
Abstract
Fibroblasts can be transdifferentiated directly into other somatic cells such as cardiomyocytes, hematopoietic cells, and neurons. An advantage of somatic cell differentiation without first generating induced pluripotent stem cells (iPSCs) is that it avoids contamination of the differentiated cells with residual iPSCs, which may cause teratoma. However, since primary fibroblasts from biopsy undergo senescence during repeated culture, it may be difficult to grow transdifferentiated cells in sufficient numbers for future therapeutic purposes. To circumvent this problem, we reversibly immortalized primary fibroblasts by using the piggyBac transposon to deliver the human telomerase reverse transcriptase (hTERT) gene hTERT plus SV40 Large T. Both approaches enabled fibroblasts to grow continuously without senescence, and neither caused teratoma formation in immunodeficient mice. However, fibroblasts immortalized with hTERT plus SV40 large T antigen accumulated chromosomal rearrangements, whereas fibroblasts immortalized with hTERT retained the normal karyotype. To transdifferentiate hTERT-immortalized fibroblasts into other somatic lineage cells, we transiently transfected them with episomal OCT4 and cultured them under neural cell growth condition with transposase to remove the transposon. Tripotent neural progenitor cells were seamlessly and efficiently generated. Thus, reversible immortalization of primary fibroblasts with hTERT will allow potential autologous cell-based therapeutics that bypass and simulate iPSC generation.
Introduction
S
An alternative is to transiently immortalize somatic cells before transdifferentiation so that enough cells can be expanded for therapy. With this approach, immortalization genes such as those encoding human telomerase reverse transcriptase (hTERT), SV40 large T antigen (SV40LT), and Bmi-1 are expressed alone or in combination in primary cells and then removed typically with the Cre-loxP system [6 –9]. However, a 34-bp fragment is left, which may disrupt the functions of other genes [6,8,10]. Using a piggyBac (PB) transposon to integrate and excise selectable marker genes leaves no residual sequence behind [11].
In this study, we used the PB transposon to reversibly immortalize primary fibroblasts and expand them in quantities sufficient for possible cell therapy. Fibroblasts transformed by hTERT or hTERT plus SV40LT proliferated for the duration of our studies at 140 days. Cells transformed with hTERT+SV40LT had chromosomal abnormalities and proliferated twice as fast as those transformed with hTERT, which retained the normal karyotype. Removal of hTERT by transfection with transposase abolished their proliferative potential. When fibroblasts immortalized with hTERT were transfected with episomal Oct4 and PB transposase, the transposon insert was seamlessly removed, and the cells could be directly converted into neural cells.
Materials and Methods
The Institutional Review Board of the California University, San Francisco (UCSF) School of Medicine and the UCSF Stem Cell Research Oversight Committee approved this study. The animal protocols were approved by the UCSF Animal Research Committee. A total of 20 NOD/SCID mice, ages 8 weeks were used for this study.
Plasmid construction
All restriction enzymes and Phusion high-fidelity DNA polymerase were from New England Biolabs. The plasmids 5′-PTK-3′ and hyperactive piggyBac transposase (PBase) were kind gifts from Allan Bradley (Wellcome Trust Sanger Institute, Hinxton, United Kingdom). Fragments containing the CMV early enhancer/chicken β actin (CAG) promoter were released from pCAGGS (kindly provided by Dr. Jun-ichi Miyazaki, Osaka University, Japan) by SalI and EcoRI digestion and ligated into SalI and EcoRI sites in pBluescript-SK (termed pSKCAG). A 2.5-kb fragment of hTERT was released from pBabe-hygro-hTERT with EcoRI and BamHI and ligated into pSKCAG (termed pSKCAGhT1).
