
Abstract
In-depth understanding of human spermatogenesis requires studying specific molecular signatures and interactions of spermatogonia with other testicular cell populations, for which isolation of pure populations of different cell types is crucial. Here, we describe a technique to simultaneously enrich pure, multiple testicular cell populations, including spermatogonia, endothelial (TECs), and perivascular mesenchymal stem/stromal cells (TMSCs), from testicular tissue by flow cytometry using a combination of defined markers. Immunohistochemical studies, multicolor staining, and cell sorting followed by multiplex quantitative real-time polymerase chain reaction (qRT-PCR) analysis revealed that spermatogonia were highly enriched in the CD49f+CD49a−HLA-ABC−SSEA-4+ fraction of primary testicular cells. In contrast to spermatogonia, TMSCs and TECs were highly enriched in the CD49f+CD49a+HLA-ABC+CD144− and CD49f+CD49a+HLA-ABC+CD144+subsets, respectively. The delineation was confirmed by the expression of specific stromal and endothelial key markers as well as by the differentiation and angiogenic capacity of the sorted populations. In this article, for the first time, we performed transcriptome profiling of highly enriched, freshly isolated human spermatogonia and compared their expression profile with that of TMSCs. Our RNA sequencing data favor the hypothesis that TMSCs are candidate niche components for spermatogonia. The composite genotype and phenotype of defined testicular cell populations combined with a robust isolation procedure from small biopsies contributes to a better understanding of cellular interactions and for the establishment of efficient culture techniques to maintain spermatogonial progenitors.
Introduction
I
Although Sertoli cells have an established role in regulating the fate of spermatogonia, cells in the peritubular compartment constituting of myoid cells, leydig cells, vascular testicular endothelial cells (TECs), and perivascular testicular mesenchymal stromal cells (TMSCs) are postulated to have direct effects on the growth and function of spermatogonial progenitor cells (SPCs) [1]. However, the role of TMSCs as a niche component is not yet elucidated, because the isolation of perivascular TMSCs in testis is difficult due to the lack of suitable markers to purify these cells. In contrast, several strategies have been developed to facilitate the prospective isolation of human spermatogonia from adult human testis based on the selective expression or absence of surface markers. Human spermatogonia are reported to express the surface markers CD49f, FGFR3, GFRA1, GPR-125, SSEA-4, SUSD2, CD90, and CD164 [2 –7]. Of these markers CD49f, CD90, and SSEA-4 were initially introduced to isolate human spermatogonia [4,5], whereas later GFRA1 and GPR-125 have been shown to be suitable for the prospective isolation of subsets of spermatogonia [3,8,9]. But most of these markers, including CD49f and CD90, are much less specific for spermatogonia than SSEA-4 and CD164 [3,4,7]. Another drawback for the isolation and characterization of different cell types from adult human testis is the limited availability of specimens of normal human testis.
In the first part of this study, we used a large panel of mAbs to establish robust markers for the simultaneous isolation of SPCs, TMSCs, and TECs from biopsies of adult human testis. We show that these populations within complex samples can be simultaneously isolated by co-staining of cells with lineage-specific antibodies, allowing unprecedented depth of analysis of heterogeneous cell populations. In continuation of our previous study, we introduce HLA-ABC as a marker to distinguish germ cells from somatic cells in testis. In addition, we provide a staining protocol to simultaneously isolate spermatogonia, TMSCs, and TECs in a single sorting step. In the second part of the study, we compared the transcriptome of freshly isolated spermatogonia and TMSCs by next-generation sequencing. Our analysis provides novel insights into the nature of the spermatogonia and the possible molecular interactions with TMSCs in vivo.
Methods
Tissue collection, ethics approval, and cell isolation
Human testicular tissue from patients under the regiment of hormonal therapy, undergoing transsexual sex reassignment surgery, subcapsular orchiectomy due to prostrate cancer, or inguinal orchiectomy due to testicular cancer was obtained after written informed consent and approval of the Ethics Committee (Nos. 273/2006 and 493/2008A) of the University of Tübingen. The ages of the patients ranged from 30 to 76 years, and the samples showed a varied degree of spermatogenesis ranging from regular mature spermatogenesis, mature spermatogenesis but reduced by 30%–60%. Samples showing no spermatogenesis were excluded from the study. From the surgical samples, testicular tissue was extracted in a sterile manner. The tissue was mechanically dissociated with a tissue chopper (Ted Pella, Inc., Wetzlar, Germany) and enzymatically digested in HBSS with Ca2+ and Mg2+ (PAA, Cölbe, Germany) containing 250 μg/mL of DNAse II (Sigma-Aldrich, Munich, Germany), 250 μg/mL of Dispase II (Roche, Mannheim, Germany), and 750 FALGPA units/mL of Collagenase XI (Sigma-Aldrich) for 90 min at 37°C. The digest was filtered through 100 and 40 μm sieves to remove undigested tissue. The cells were centrifuged at 1,000 rpm for 10 min, and pellets were incubated in ammonium chloride (Sigma-Aldrich) for 10 min on ice for selective lysis of erythrocytes. The cells were washed twice with PBS and maintained on ice for further use.
