
Abstract
A healthy and transparent cornea is essential for exquisite vision. During adulthood, its epithelium is constantly replenished through the activity of its stem cells (SCs). Precisely when these cells develop and their distribution across the ocular surface remain incompletely characterized in man. We postulated that the human fetal cornea harbors SCs that can be identified with keratin (K) 14 and αv-integrin, two markers we and others previously used to identify their adult counterparts. Immunofluorescence, cell culture, quantitative real-time polymerase chain reaction (qRT-PCR), and colony-forming assays were performed on fetal and adult biomaterial to locate progenitors and establish their phenotypic and functional properties. K14 was used to map the spatiotemporal distribution of precursor cell activity across the developing cornea, divulging a dynamic pattern of vertical and horizontal consolidated expression with increasing gestational age. K14 was coexpressed with αv-integrin in fetal and adult corneas and cultured corneolimbal epithelium, and colony-forming efficiency (an indicator of SC activity) was similar in cells from both sources. Finally, fetal cells were adherent, grew well, and maintained a K14 phenotype on contact lenses, a substrate we previously used to deliver cells to patients with blinding corneal disease. This study provides valuable insights into the development of the cornea, including the formation of the SC repository, the distribution of these cells across the ocular surface, and a preliminary attempt at harnessing, phenotyping, and functionally characterizing these cells. Future studies will focus on isolating fetal SCs to determine their utility as an alternative cell therapy for patients suffering from corneal blindness.
Introduction
T
The anatomical development of cornea has been extensively studied in animals, including chicks [12], rabbits [13], rodents [14], and fish [15]. Pivotal studies in humans were conducted by Duke-Elder and Cook [16], Mann [17], and Sevel and Isaacs [18]. The formation of the human cornea is triggered by the interaction between the lens vesicle and the overlying surface ectoderm, which occurs during the final stages of eye development [19]. The corneoscleral junction, which eventually forms the limbus, is first apparent at the end of the embryonic period [8 weeks' gestation (WG)] and is marked by morphological changes in stromal cells in the peripheral cornea [16]. The palisades of Vogt that characterize the adult limbal SC niche are not visible in fetal specimens. Instead, a dome-like structure with a thicker epithelium is apparent, which is hypothesized to represent the primitive SC lair. Work in our laboratory and others suggest that maturation of the limbal niche in humans may extend well into the postnatal period [20, 21]. Further investigations are needed to gain comprehensive insights into the morphogenetic programs and biological processes that dictate SC activity on the developing human ocular surface, with a view to harnessing these cells for potential clinical use.
To date, studies in the human fetal cornea to detect SCs have been scarce. Rodrigues et al. [22] localized K3 suprabasally throughout the entire fetal corneal and limbal epithelium. This finding led the authors to postulate that SCs are deposited uniformly in the basal epithelial layer of both the cornea and limbus until the postnatal period. A study conducted in our laboratory found that K15 was expressed across the ocular surface during the early gestational period (8–14 WG); its expression spatially segregated to the limbal epithelium from 17 WG onward, and by 22 WG, staining was confined to the basal limbal epithelium alone [20]. Notably, other studies performed on the developing rat and mouse corneas [14, 23, 24] also support the notion that SCs are initially distributed across the entire ocular surface but consolidate to the limbal zone with increasing gestational age.
The current study was initiated to (i) document the dynamic changes that occur within the primordial epithelium during corneal development in humans, (ii) determine whether K14, a marker used to illuminate corneolimbal epithelia in the developing mouse [25 –27] corneas, could serve to detect SC activity on the human fetal ocular surface during a narrow but critical developmental window, that is, before eyelid contraction, and (iii) establish whether the phenotypic and functional features of fetal cells are comparable with those of adult equivalents, with a view to developing an alternative cell therapy for patients suffering from blinding corneal disease.
Materials and Methods
Human tissue specimens
Fresh–frozen and paraffin-embedded (n = 41) human fetal and adult eyes were used for immunohistological assessments. For culture purposes, fresh fetal eyes (n = 10) were obtained from legal terminations, and fresh normal donor corneas (n = 10) were acquired from the Lions NSW Eye Bank (Sydney, Australia). Institutional Human Research Ethics Committee clearance was obtained to use these samples. Research using these specimens was conducted according to the principles of the Declaration of Helsinki.