The coding fragments of hTERT (hT2), enhanced green fluorescent protein (EGFP), and SV40LT were amplified from pBabe-hygro-hTERT, pEGFP, pEGFP-N3 (Invitrogen), respectively, and pSG5-LT with primers containing 2A self-cleaving peptide sequences (Supplementary Table S2) and subcloned into pGEM-T for sequencing. CAG fragments were released from the SalI and BamH I sites of pSKCAGhT and ligated into the XhoI and MfeI sites of 5′-PTK-3′ with hT2, which was released from pGEM-T-hT2 by BamH1 and EcoRI digestion. This vector was designated pT1. To generate the pT2 vector, the coding fragments of hT2, SV40LT, and EGFP were released from their individual T vectors and ligated together into the AscI and NotI sites of 5′-PTK-3′ with CAGhT1. pBabe-hygro-hTERT and pSG5-LT plasmids were from Addgene.
Cell culture
The fibroblasts we used in this study were from one donor given to us anonymously. and maintained in the fibroblast medium consisting of Dulbecco's modified Eagle's medium (DMEM) (Gibco), 10% fetal bovine serum (Hyclone), 1 mM
Generating tripotent neural progenitor cells from immortalized fibroblasts
hTERT fibroblasts (4 × 105) were transfected with 4 μg of episomal human Oct4 and 2 μg of PBx (Transposagen Bio) using Lonza primary fibroblasts transfection kits. Transfected cells were cultured in a reprogramming medium consisting of DMEM/F12, 10% knockout serum replacement (Gibco), 1 mM
Statistical analysis
All experiments were performed at least twice, and all samples were tested in triplicate. Data from each group are expressed as mean ± standard error. The Student's t-test was used for statistical analysis.
Results and Discussion
Conditional immortalization of fibroblasts with PB transposon
Traditionally, cells have been immortalized by expressing hTERT, Bmi, SV40LT, or oncogenes such as HPVE6/E7 or c-MYC. Ectopic expression of hTERT has been used to immortalize somatic cells without altering the normal karyotype [9]. The ability of SV40LT to immortalize somatic cells is largely dependent on its ability to complex with p53 [14]. Thus, SV40LT has become a popular gene to immortalize primary cells; the immortalized cells are generally not tumorigenic [14,17].