Cell culture
TMSCs were plated and expanded in 0.1% gelatin-coated dishes with Embryonic Stem cell (ES) media containing KnockOut DMEM supplemented with 20% KnockOut serum replacement (Invitrogen, Karlsruhe, Germany), 1% penicillin-streptomycin, 1%
Flow cytometric analysis and cell sorting
The details of antibodies used for FACS analysis and sorting are described in the Supplementary Data; Supplementary Data are available online at
Gene expression analysis by multiplex quantitative real-time polymerase chain reaction
Multiplex quantitative real-time polymerase chain reaction (qRT-PCR) of freshly isolated sorted populations was performed directly without any culture by using a 48.48 Dynamic array integrated fluidic circuit in the BioMark™ HD System. Details of this procedure are presented in the Supplementary Data.
Transcriptome analysis and genome-wide genotyping
Approximately 2.5 × 104 freshly sorted cells from CD49f+HLA-ABC−SSEA-4+ spermatogonial population and CD49f+HLA-ABC+CD49a+CD144− TMSC population were used for commercial RNA sequencing (RNA-Seq) analysis by using illumina next-generation sequencing technology (MFTService, Tübingen, Germany). RNA was isolated directly from the sorted populations without any culture by using Qiagen RNeasy-mini kit (Qiagen, Valencia, CA). The preparation of cDNA libraries was performed by using True-seq RNA and cDNA sample preparation kit v2 (Illumina, San Diego, CA), following the prescribed guidelines in the manual [12]. The sequencing was performed on cells isolated from two patient samples (two biological replicates). The detailed procedure is described in Supplementary Data.
Differentiation and angiogenesis assays
TMSCs derived from the CD49f+HLA-ABC+CD49a+CD144− sorted population were subjected to osteogenic, adipogenic, and chondrogenic differentiation (n = 3). An in vitro matrigel angiogenesis assay was utilized to assess the tube-formation capabilities of TECs derived from CD49f+HLA-ABC+CD49a+CD144+ sorted cells (n = 3) [13]. The procedure used is described in the Supplementary Data.
Immunohistochemical staining
The tissue was fixed overnight in 4% paraformaldehyde (Sigma-Aldrich), equilibrated the next night in 20% sucrose, and frozen in Tissue Tek OCT compound (Sakura). Immunohistochemical staining was performed on 5 μm cryosections. The sections were washed thrice in Tris-buffered saline (TBS) for 5 min and blocked for 30 min with TBS containing 0.1% Triton X-100, 10% goat serum, 0.1% sodium azide, and 0.1% cold fish skin gelatin (CFSG). The sections were stained with isotype controls (negative control) or purified antibodies diluted in staining buffer containing TBS supplemented with 5% goat serum, 0.1% sodium azide, and 0.1% CFSG for overnight at 4°C. The sections were washed three times for 5 min with TBS containing 0.05% Tween 20 (TBST), followed by staining with fluorophore-conjugated secondary antibody diluted in staining buffer. The sections were washed three times for 5 min with TBST and mounted with 4′6′-diamidino-2 phenylindole (DAPI) containing Vectashield hard set mounting medium (Vector laboratories, Peterborough, United Kingdom). The slides were visualized by using a Zeiss Observer.Z1 AX10 microscope with ApoTome and AxioVision 4.8 imaging software.
Statistics
The RT-qPCR data are represented as mean ± standard deviations for at least duplicates for each experiment. For analysis of RNA sequencing, significance of differential expression was determined by testing the negative binomial model. Test P values were corrected for multiple testing by the FDR procedure. A cutoff of 0.001 was chosen as significant.
Results
Isolation and enrichment of spermatogonia, TMSCs, and TECs
We have recently shown that spermatogonial progenitors and somatic cells in human adult testis are present in the CD49f+CD49a−SSEA-4+ and CD49f+CD49a+SSEA-4− fractions, respectively [7]. Based on the knowledge about the distribution of extracellular matrix, especially laminin in testis, it is evident that apart from spermatogonia, many other cell types in testis also associate with laminin and, hence, express the laminin receptor CD49f [14]. For in-depth characterization of spermatogonia and for purifying the other somatic cell types in CD49f+CD49a+SSEA-4− fraction, which are attached to the laminin, we screened a large panel of 327 antibodies against surface antigens for their reactivity with germ cells (CD49f+CD49a−SSEA-4+) and somatic cells (CD49f+CD49a+SSEA-4−), using single-cell suspensions prepared from testicular samples (Supplementary Table S1). Flow cytometric analysis revealed that antibodies against HLA-ABC and CD144 specifically reacted with subpopulations of CD49f+ or CD49f− somatic cells in the testis, but not with the germ cells (Fig. 1A, B).