Histology and immunofluorescence
Fresh–frozen 95% ethanol or 4% paraformaldehyde-fixed and 10% formalin-fixed, paraffin-embedded fetal and adult corneas, as well as whole eyes, were examined. Some tissues were cut and processed for H&E staining. Others were assessed by immunofluorescence. In brief, tissue was heated by microwaving in 0.01 M citrate buffer (pH 6.0) and then incubated in blocking solution [20% goat serum in 2% bovine serum albumin (Sigma, Sydney, Australia) containing Tris-buffered saline (0.05 M TBS, pH 7.6)] for 30 min. Predetermined dilutions of mouse monoclonal or rabbit polyclonal primary antibodies (Supplementary Table S1; Supplementary Data are available online at
Cell culture
Fetal eyes were obtained with <4-h postmortem delay. Fresh whole adult donor corneas were stored in Eye Bank Organ Culture Media at 34°C for several days before use. Fetal corneal and adult limbal tissue explants (∼3 mm2) were placed epithelium side down in six-well plates (CELLSTAR™; Greiner Bio-One, Frickenhausen, Germany) in the presence of keratinocyte serum free medium (KSFM) (Gibco®; Fremont, CA) or CnT-PR (CELLnTEC, Bern, Switzerland) media and maintained at 37°C in a humidified 5% CO2 incubator. Explants were removed after 14 days to minimize the risk of fibroblast contamination, and cells were subcultivated by enzymatic dissociation with TrypLE™ (Invitrogen). Some cells (∼1 × 104) were dispensed into vitronectin-coated eight-well chamber slides (BD Falcon, Franklin Lakes, NJ). After expansion, they were fixed in cold 100% methanol and then stained for respective antigens. In other experiments, epithelial cells were seeded on contact lenses (CIBA Vision®; Duluth, GA) as previously described [28]. One week later, lenses were fixed in 4% paraformaldehyde and stained for K14 (as above). Immunoreactive cells on glass slides or contact lenses were counted in 10 randomly selected fields by two researchers.
For colony-forming assays, epithelial cells were seeded on six-well plates at a density of 5 × 104 cells/well, and their ability to form colonies was monitored. After 2 weeks, cells were fixed and stained with 1% rhodamine blue (Sigma). Densely populated colonies with smooth edges and a diameter of ≥3 mm were independently counted by two researchers. Colony-forming efficiency (CFE) was determined by dividing the number of colonies by the total number of cells seeded.
Quantitative real-time polymerase chain reaction
