique for hPDLSC delivery by improving their proliferation, survival, and differentiation. After demonstrating the beneficial effects of graphene–fibroin substrate composites on hPDLSC proliferation and survival (Figs. 2 and 3), we carried out gene expression assays to further characterize the effect of those on hPDLSC differentiation into bone-like forming cells. The experiments were performed during a 10-day time course to determine the potential effect of graphene–fibroin composites on hPDLSC differentiation in association with the reported proliferation and survival data.
Graphene–fibroin preparations, particularly the reduced configurations, were found to induce the overexpression of early osteoblast/cementoblast markers, including BMP2, RUNX2, ALP, and COL1A1. However, definitive markers of osteoblast (SP7/OSX, BGLAP) or chondroblast (SOX9) differentiation did not show such expression change. Concomitant overexpression of BMP2 and RUNX2 was expected during hPDLSC differentiation [54]. Graphene and fibroin promoted per se the stimulation of BMP2 and RUNX2 overexpression and their combination into bilayer preparations was concordant. Similar results in RUNX2 overexpression were reported by Xie et al. using 2D and 3D graphene scaffolds [41]. Both osteoblast and cementoblast differentiations involve a series of transcriptional events that are dependent on Runx2/Cbfa [55], the downstream effector for BMP-2. Several growth factors belonging to the TGF-β family, including BMPs (such as BMP-2, BMP-3, BMP-4, and BMP-7), have been described to promote proliferation, differentiation, and mineralization of putative cementoblasts [55], also rendering dentin resorption and new cementum formation on dentin surfaces [56]. Runx2/Cbfa is a promoter for Osterix expression at late osteoblast differentiation stages. Importantly, Osterix was found downregulated during terminal differentiation of cementoblasts [54], and thus, our results might suggest a trend to a cementoblastic differentiation instead of therapeutically nondesired osteoblastic differentiation (Fig. 8).
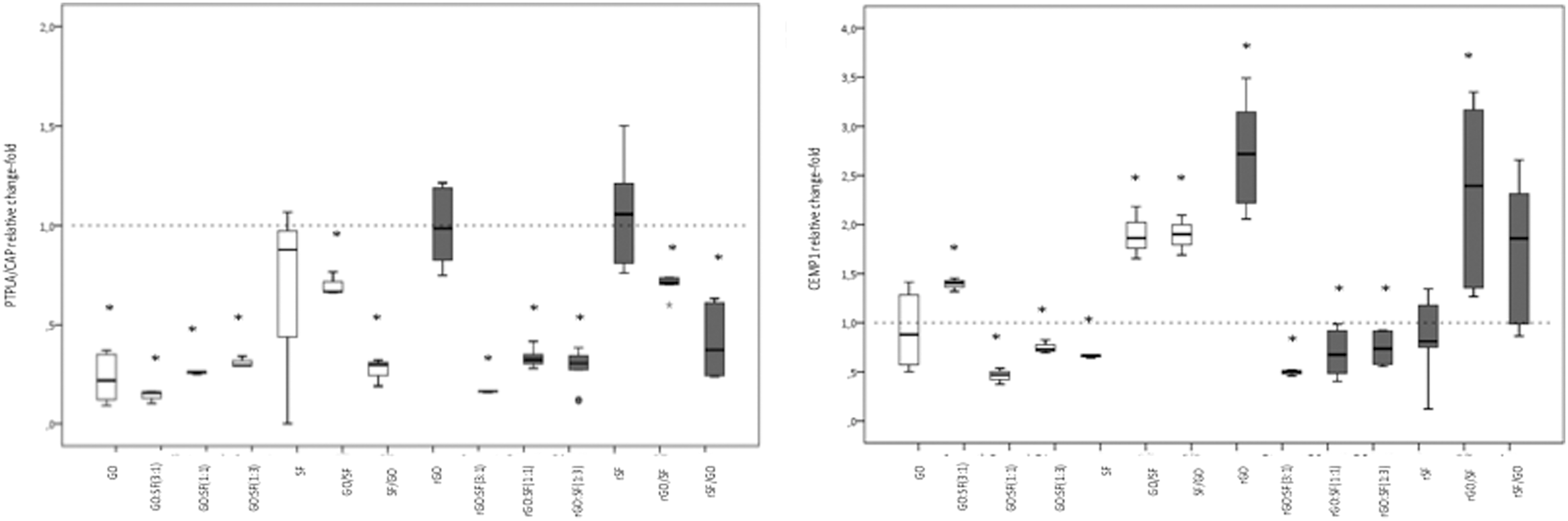
Graphene–fibroin substrates regulate the spontaneous expression of cementoblast-related genes in hPDLSCs after 10 days of culture. Analysis of expression of specific cementum-related genes at early (PTPLA/CAP) or late (CEMP1) differentiation stages to evidence desirable graphene-derived cementoblast differentiation. GO represents oxidized substrates, whereas rGO means reduced substrates. Asterisks represent statistical significance (P < 0.05); ±mean statistical trend. Dotted line indicates control expression of each targeted gene. Each composite was compared to control conditions that were considered to have a 2DDCT of 1. White box plot, oxidized preparations; gray box plot, reduced preparations.