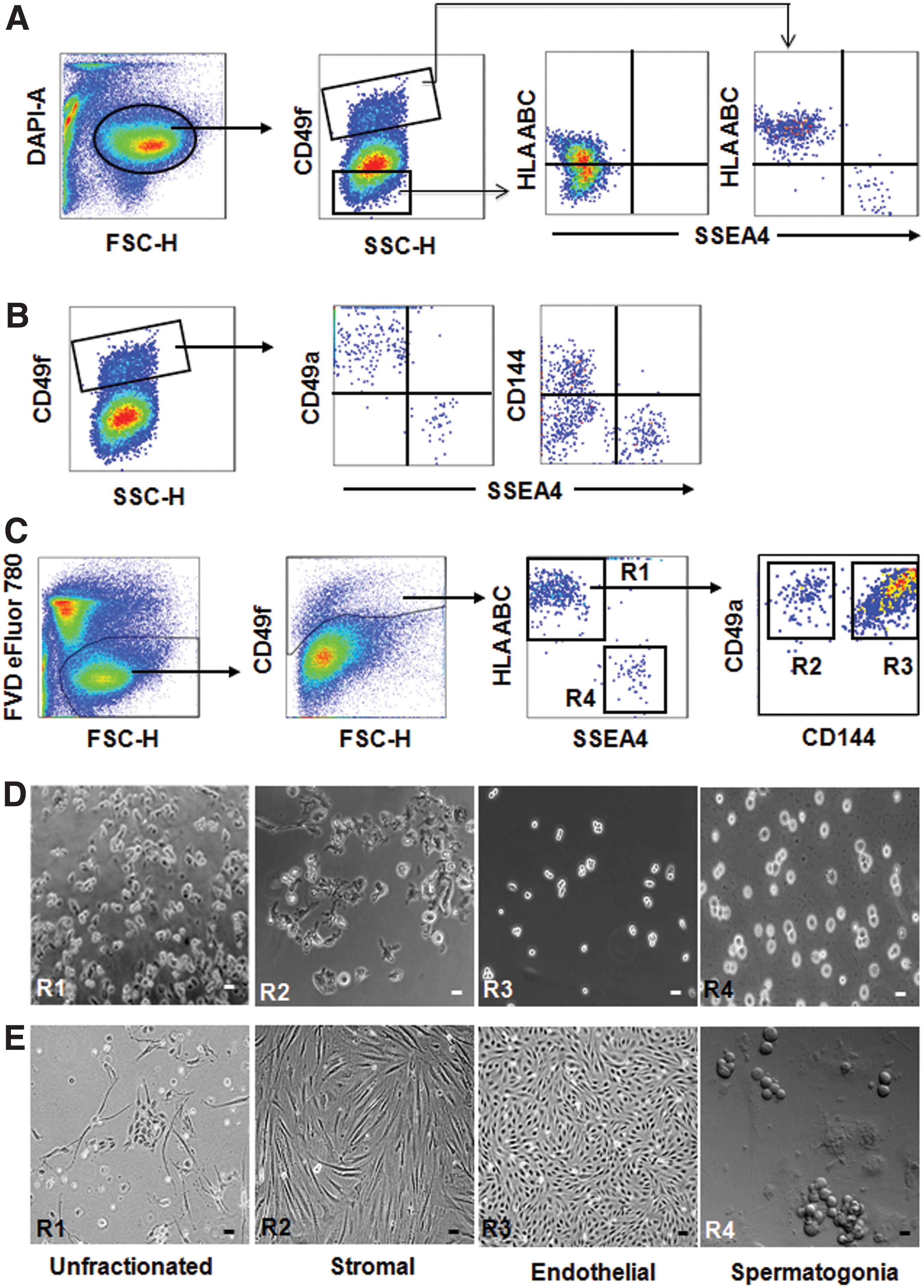
Isolation of different cell types from adult human testicular tissue.
Multicolor staining with antibodies against CD49f, CD49a, CD144, SSEA-4, and HLA-ABC, followed by flow cytometric analysis and FACS sorting, revealed that spermatogonia are exclusively found in the CD49f+HLA-ABC−CD49a−SSEA-4+ population (Fig. 1C). These cells were morphologically distinct from CD49f+HLA-ABC+CD49a− cells and identified as round cells with a large nucleus-to-cytoplasm ratio as verified by a microscopic examination of the morphology of sorted cells [4] (Fig. 1D). After initiation of culture in germ cell media, spermatogonia formed grape-like clusters (Fig. 1E) after 7 days. We, additionally, found that CD144 is a suitable marker to further fractionate the CD49f+HLA-ABC+CD49a− subpopulation into CD49f+HLA-ABC+CD49a+CD144− and CD49f+HLA-ABC+CD49a+CD144+ cells, respectively (Fig. 1C). Unlike spermatogonia, these cells showed an irregular morphology (Fig. 1D). The sorted cells were plated into culture flasks at defined cell numbers. Enumeration of the morphology of resulting colonies indicated that TMSCs were enriched in the CD49f+HLA-ABC+CD49a+CD144− subset and TECs in the CD49f+HLA-ABC+CD49a+CD144+ subset. The CD49f+HLA-ABC+CD49a+CD144− cells (TMSCs) were cultured in ES media and gave rise to fibroblast-like colonies (Fig. 1E). The percentage of CD49f+ cells in the total testicular cells is 9.1% ± 3.7%. The percentage of each of the isolated populations, including TMSCs, TECs, and SPCs, in the total testicular cells varies from 0.49 ± 0.29, 6.08 ± 1.4, and 0.15 ± 0.05, respectively. These estimates were made from six samples. As CD49f+HLA-ABC+CD49a+CD144+ cells (TECs) failed to proliferate in ES media, they were cultured in media suitable for endothelial cell growth (Vasculife cell culture media coated with 1% gelatin). The TECs displayed a cobblestone-like morphology (Fig. 1E) similar to human umbilical vein endothelial cell (HUVEC).