RNA was extracted using the RNAgents™ Total RNA Extraction Kit (Promega, Madison, WI) and reverse transcribed into cDNA using the SuperScript™ III RT system (Invitrogen) with oligo dT primers. Aliquots of cDNA (1 μL) were amplified by polymerase chain reaction (PCR). Each PCR mixture contained 1 μL of cDNA template, 2 μL of primer mix (Qiagen, Hilden, Germany) (Supplementary Table S2), 10 μL of LightCycler® 480 SYBR Green I Master (Roche Diagnostics, Indianapolis, IN), and RNase-free water in a total volume of 20 μL. All other conditions have been previously described [29].
Statistical analysis
Statistical analysis was performed using Prism V6 (GraphPad Software, Inc., La Jolla, CA). One-way analysis of variance was used to analyze data from immunocytochemistry, whereas all other data were analyzed using unpaired two-tailed t-tests. P < 0.05 was considered significant.
Results
Histology of the human fetal cornea
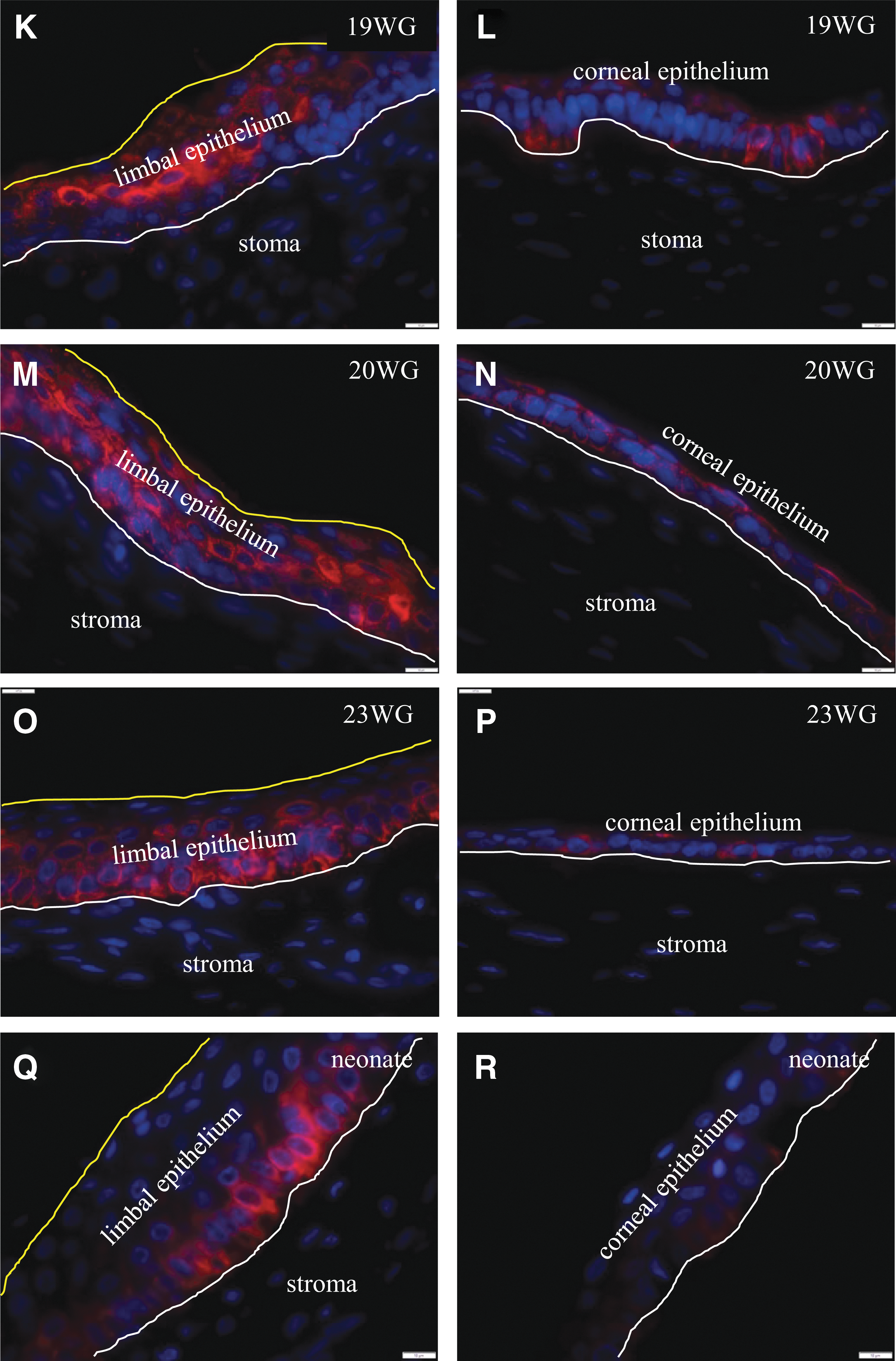
Histological features of human fetal corneas were determined after staining representative sections with H&E (Fig. 1). Specimens at 8–10 WG displayed a bilayered epithelium and a densely packed stroma, common to the limbus and central cornea (Fig. 1A, B). By 12 WG, the limbal epithelium, a region previously dubbed the limbal dome [20], had increased to three layers (Fig. 1C, solid line), while the central corneal epithelium remained bilayered with a basal cuboidal and a superficial squamous cell layer (Fig. 1D). At this stage of development, mesenchymal cells within the stroma were sparser than in samples of earlier gestational age. From 17 to 23 WG, the epithelium within the limbal dome steadily increased to five layers (Fig. 1E, G, and I, solid line). In the neonate, the limbal epithelium was at least six layers thick and consisted of basal cuboidal intermediate wing and superficial squamous cells (Fig. 1P) similar to the adult cornea (not shown). The central corneal epithelium remained bilayered from 8 to at least 23 WG (Fig. 1B, D, E, H, and J), and by this time point, Bowman's layer was readily discernable (Fig. 1J, arrows). A multilayered epithelium was noted in the neonatal central cornea (Fig. 1L). The palisades of Vogt were not detected in any fetal or neonatal cornea (Fig. 1A–L).

Histological features of the human cornea. Histological features of the human fetal
Keratin 14 expression in the fetal cornea
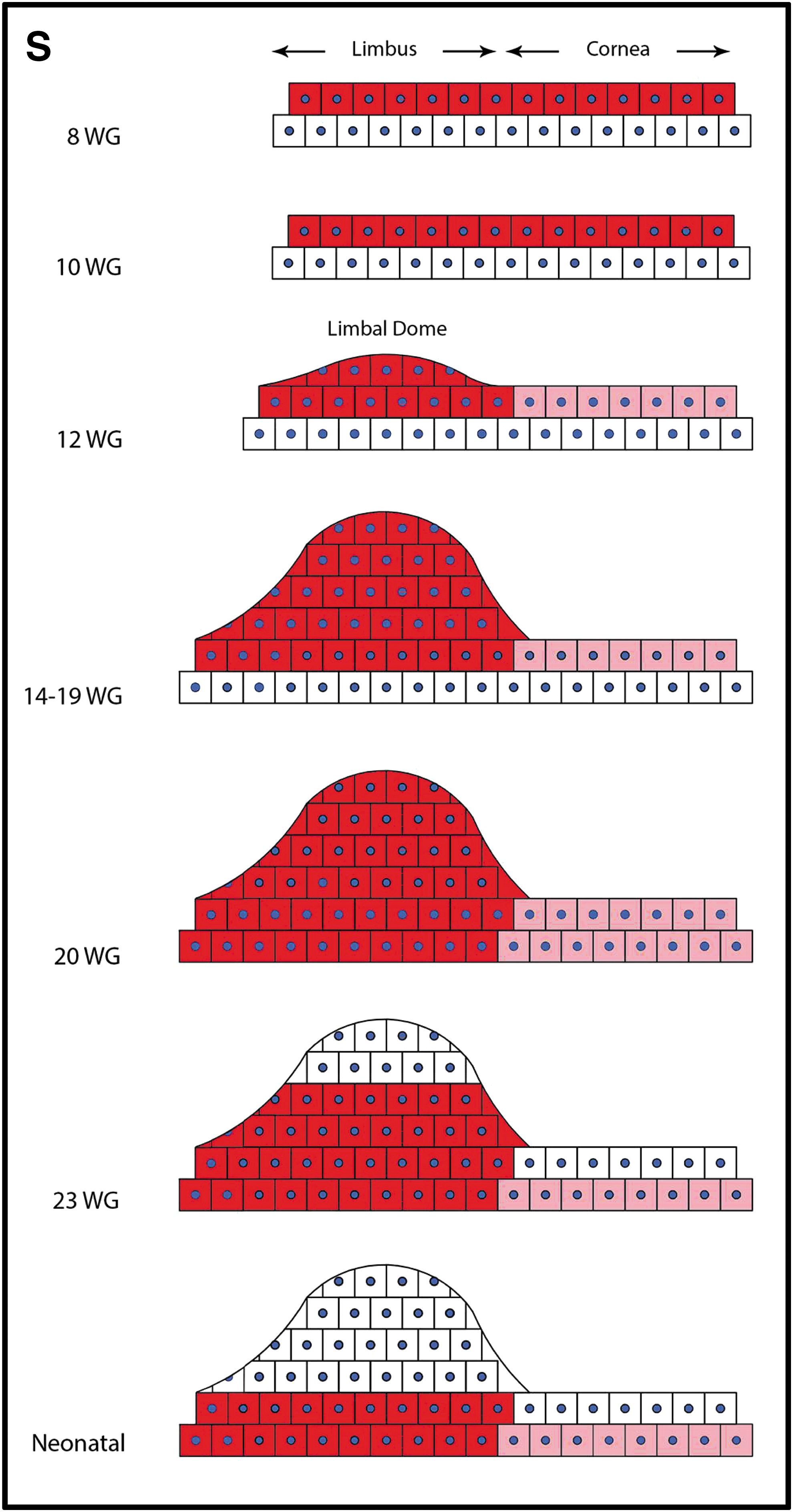