In contrast, proper synthesis and assembly of a collagen-enriched ECM play a crucial role in osteoblast and cementoblast differentiation during the ECM synthesis stage since ECM is a surrogate structure for mineralization. Collagen type I is the most abundant collagen found in bone and teeth [57]. It mainly consists of a heterotrimeric structure of the individual polypeptide α chains specifically containing 2 pro-α1 chains and 1 pro-α2 chain. Therefore, we assessed COL1A1 mRNA expression since it is the main contributor to the procollagen molecule. Almost all the substrate preparations were associated with consistent COL1A1 overexpression although it resulted particularly enhanced when using reduced preparations containing rGO, rSF, or rGO/rSF bilayer. ALP gene expression was also assayed as part of the ECM synthesis and mineralization stage description. Reduced substrates prepared as rGO, rSF, or rGO/rSF bilayer preparations were found to highly induce ALP overexpression. Also, SF was significantly associated with ALP overexpression, contrary to other oxidized substrate preparations assayed (Fig. 7B). ALP is a well-known marker for bone-like differentiation processes since its pyrophosphatase enzymatic activity allows pyrophosphate (a natural inhibitor for tissue mineralization) cleavage into inorganic phosphate [52,55]. The resulting supersaturation of inorganic phosphate released renders hydroxyapatite formation and deposition [52,55].
Regarding the osteocalcin gene (BGLAP) expression, similar to SP7/OSX, we found a significant downregulation in all the substrate preparations, except for rGO and rGO/rSF bilayer preparation, which remained unaffected. Osteocalcin is a small protein related to mineralization and abundantly found in the ECM of bone, dentin, and cementum. However, osteocalcin may not be expressed during hPDLSC maturation and differentiation [57] and some histological studies have unequivocally shown that positive osteocalcin immunostaining may depend on the cementum type and localization [52]. Accordingly, cementoblasts associated to acellular extrinsic fiber cementum are negative for osteocalcin and a weak or absent immunostaining is also found for cellular intrinsic fiber and related cementoblasts [52]. Despite the controversial results, some authors have suggested that they could be the result of phenotypic differences among cementoblasts and cementocytes expressing a more osteoblast-like phenotype [52]. Moreover, recent findings in cell therapy have demonstrated an odonto/cementoblastic differentiation with lack of expression of osteocalcin, demonstrating that this cell differentiation bore little resemblance to osteoblasts [55].
However, the main goal of this article was to demonstrate, for the first time to our knowledge, that hPDLSCs cultured on graphene–fibroin composites can induce cementoblast differentiation, as demonstrated by CEMP1 expression and osteochondroblast-specific gene downregulation. Since substrate preparations were not associated either to late osteoblast differentiation (as demonstrated by OSX or BGLAP expression) or to chondroblast differentiation (since lowered SOX9 expression was obtained), we explored the expression of cementum-related genes, including PTPLA/CAP and CEMP1. Whereas the OsteoDiff medium promoted overexpression of PTPLA/CAP (Fig. 6B), almost all substrate preparations were associated with significant downregulation of PTPLA/CAP, except for rGO and SF (both reduced and oxidized). Nevertheless, rGO and the bilayer preparation, including rGO/rSF or GO/SF highly promoted CEMP1 overexpression, a novel cementum component whose expression is limited to cementoblasts and their progenitors. Noteworthy, proliferation rates in these preparations were moderate and no significant differences (P > 0.05) were found among them (Fig. 2) [58]. Recent findings have identified the existence of functional transcription activator binding regions in the promoter region of PTPLA/CAP and CEMP1, in response to BMP members, including BMP-2 and BMP-7 [54,59]. Importantly, a time-dependent expression profile for PTPLA/CAP and CEMP1 has been recently described in cementoblast-differentiating hPDLSCs. Torii et al. found that BMP-7-dependent cementogenesis was associated with an early overexpression of PTPLA/CAP and downexpression of CEMP1 after 7 days. After 14 days, the authors reported a PTPLA/CAP downregulation accompanied by a CEMP1 overexpression in association with still enhanced RUNX2, ALP, and OSX [54]. Accordingly, our findings regarding PTPLA/CAP and CEMP1 expression may suggest an advanced cementoblast differentiation with a trend to terminal differentiation, given the lowered expression of OSX [54]. Importantly, we found expression of CEMP-1 at the protein level using immunofluorescence experiments (Fig. 9), especially in the substrates rGO and the bilayer preparation, including rGO/rSF or GO/SF. This result confirms the spontaneous cementoblastic differentiation of hPDLSCs observed by qPCR.