Localization of CD49a, CD144, and SSEA-4 in adult human testis
An immunohistochemical examination of adult human testicular tissue shows that almost all cells adjacent to the basement membrane of seminiferous tubules expressed CD49f (Supplementary Fig. S1A). SSEA-4 expression was confined to the cells located adjacent to the basement membrane of the tubules. The majority of the cells positive for SSEA-4 in the tubule expressed DAZL and DDX4, indicating a germ line-specific expression of SSEA-4 (Supplementary Fig. S1B, C). In contrast, CD49a (perivascular) and CD144 (vascular) were expressed only in the peri-tubular region and did not co-localize with DDX4 within the tubules, indicating that these markers are expressed on somatic cells but not germ cells (Supplementary Fig. S1D, E).
Gene expression pattern of isolated cell populations of testis
To further confirm the identity of each of the sorted populations, we performed gene expression analysis by multiplex qRT-PCR in three biological samples. The relative gene expression of the tested genes in different sorted populations in one patient sample is shown in Fig. 2A–D, and two other samples are shown in Supplementary Fig. S2. The expression of germ cell-specific markers DAZL and DDX4 was highly enriched in the CD49f+HLA-ABC−CD49a−SSEA-4+ population compared with CD49f+HLA-ABC+CD49a+CD144− and CD49f+HLA-ABC+CD49a+CD144+ cells, confirming the germ cell origin of these cells. In addition, this population was specifically enriched for the spermatogonia-specific markers UTF1, DSG2, and GFRA1 as well as other markers, including TERT and TSPY1. In contrast to spermatogonia, the fibroblast/stromal cell-specific markers FN1, VIM, COL1A1, and COL1A2 were enriched in CD49f+HLA-ABC+CD49a+CD144− cells but not in CD49f+HLA-ABC+CD49a+CD144+ or CD49f+HLA-ABC−SSEA-4+ cells, confirming the stromal nature of these cells. Similarly, the expression of the endothelial cell-specific markers MMRN1, ESM1, and PECAM1 was significantly higher in CD49f+HLA-ABC+CD49a+CD144+ cells than the other two populations, supporting the endothelial nature of these cells.