Next, the spatiotemporal expression of K14 was evaluated in developing human corneas. In specimens of 8–10 WG, K14 was consistently expressed in the superficial epithelia across the entire cornea, both peripherally and centrally (Fig. 2A–D). Notably, this is the time frame in which eyelids envelope and fuse over the ocular surface (Fig. 2A–C), and because K14 also marks cutaneous epithelial progenitors [30], its expression was intense in these evolving structures (Fig. 2A, B), especially in the eyelid fusion zone (Fig. 2C) [26]. With increasing gestational age (ie, between 12 and 19 WG), and with the appearance of the limbal dome at about 12 WG (Fig. 2E, yellow line), K14 was the strongest in suprabasal and superficial cells within this structure but was absent from the basal layer (Fig. 2E, G, I, and K). At 20 WG, K14 was present in all layers of the limbal epithelium (Fig. 2M), then regressed to the suprabasal and basal cells at 23 WG (Fig. 2O), and was restricted to basal epithelia in the neonatal cornea (Fig. 2Q) similar to its distribution in adult specimens [31]. While K14 expression was initially widespread across the corneal surface during early gestational periods (ie, 8–10 WG), its expression faded from this region with increasing time postconception (ie, from 12 to 23 WG) (Fig. 2F, H, J, L, N, and P). Finally, in the neonate, staining for K14 was negligible in the central corneal epithelium (Fig. 2R) corresponding to our observations in adult corneas [31]. The schematic diagram summarizes the spatiotemporal distribution of K14 during corneal development (Fig. 2S).

Spatiotemporal expression of K14 in developing human cornea. Immunofluorescence was performed on formalin-fixed corneas from 8.0
Coexpression of K14 with other putative LESC markers
Following from these results, we wanted to identify other markers that could discern fetal corneal epithelial precursors. Among the antigens we searched for, Hes1 (Fig. 3B, E), Notch1 (Fig. 3H, L), and αv-integrin (Fig. 3P, S) were consistently coexpressed with K14 in both the fetal and adult limbal epithelia (Fig. 3C, F, I, M, Q, and T). In contrast, negligible expression of Hes1, Notch1, and αv-integrin was found in the fetal and adult central corneal epithelium, matching the staining pattern for K14 within this region (data not shown). Importantly, other SC-related markers were included in our analyses (eg, N-cadherin, ABCG2, and Bmi1); however, inconsistent staining for these antigens arose between (and within) fetal and adult specimens (data not shown). Therefore, a decision was made to focus on αv-integrin since we previously proposed it as a candidate putative LESC marker [32], and its location on the cell surface is an attractive feature that can facilitate the isolation of these cells.