We inserted hTERT (driven by CAG promoter) and the puromycin-TK hybrid gene (Puro-TK), joined by an internal ribosome entry site, between the inverted terminal repeats of the construct pT1. In pT2, we inserted both these genes and SV40LT and EGFP; all four genes were joined by sequences encoding 2A peptides (Fig. 1A). The TTAA sequences on both ends of the inverted terminal repeats allow PB-mediated integration and seamless excision of the constructs from the integration sites [11]. Each construct was transfected in the presence of PBase into foreskin fibroblasts. To monitor transfection efficiency, 293T cells were transfected with pT2. Expression of EGFP was strong in 293T cells and weak in primary fibroblasts (Supplementary Fig. S1; Supplementary Data are available online at
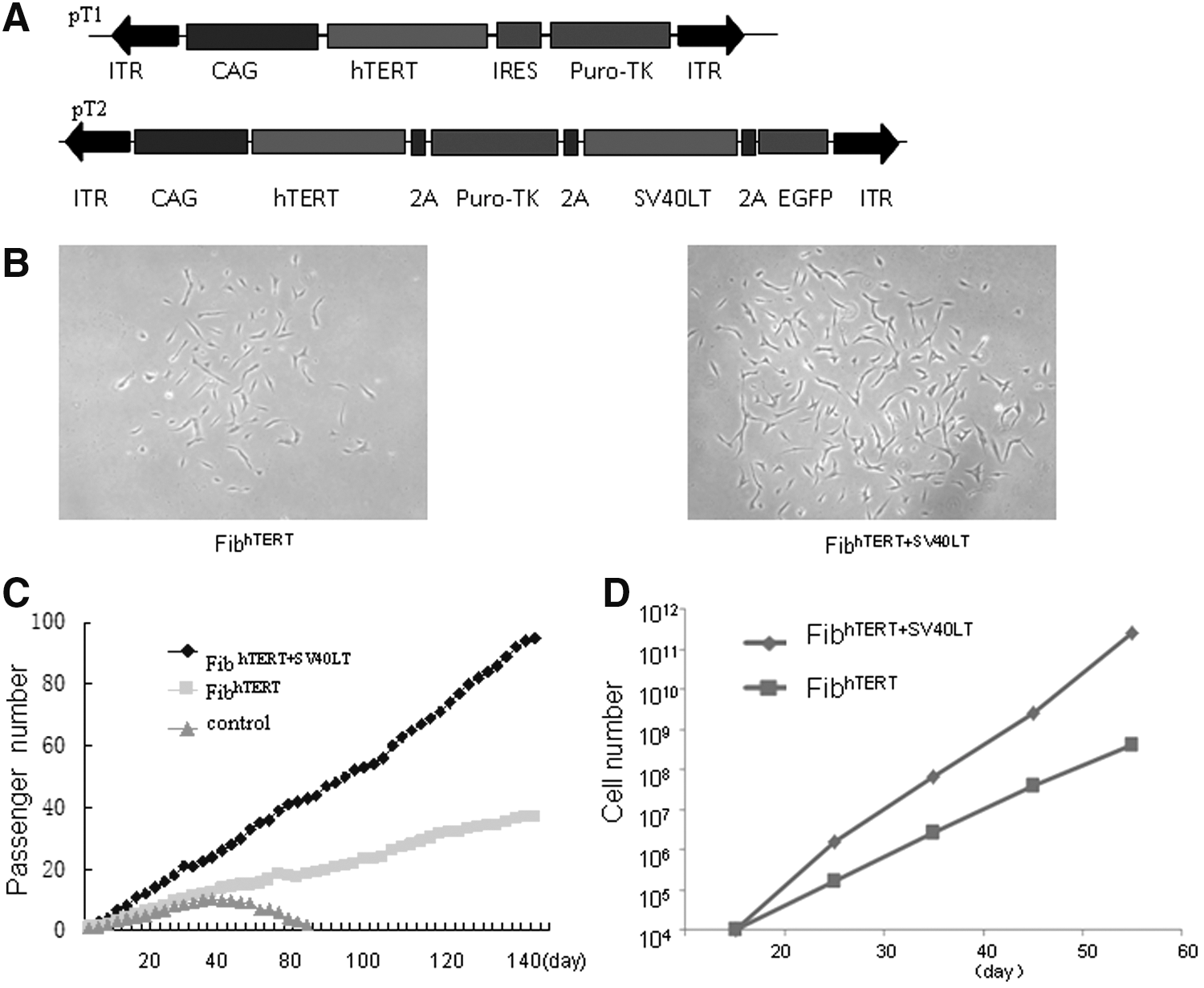
Generation of immortalized fibroblasts by PB-mediated gene transfer.
Karyotype, soft agar colony assay, and in vivo tumorigenicity
As shown by karyotype analysis at passage 55, fibroblasts immortalized with hTERT had normal diploid chromosomes, but fibroblasts immortalized with hTERT+SV40LT had abnormal karyotypes, including complex chromosome losses and multiple (>20) overlapped clonal abnormalities (Supplementary Fig. S4A). These chromosome defects were not surprising, as large T antigen often causes genomic instability and malignant transformation. Neither type of immortalized fibroblasts formed colonies in soft agar during 3 weeks of culture. In contrast, 293T cells did form colonies (Supplementary Fig. S4B). Three months after subcutaneous injection into NOD/SCID mice, neither hTERT nor hTERT/SV40LT-transduced cell lines gave rise to tumors, even though the latter had grossly abnormal karyotypes. As control, tumors were found in mice injected with 293T cells (Supplementary Table S1).
These findings show that fibroblasts can be immortalized by both constructs. However, the cell phenotypes were distinctly different. Cells transduced with SV40LT+hTERT proliferated rapidly, and led to round cells, aggregation, and absence of contact inhibition and anchorage-independency in the soft agar assay. In contrast, hTERT-transduced cells proliferated much more slowly and exhibited contact inhibition and anchorage-dependency. For further studies, we used only fibroblasthTERT with normal karyotypes.