Relationship of gene expression patterns among different sorted populations from adult human testis.
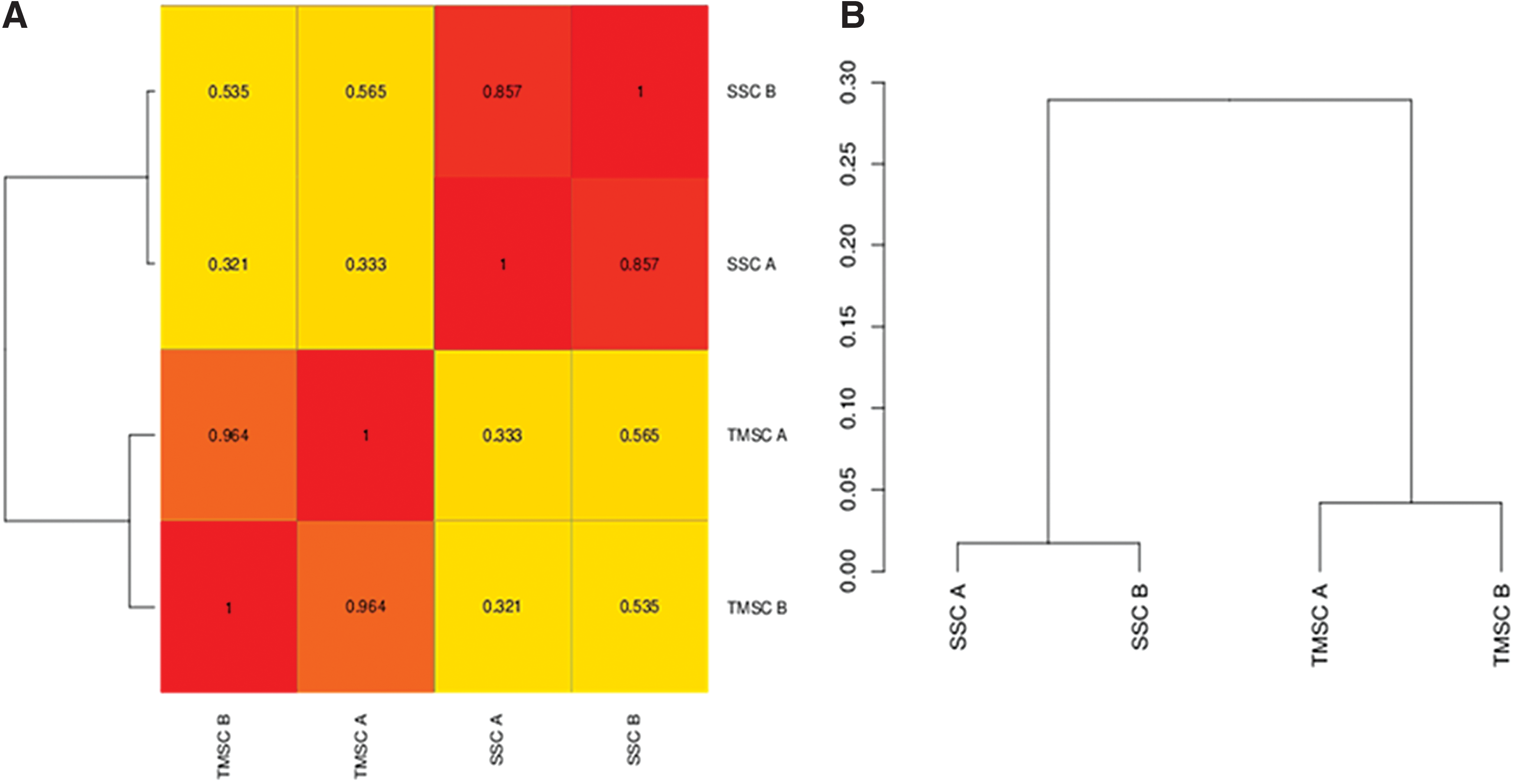
To further explore the relationship among the different sorted populations, we performed dendrogram cluster analysis based on the expression of 14 genes, as shown in Fig. 2A–D. Depending on the branching pattern in the dendrogram, the different sorted populations from the primary testis can be organized into three main groups, namely spermatogonia/germ cells, perivascular fibroblasts/stromal cells, and endothelial cells (Fig. 2E). CD49f+HLA-ABC+CD49a+CD144− cells along with cultured human fore skin fibroblasts (positive control) clustered together as fibroblasts/stromal cells and CD49f+HLA-ABC+CD49a+CD144+ cells along with HUVEC (positive control) clustered together as endothelial cells. Spermatogonia along with the human germ cell tumor (seminoma) derived CD49f+PLAP+ cells (positive control) clustered together as germ cells. As expected, unfractionated cells as well as cells of the CD49f+HLA-ABC+ subset (R1 in Fig. 1C), which contain a mixed population of cells, clustered together with somatic cells (either fibroblasts or endothelial cells).
Phenotype of primary spermatogonia
To analyze the expression profiles of surface markers on spermatogonia, primary testis cells were stained with CD49f and SSEA-4 along with markers known to be present or absent on spermatogonial stem cells. FACS analysis revealed that the majority of CD49f+HLA-ABC−SSEA-4+ spermatogonia express CD164, CD9, SUSD2, and CD326 and a subpopulation expresses CD90. In contrast, most of CD49f+HLA-ABC−SSEA-4+ spermatogonia did not express CD117, thus confirming that CD49f+HLA-ABC−SSEA-4+ cells contain spermatogonia (Fig. 3A, B).
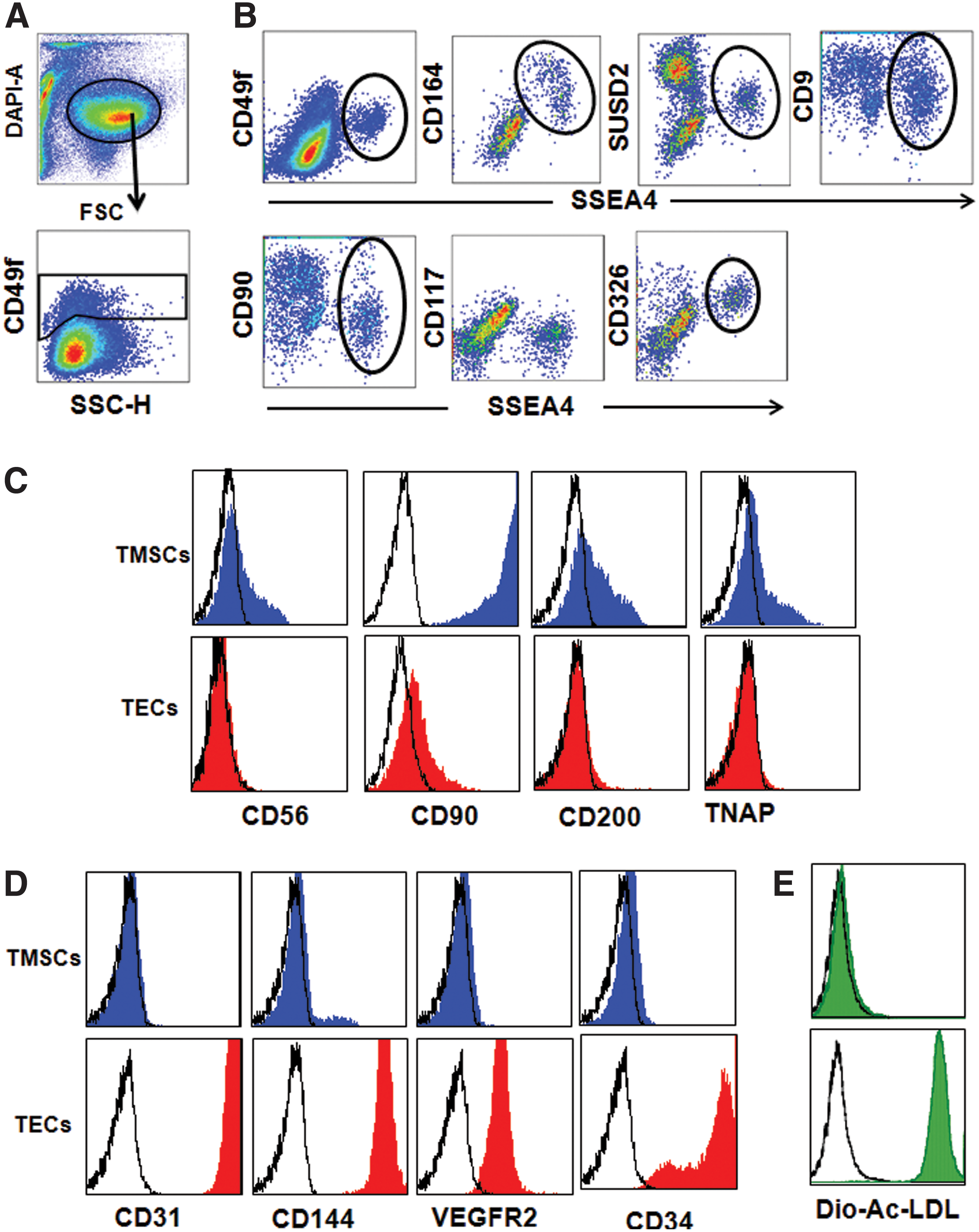
Phenotype of testis-derived SPCs, stromal cells, and endothelial cells.