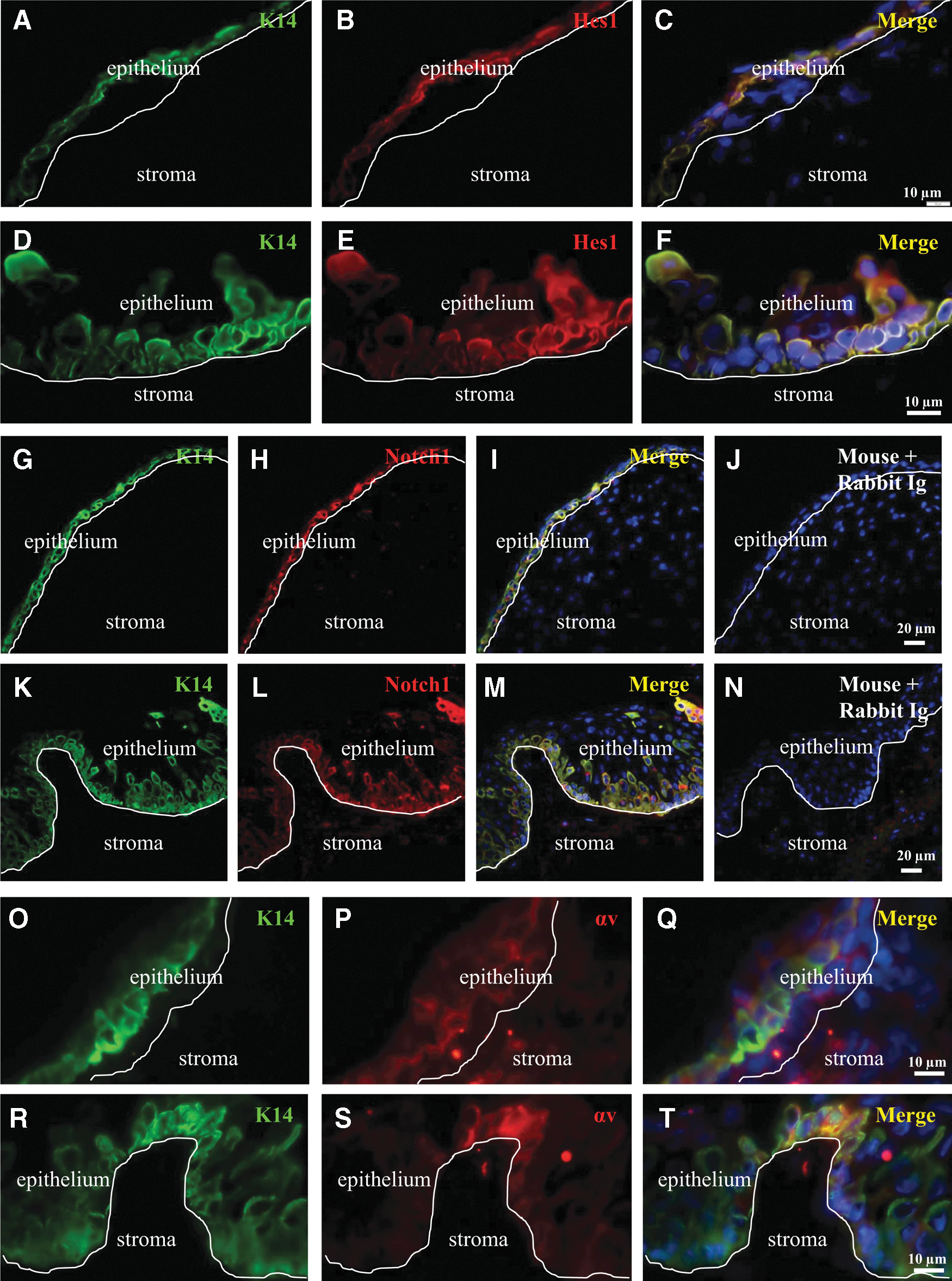
Coexpression of K14 with other putative LESC markers in fetal and adult corneas. Double immunofluorescence was conducted on sections derived from a 17 WG fetal
In vitro expansion of corneolimbal epithelia
To this end, different culture media were assessed to determine the growth properties of fetal explant-derived epithelia. First, epithelial cells emerged from fetal explants within 2–3 days (Fig. 4A, C). Second, cells from early generations (P1) were generally smaller, while those from later passages (>P6) were larger and vacuolated, implying progression to differentiation and senescence (data not shown). Third, cells nurtured in CnT-PR (Fig. 4A, B) were morphologically and functionally (mitotically) similar to those cultivated in KSFM (Fig. 4C, D) and resembled those that expanded from adult limbal explants (data not shown). Therefore, for consistency, all subsequent characterization studies were conducted on cells nurtured in KSFM.