Determination of integration sites mediated by PB transposon
PB transposons can integrate into multiple chromosomal sites. This property has been used for gene discovery, as insertion in or near some genes disrupts their functions [18]. However, for seamless alteration of genes, cells with a single integration site are preferable because it would be difficult to excise all the insertions in cells with multiple integration sites. We used Southern blot to determine the number of PB integrations in the cell lines (Fig. 2A) and inverse PCR to determine the chromosomal location of the integration in cell lines with a single integration site (Fig. 2B). The PCR products were separated on agarose gels, the fragments isolated for sequencing (Fig. 2C), and BLAST searches used to align the insertion site to the NCBI human genomic DNA database. Analysis of the PB-mediated integration site bearing “TTAA” indicated that there were five integration sites, 2 on chromosome 11 and one each on chromosomes 8, 9, and 22 (Fig. 2D); four were in intergenic locations and one was in an intron. The 2 integration sites on chromosome 11 were at the β-globin cluster, one in the intron of the β-globin gene, and the other in the region between the δ- and the β-globin genes. Perhaps the PB we used preferentially recognizes and integrates in this area. For deimmortalization, we selected the immortalized fibroblasts with an integration site at chromosome 9, as that site is farthest from neighboring genes.
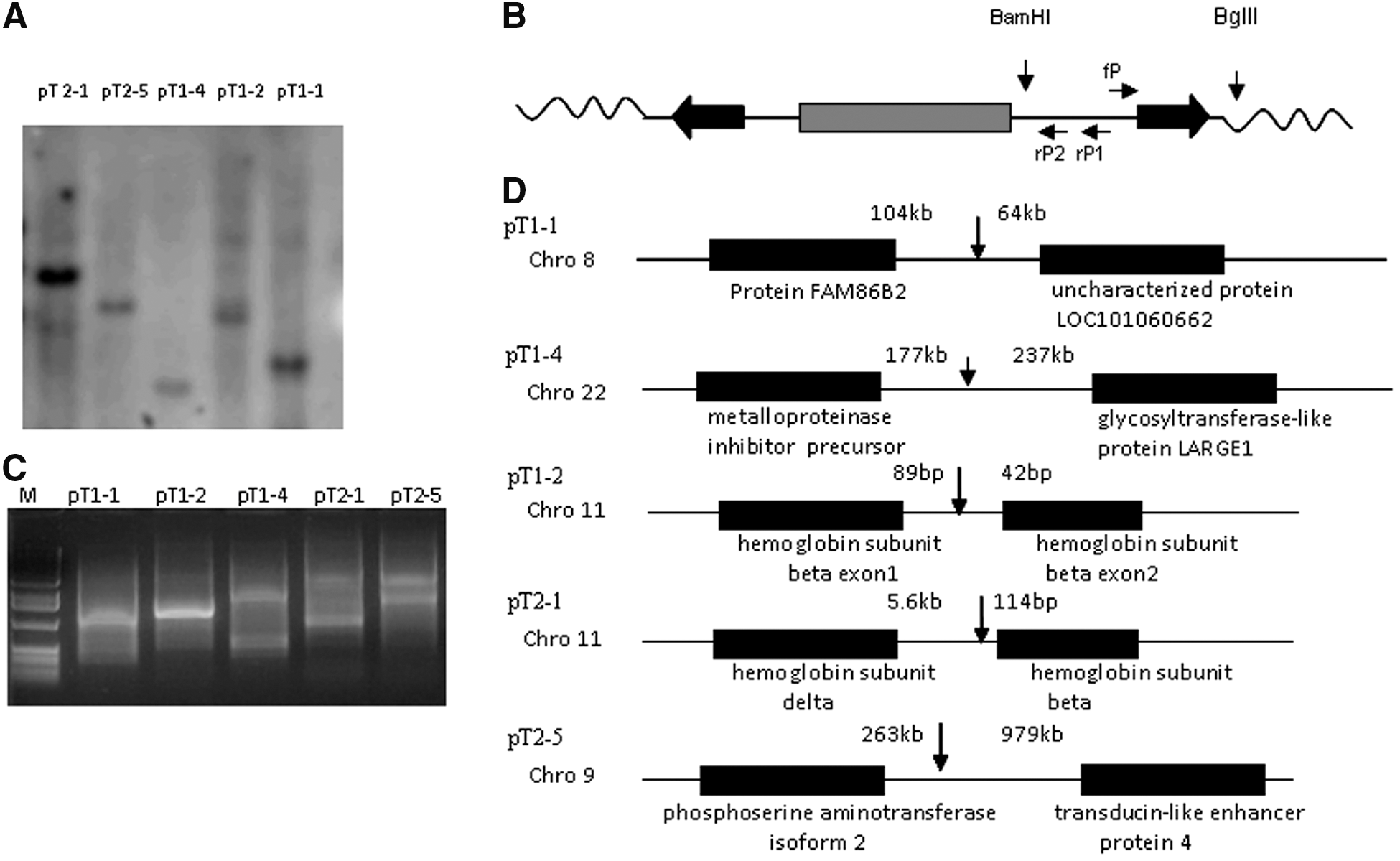
Determination of integration sites in immortalized cell lines by inverse PCR.