Phenotype of TECs and TMSCs
After sorting, cells of the CD49f+HLA-ABC+CD49a+CD144− and CD49f+HLA-ABC+CD49a+CD144+ subsets were cultured in ES media and endothelial cell culture media, respectively. After 10 days, the cells were analyzed for stromal and endothelial-specific markers. Figure 3C shows that cells derived from the CD49f+HLA-ABC+CD49a+CD144− subset predominantly expressed the mesenchymal/stromal cell-specific key markers CD90 and CD200, whereas CD56 and TNAP were expressed only in subpopulations of cultured stromal cells. In contrast, cells derived from the CD49f+ HLA-ABC+CD49a+CD144+ subset were enriched for TECs, as verified by their selective expression of the endothelial cell-reactive key markers CD34, CD31, CD144, and VEGFR2 (Fig. 3D). In conclusion, TMSCs are enriched in the CD49f+HLA-ABC+CD49a+CD144− subset and TECs are exclusively found in the CD49f+HLA-ABC+CD49a+CD144+ subset. This was further confirmed by the selective uptake of Dio-Ac-LDL by TECs but not by TMSCs (Fig. 3E).
Differentiation capacity of TMSCs and angiogenic ability of TECs
To analyze the differentiation capacity of TMSCs and TECs, sorted CD49f+HLA-ABC+CD49a+CD144+/− cells were expanded until passage 2 in ES medium or Vasculife cell culture medium. Defined numbers of the resulting TMSCs were then induced to differentiate into cells of the osteogenic, adipogenic, and chondrogenic lineages. TMSCs were able to differentiate into osteoblasts and chondrocytes as verified by Alizarin Red S staining and Alcian Blue staining, respectively (Fig. 4A). Surprisingly, TMSCs were unable to differentiate into adipocytes, which is in contrast to the capacity of bone marrow-derived mesenchymal stromal cells. These cells, which were used as a positive control, were, indeed, able to differentiate into osteogenic, adipogenic, and chondrogenic lineages (Fig. 4B).
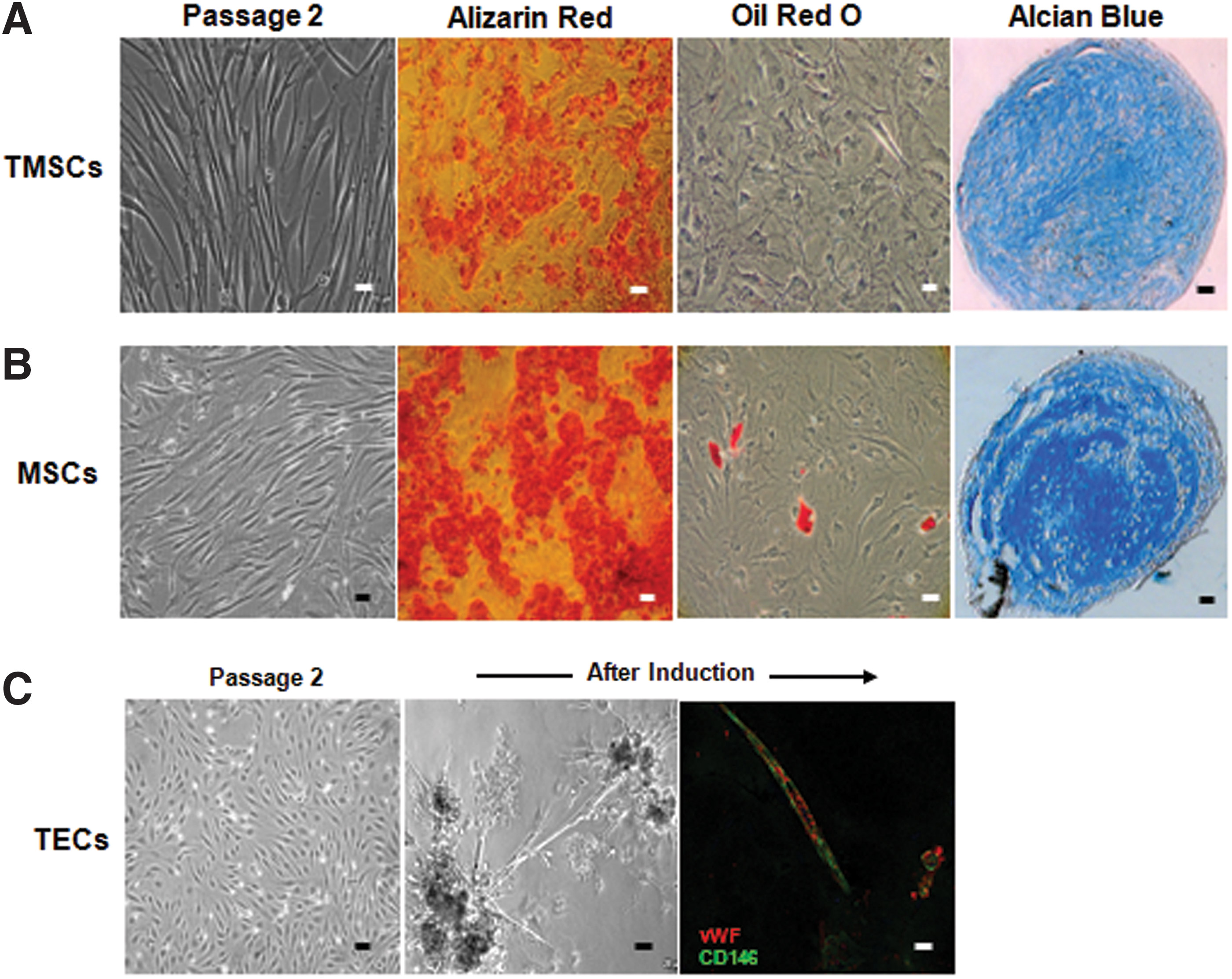
Differentiation potential of TMSCs and angiogenic ability of TECs.
The angiogenic capability of TECs was assessed by using an in vitro endothelial tube formation assay. After 24 h of induction of tube formation on matrigel, TECs exhibited capillary-like interconnected structures (Fig. 4C). To further confirm the endothelial differentiation of TECs, cells on matrigel were stained with CD146 and vWF and visualized under a fluorescence microscope. The data presented in Fig. 4C demonstrate significant CD146 and vWF staining of TECs under induction culture conditions, further supporting their endothelial commitment.
Genome-wide expression profiling of sorted testicular somatic and spermatogonial progenitors
After we confirmed the identity and purity of the isolated cell populations, we performed RNA sequencing (MFTService) to identify the molecular signatures of freshly isolated spermatogonia (SPCs) and TMSCs, to determine the possible molecular interaction between the spermatogonia and TMSCs.