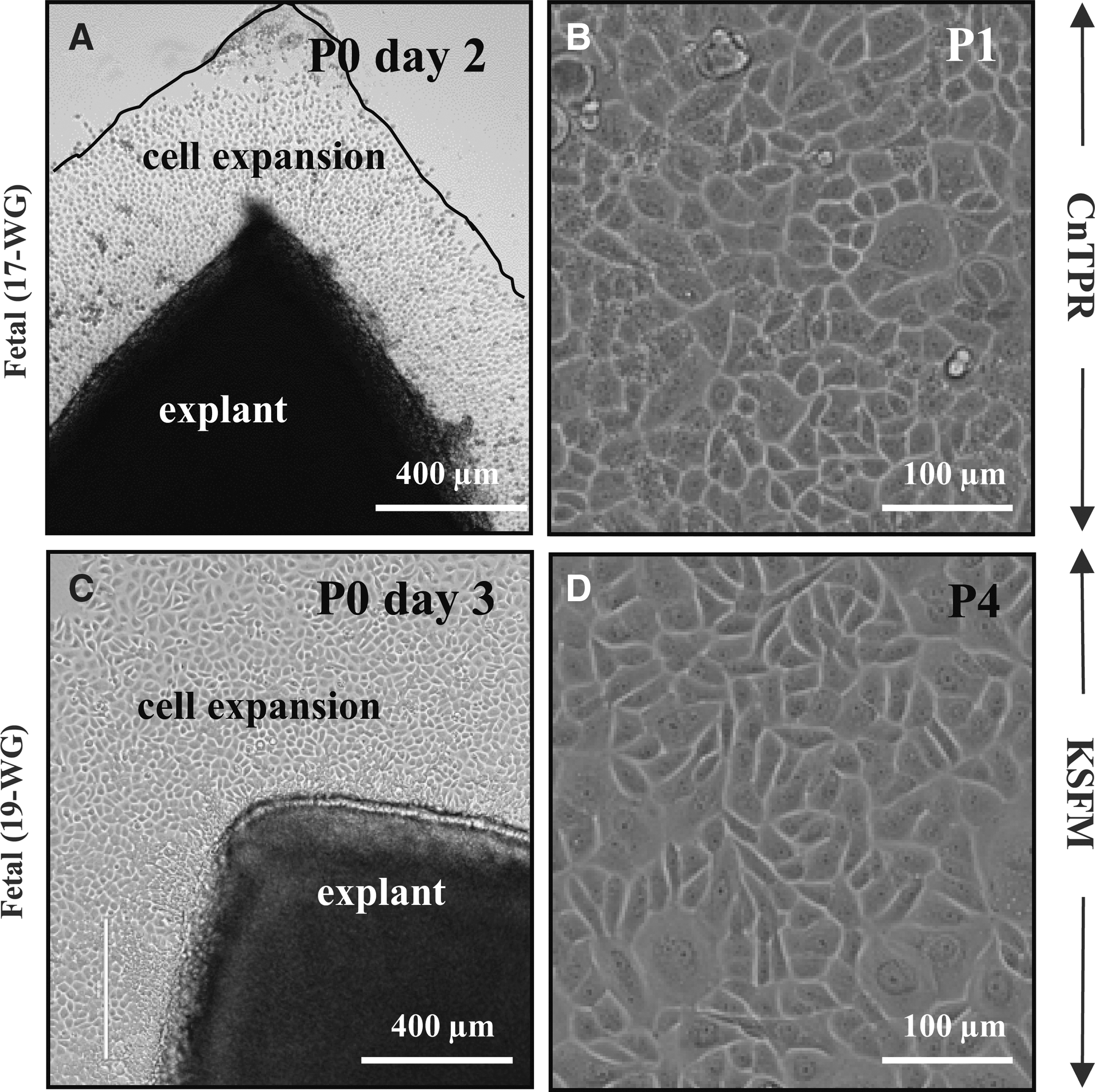
Explant-derived human fetal corneolimbal epithelial cells. Fetal ocular surface epithelial cells were established from corneal explants. Tissue was placed in either CnT-PR
Characterizing corneolimbal epithelial cells
Epithelial cells were seeded into vitronectin-coated chamber slides and stained for K14 and αv-integrin. A similar immunostaining pattern evolved for fetal and adult epithelia, where some cells that expressed K14 coexpressed αv-integrin (Fig. 5A–F). K14+, αv-integrin+, and K14/αv-double-positive fetal epithelial cells represented 13.4% ± 0.8%, 10.8% ± 0.3%, and 3.7% ± 0.4%, respectively, of the total population (Fig. 5G). This was comparable with adult epithelial cells: 9.6% ± 0.5%, 7.7% ± 0.6%, and 3.5% ± 0.1%, respectively (Fig. 5H). Moreover, quantitative real-time PCR (qRT-PCR) disclosed a similar gene expression profile for K14 and αv between fetal and adult epithelia (Fig. 5I, 5J, respectively), and CFE was not significantly different (P = 0.35) between fetal and adult cells (0.023% ± 0.004% versus 0.0180% ± 0.002%, respectively) (Fig. 5K–M).

Phenotypic and functional characterization of cultured fetal and adult corneolimbal epithelia. Double immunofluorescence for K14 (green) and αv-integrin (red)
Growth and phenotype of corneolimbal epithelia on contact lenses
Finally, fetal epithelia were cultured on contact lenses to determine whether they sustained these cells as we have previously demonstrated for adult equivalents [28, 33]. Fetal and adult cells adhered to the contact lens polymer, proliferated (Fig. 6A, C), and retained a K14 phenotype (Fig. 6B, D). The proportion of K14-expressing cells was not significantly different (P = 0.31) between fetal and adult epithelia (11.5% ± 0.4% versus 10.9% ± 0.2%, respectively) (Fig. 6E) and was similar to cells cultivated on vitronectin (Fig. 5G, H).