Reversal of immortalization of fibroblasts after PBase transfection
We sought to evaluate whether the immortalization-related phenotype of the fibroblasts could be reversed after removing the transgene integrated at the TTAA sites between two inverse terminals repeats. One hTERT-transduced cell line was expanded after passage 35, transfected with PBase, and negatively selected with 0.5 μM FIAU for 7 days. Analysis of half of the surviving cells by PCR with primers designed to amplify the CAG in the cassette or at the vector–genome junction confirmed that the selection cassette had been removed (Fig. 3A–D). Sequencing of the PCR product with primers flanking the integration site showed seamless replacement of the vector–genome junction with the normal genomic sequence (Fig. 3B). The remainders of the surviving cells were analyzed for proliferation rate. PBase-transduced FIAU-resistant fibroblasts with the PB insert removed had significantly decreased proliferation and quickly became senescent showing positive SA-β gal expression in days (Fig. 3E–G). Thus, the immortalized fibroblasts had a long-term high proliferative activity that was reversible.
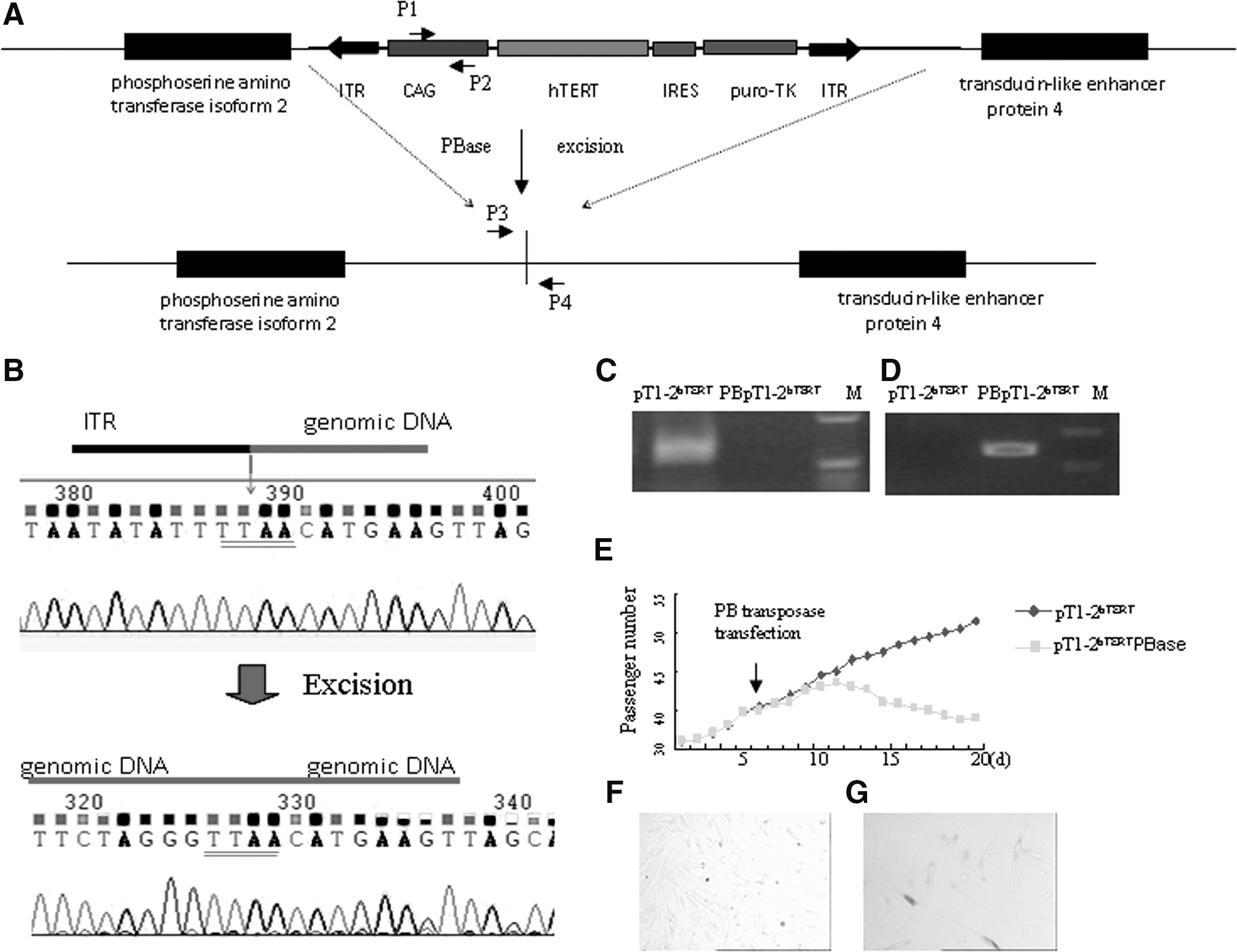
Reversal of cell immortalization after excision of PB with transposase.
Direct conversion of immortalized fibroblasts to PB-free NPCs
Fibroblasts have been converted into NPCs with a single transcription factor, human Oct4, delivered by lentivirus [12]. To determine whether hTERT-immortalized fibroblasts can be transdifferentiated into NPCs, we transfected hTERT fibroblasts (passage 20) and control fibroblasts (passage 10) with an episomal plasmid encoding human Oct4 and a plasmid encoding PBase, cultured the cells under neural transdifferentiation conditions for 10 days (Fig. 4A), and transferred them to the NPC medium. After 1 week, neural sphere-like cells were detected (Fig. 4B). The immortalized cells generated 10-fold more spheres than the controls (Fig. 4C). After the neural spheres were dissociated and cultured on the Matrigel for 1 week, the cells stained positively for the NPC markers N-cadherin, PAX6, and nestin (Fig. 4D, F, G). PCR analysis of cells cultured with FIAU (negative selection) confirmed the removal of the hTERT cassette (Fig. 4E).