Hierarchical clustering and correlation heatmap reveal that SPCs and TMSCs cluster separately (Fig. 5A, B). Germ cell-specific genes, including genes needed for the formation of P granule, pole plasm, spermatogenesis, germ stem cell maintenance, and stem cell maintenance, were upregulated in the CD49f+HLA-ABC−SSEA-4+ spermatogonial population, confirming their spermatogonial progenitor characteristics (Supplementary Table S2). Fibroblast-specific genes were highly upregulated in the CD49f+HLA-ABC+CD49a+CD144− TMSC population, confirming their stromal-like nature (Supplementary Table S3). Furthermore, smooth muscle cell-specific genes, including ACTA1, ACTA2, MYLK, PPP1R21, SMTN1, CNN1, and NMU, were not upregulated in the TMSC fraction (Supplementary Raw Data 1), demonstrating the purity of the isolated cells, and excluding a potential contamination with smooth muscle or myoid cells. The P values of all the genes listed in Supplementary Raw Data 1 are listed in Supplementary Raw Data 2.
We performed pathway analysis through KEGG (Kyoto Encyclopedia of Genes and Genomes) and Pathway commons database to find the genes involved in the crosstalk between TMSCs and SPCs. The most common enriched pathways in both the cell types are listed in Supplementary Tables S4 and S5. The genes, which encode for growth factors that are highly expressed by TMSCs, include FGF2, INHBA, KITLG, and FLT3LG (Supplementary Tables S3 and S5), which may be potentially involved in the maintenance and differentiation of the SPCs. The mRNA of the corresponding receptors of these factors, including FGF2, Activin, KITLG, and FLT3LG, are highly expressed in SPCs based on RNA-seq analysis, which are translated on differentiation of spermatogonia. The important positive downstream regulators of these receptors converge in intracellular signaling through TGFβ, PI3K/Akt, or Ras/ERK pathway and are upregulated in SPCs (Supplementary Tables S2 and S4). Apart from these factors described earlier, many other growth factors and cytokines are enriched in TMSCs but the corresponding receptors of these factors are not enriched in spermatogonia (Supplementary Tables S2–S5).
On the other hand, SPCs express PDGFB, which may possibly interact with the PDGFRB receptor expressed by TMSCs. The possible ligand-receptor interactions of spermatogonia and TMSCs are listed in Supplementary Fig. S3. Spermatogonia also express the FGFR3 receptor and its cognate ligand, FGF9, suggesting that both paracrine (FGF2) and autocrine (FGF9) signaling may activate the FGFR pathway in germ stem cells. In addition, SPCs express BMP7, its receptor BMPR1B, BMP7 antagonist NOG and BMP inhibitor BAMB1, which may be involved in positive or negative regulation of downstream TGFβ signaling through Smad1.