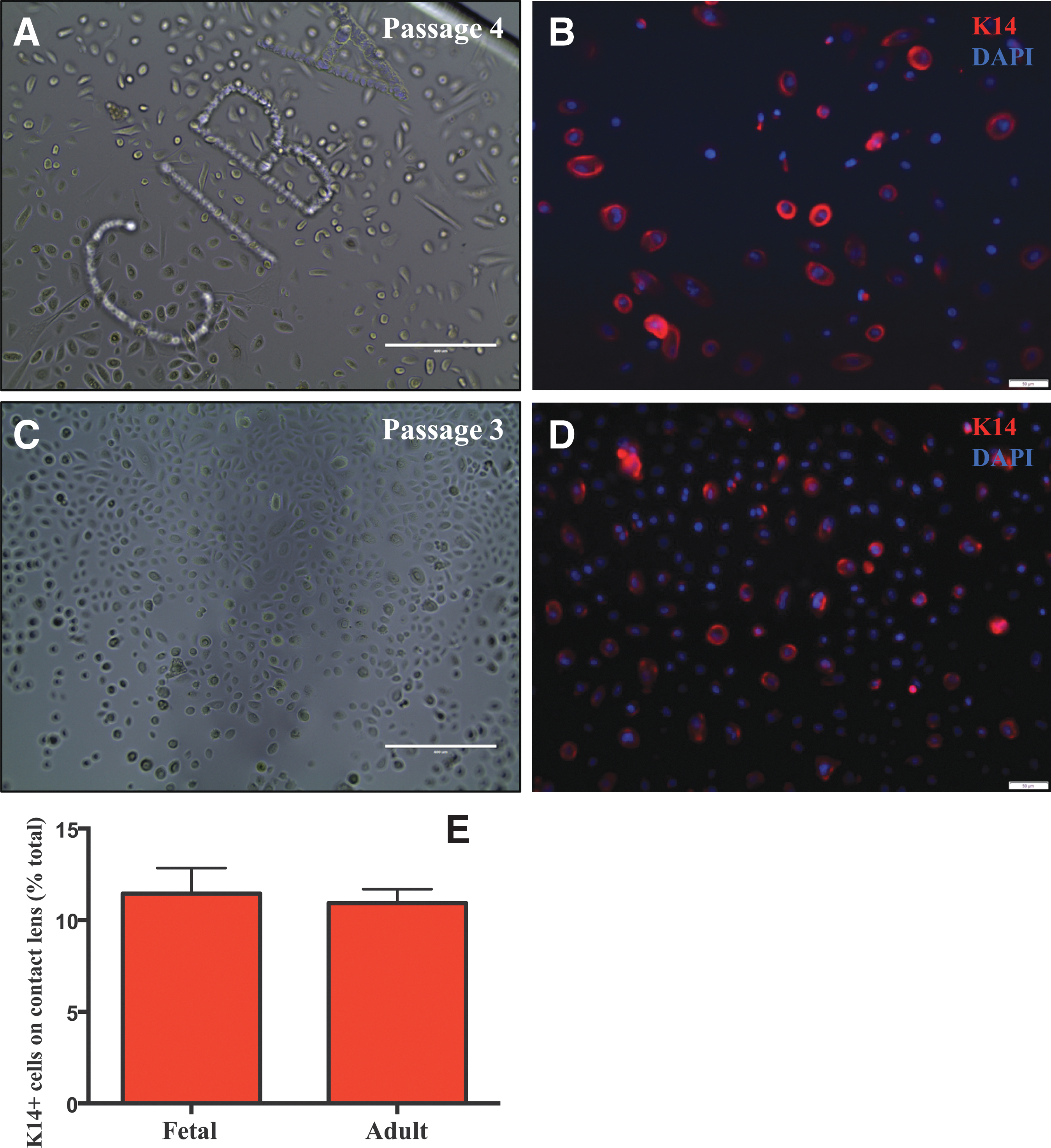
Growth and phenotypic characteristics of fetal and adult corneolimbal epithelia on contact lenses. Phase contrast microscopy was used to record the morphological appearance of fetal
Discussion
Our histological and histochemical findings provide key insights into the developmental dynamics of the human cornea, in particular the formation of the primordial SC repository, and the distribution of precursor cells across the ocular surface. The increased number of epithelial layers in the periphery, which was only apparent at 12 WG, is consistent with our previous report on the formation of the rudimentary LESC residence [20]. Interestingly, the palisades of Vogt, which characterize the adult limbus, were not detected in fetal or neonatal corneas. These structures have not been identified in human corneas up to 41 WG [20, 22], suggesting maturation of the limbal niche, and the distribution of its SCs extends well into postnatal life. In the present study, progenitor cell activity, as denoted by K14 staining, was initially widespread across the corneal surface and then consolidated horizontally to the limbus in neonatal and adult corneas. These findings are consistent with those of Davies et al. [20], which divulged a similar pattern for K15; however, a different temporal expression profile was noted for this marker. Notably, studies performed on the developing mouse cornea [14, 23, 24, 34] also support the notion that SCs are initially distributed across the ocular surface but eventually become confined to the limbus with time postconception.