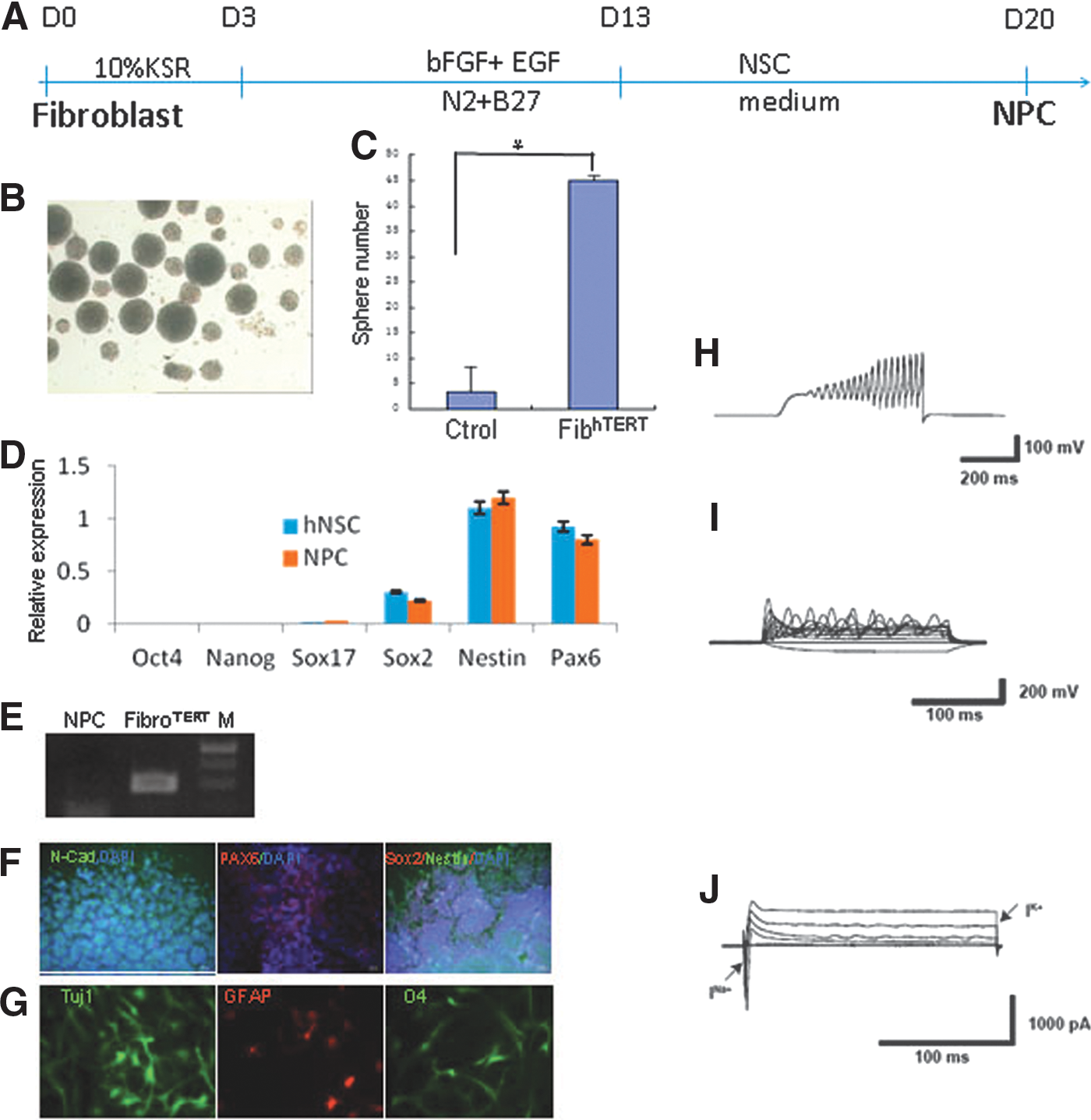
Direct generation of NPC from immortalized fibroblast.
To further differentiate the NPCs, we removed the growth factors and used culture protocols reported to differentiate NPC into different lineages. After 3 weeks of differentiation, neurons, astrocytes, and oligodendrocytes were detected (Fig. 4G). To characterize the neurons, patch clamp recording was used to examine their electrophysiological properties. Totally 13 neurons were recorded and 9 of them (69.2%) showed action potential firing, fast sodium currents, and outward potassium currents (Fig. 4H–J). Those NPCs cultured for only 1 week were used as controls. For control cells, none of them (n = 6) showed action potential firing or sodium currents.
In summary, we reversibly immortalized primary fibroblasts by using the PB transposon to deliver hTERT and as an example, directly transdifferentiated them into neural cells. This strategy will make it possible to generate large numbers of primary cells from small clinical biopsies for gene editing or making other modifications for therapeutic purposes. The cells can then be directly transdifferentiated into other somatic cells, thereby bypassing the potentially tumorigenic pluripotent state.
Footnotes
Acknowledgments
We thank Changsheng Lin at the Gladstone Institutes for kindly providing cDNAs from hNSC. This work was supported by NIH grant P01-DK088760.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.