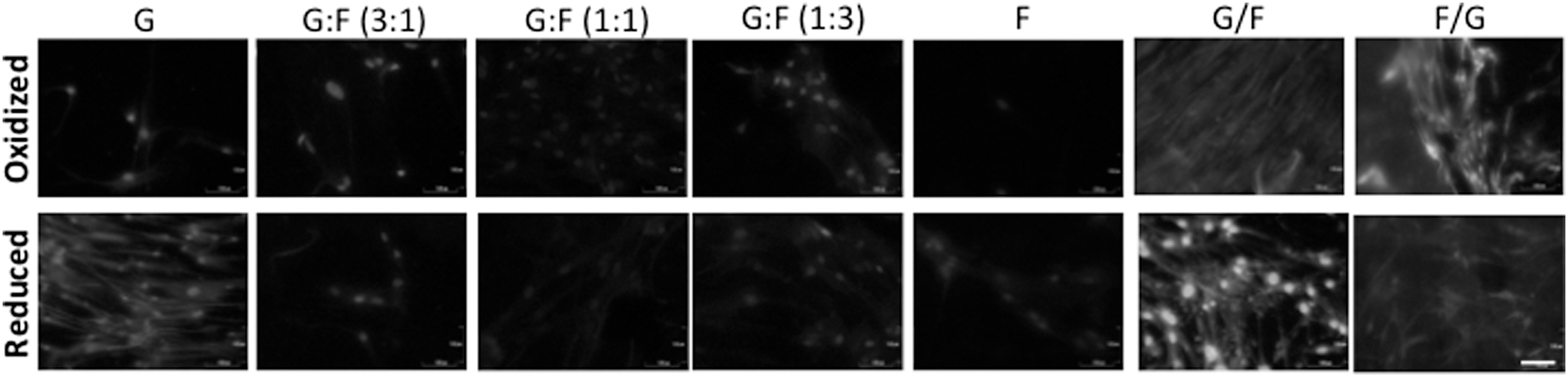
Graphene–fibroin substrates regulate the spontaneous expression of CEMP-1 in hPDLSCs after 10 days of culture. Immunocytochemical analysis of expression of the specific cementum-related protein CEMP1 at late differentiation stages to evidence desirable graphene-derived cementoblast differentiation. Scale bar: 100 μm.
In addition, CEMP1 has been demonstrated to be a cementoblast phenotype marker and a regulator for hPDLSC commitment into cementoblast-like cells being reduced when hPDLSCs differentiate into osteoblasts [60]. Particularly in bone tissue engineering, almost all artificial materials require multiple growth factor administration to promote hMSC differentiation. Conversely to Torii et al., our results provide a new stage for cementoblast differentiation because any biochemical factor is needed and composites alone are enough, which is a very appreciated situation in cell therapy [54].
In this study, we have demonstrated the beneficial effects of combining graphene with fibroin on hPDLSC differentiation into osteo/cementoblast-like cells with moderate proliferation rates. As reported by Kapinas et al., stem cells suppress gene expression conferring lineage commitment, whereas they are self-propagating [58]. Accordingly, an equilibrium between proliferation rates and differentiation stages may be achieved during MSC-like grafting and scaffolding. Noteworthy, here we have demonstrated the best hPDLSC differentiation into osteo/cementoblasts, with gene expression analysis, for those substrate preparations with modest proliferation rates.
Conclusions
Graphene–fibroin-based biomaterials provide a new type of biocompatible scaffolds for stem cells. In graphene–fibroin composites, graphene supplies differentiating capacity, whereas SF causes no inflammatory response after implantation, also providing to GO a better handling and 3D characteristics. Generally, reduced configurations gave better results in proliferation and gene expression, these configurations being the optimal to perform future in vivo experiments. In addition, the ECM resulting from our osteo/cementoblast-like cells may be more physiologically synthesized and mineralized than those from hydroxyapatite-derived substrates. The hPDLSC proliferation rate was consistently improved in certain combinations containing low amounts of graphene and a high fibroin dose. Remarkably, bilayer composites with graphene on the contact surface with cells promoted a moderate proliferation, and favored a osteo/cementoblast differentiation of hPDLSCs in the absence of any growth factors.
Footnotes
Acknowledgments
This work was supported by the Spanish Net of Cell Therapy (TerCel) provided by Carlos III Institute of Health (ISCiii) (RETICS RD07/0010/2012 and RD12/0019/0001) together with the Junction Program for Biomedical Research in Advanced Therapies and Regenerative Medicine from ISCiii and FFIS. Experiments made at the IMIDA were supported by funds from the Spanish Government (MCI) projects MAT2008-06702 and FEDER Operative Program of the Region of Murcia 2007–2013 from the UE.
Author Disclosure Statement
No competing financial interests exist.