Another intriguing observation was the consolidated vertical K14 staining within the epithelial layers of the developing limbus and central cornea. From the earliest fetuses investigated, K14 activity was evident in the superficial layers across the entire ocular surface spanning the central and peripheral zones. At 20 WG, this pattern transitioned to full thickness in the limbal dome, then regressed from the superficial layers at 23 WG, and became basal in the neonatal cornea similar to its expression in adult tissue. The reason for this differential expression is not known, but it is likely that the molecular programs that govern SC positioning, movement, and differentiation are concurrently executed during this critical developmental period in preparation for eyelid opening, which occurs at 26 WG in humans. We postulate that before exposure to amniotic fluid, the superficial epithelium of the cornea becomes differentiated and K14 precursor cells segregate distally, where they can be further protected. The molecular cues and niche-specific factors that govern SC activity warrant further exploration since their identity may be critical for devising ideal culture conditions for preserving stemness. Certainly, similar observations have been recorded in the embryonic and neonatal mouse [26, 27], where K14-positive cells populate the limbal zone as well as cells nestled within a unique structure called “compound niches.”
Another aim of this study was to identify reliable adult LESC markers that could serve to locate fetal counterparts. As previously noted by us [31] and others [35], K14 was restricted to the human basal limbal epithelium and highly expressed in cells isolated from the adult limbus [36]. Results from our qRT-PCR showed a similar pattern of K14 in cultured fetal and adult corneolimbal cells, with higher K14 mRNA compared with αv-integrin (data not shown). Moreover, we recently determined the long-term clonal expansion of K14+ cells in adult transgenic Confetti reporter mice using lineage tracing [37]. Overall, these data imply that K14 marks a population of long-living progenitor cells. However, one disadvantage is its intracellular location, rendering it unsuitable as a tethering tool for LESC enrichment. Among the other markers examined, Hes1, Notch1, and αv-integrin were consistently coexpressed with K14 in fetal and adult limbal tissues (Fig. 3) and cultured cells (Fig. 5). Although Notch1 is a surface receptor, ligand-dependent engagement results in its enzymatic cleavage, nuclear translocation, and activation of downstream transcription events, including targeting the basic helix–loop–helix family of proteins, such as Hes1 [38]. Therefore, due to its intracellular translocation, Notch1 may not be ideal for isolating cells, despite its specificity for basal limbal epithelia [39]. αv-integrins appear to be particularly important during tissue remodeling associated with wound repair and angiogenesis [40] and are also required for the long-term self-renewal of murine embryonic SCs [41]. αvβ5-integrin is a receptor for vitronectin, an extracellular matrix protein localized adjacent to SC-like clusters within the human limbus, and an in vitro LESC support factor [32, 42]. In these studies, αvβ5-integrin was used to enrich adult LESCs by fluorescence-activated cell sorting. Similar strategies have assisted the isolation of viable adult LESCs using α6-integrin [43]. We suspect αv will be a promising cell surface antigen to assist in isolating and enriching SCs from the developing ocular surface.
The importance of K14 and αv-integrin for the cornea and its SC population is exemplified in genetic ablation studies that have resulted in ocular sequelae. For example, K14 null mice develop thinner than normal corneal epithelia [44], and loss of α9β1-integrin in K14+ corneal and cutaneous cells results in poor reepithelialization capacity [45]. In addition, epithelial-specific knockdown of αv-integrin results in conjunctival and eyelid tumors [46], and mice lacking αvβ5-integrin develop retinal blindness [47]. These observations support the notion that K14 and integrins (especially αv) are critically important for eye health and vision.
Since 1997, numerous studies have identified the beneficial effects of cultured LESC therapy for treating patients with LSCD [8, 48 –52]. However, since none of the nonlimbal lineages used in clinical trials has proved to be successful in more than about 75% of cases [51, 52], and because long-term preservation of donor corneas in Eye Bank Organ Culture Media affects LESC viability [53], the search for alternative cell types that are more efficacious is ongoing. Clinicians are inclined to use autologous cellular sources as the risk of rejection is minimal, and there is no requirement for immunosuppression. Although fetal SCs are from an allogeneic source, these cells have reduced antigenicity due to their expression of HLA-G antigen for immune tolerance during pregnancy and are therefore less likely to be rejected following grafting [54]. These cells have also been extensively used in the treatment of various diseases for more than 90 years [55 –57]. Another major advantage is the fact that fetal corneal SCs are preprogrammed to differentiate and function as adult corneal epithelia. However, there would be significant ethical and legal hurdles to overcome before these cells are considered for treating severe corneal diseases.
This study offers novel insights into the formation of the limbal niche and the spatiotemporal distribution of SC activity across the developing human ocular surface. It also identifies K14 and αv-integrin as two adult LESC markers, which were used to locate fetal corneal SCs. Initial in vitro investigations demonstrate that limbal epithelial cells can be propagated and possess phenotypic and functional properties comparable to their adult analogues, and results from our colony-forming assays indicate that fetal corneal epithelial cells form colonies at a similar rate compared with their adult counterparts. In addition, they were successfully cultivated on a common therapeutic contact lens scaffold on which they maintained a K14+ progenitor cell phenotype. In the future, it will be our goal to determine their SC content with a view to using these cells as a potential therapy for patients with LSCD.
Footnotes
Acknowledgments
Grant sponsorship: University of New South Wales. The authors thank Mr. Raj Devasahayam at the Lions NSW Eye Bank for providing the donor human adult corneas.
Author Disclosure Statement
The authors (Y.E., A.R., M.K., B.H., G.J.G., D.W., and N.D.) have no conflict of interest either financial or personal to declare.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.