Discussion
One of the aims of the current study was to evaluate the most suitable marker combinations for the prospective isolation of spermatogonia and other somatic cell types within the CD49f+ fraction, which are associated with the basement membrane of adult human testis. We have identified cell-surface markers that serve as valuable tools for the simultaneous isolation of spermatogonia, TMSCs, and TECs attached to laminin in the basement membrane of human adult testis. The laminin receptor CD49f+ population contains spermatogonia and other somatic cell types attached to the basal lamina [4,14]. In a first step, to characterize these other cell types in depth, we prepared single-cell suspensions from testicular samples and screened a large panel of mAbs for their reactivity with CD49f+CD49a+SSEA-4− somatic cell fraction and CD49f+CD49a−SSEA-4+ spermatogonia. We identified HLA-ABC and CD144 as suitable targets to further fractionate the CD49f+CD49a+SSEA-4− population. As CD49a and CD144 are selective markers of the fibroblast and endothelial compartments [15 –17], we expected to separate most of these other somatic cell types from spermatogonia. Using this strategy, we were able to enrich spermatogonia to almost complete purity and could also separate fibroblasts and endothelial compartments. Using mAbs against CD49f, SSEA-4, and HLA-ABC, we could demonstrate that all CD49f+ spermatogonia are HLA-ABC− whereas most CD49f+ somatic cells are HLA-ABC+. In line with this finding, Dovey et al. showed that human spermatogonia do not express HLA-ABC [18]. They showed by multiparameter sorting of MOLT-4 contaminated human testicular cell suspensions employing antibodies against HLA-ABC, EpCAM, and CD49e that putative spermatogonia reside in the EpCAM+HLA-ABC−CD49e− fraction and the MOLT-4 cells reside in the EpCAM−HLA-ABC+CD49e+ fraction.
One of the markers, either HLA-ABC or SSEA-4 along with CD49f, is sufficient to isolate spermatogonia. Similarly, one of the markers, either HLA-ABC or CD49a along with CD49f, is sufficient to isolate all somatic cell populations attached to the basement membrane. But to ensure high enrichment, to prevent donor variability in the expression of any of these markers, and to prevent contamination of the germ cell population in the somatic cells and vice versa, we used these markers simultaneously. In this case, HLA-ABC serves as an important robust additional marker to separate germ cells from somatic cells.
Desmoglein 2 (DSG2), human telomerase reverse transcriptase (TERT), undifferentiated embryonic cell transcription factor 1 (UTF1), zinc-finger, BTB-domain containing 16 (ZBTB16), and GRR-α1 are reported as specific biomarkers of human spermatogonia [8,9,19 –23]. In particular, TERT, ZBTB16, and GFRA1 are expressed at high levels in undifferentiated spermatogonia, which decrease on differentiation [4,19,20,22]. In addition, UTF1 expression is known to decrease in human embryonic stem cells on differentiation [24]. In accordance to this, our study revealed that CD49f+HLA-ABC−CD164+ spermatogonia were highly enriched for DSG2, TERT, ZBTB16, GFRA1, and UTF1, which confirms their spermatogonial progenitor-like nature.
Human spermatogonia are reported to express CD49f, CD90, CD9, EpCAM, and SSEA-4 but not c-kit or HLA-ABC [4,18]. This is in line with our observation that CD49f+HLA-ABC−CD164+ spermatogonia are positive for CD9, CD90, SSEA-4, and EpCam and negative for CD49a, HLA-ABC, and c-kit. However, we could demonstrate that CD9 and CD90 are expressed not only on spermatogonia but also on many other cell types in the testis, indicating that these markers are not selective for spermatogonia. It was reported that CD49f is a suitable marker for the prospective isolation of spermatogonia by MACS [25]. In our studies, we observed that spermatogonia are CD49f dim whereas CD49fbright cells consist of mainly somatic cells. This raises a question as to whether the selection of spermatogonia by MACS with the single use of CD49f for isolation is sufficient to obtain pure spermatogonia.
Stem cells (SCs) from most tissues express HLA-ABC molecules at various levels. Di Trapani et al. showed that HLA-ABC molecules are more abundant in olfactory-ecto mesenchymal SCs and leptomeningeal SCs than in bone marrow-derived mesenchymal SCs, amniotic fluid SCs, cardiac SCs, and lung SCs [26]. However, after inflammatory priming, HLA-ABC molecules increased in all these SC categories. Similarly, other groups have shown low expression levels of HLA-ABC proteins on the surface of human ES cells that moderately increase after in vitro and in vivo differentiation [27,28]. In contrast to all these stem cell types, male germ cells do not express HLA-ABC. This is supported by our current work and other previous studies [18,29]. To the best of our knowledge, spermatogonia and oocytes are the only stem cells that completely lack HLA-ABC expression on their cell surface.
Testicular peritubular cells surround the seminiferous tubules and cooperate with Sertoli cells in the maintenance of the tubular architecture and in the formation of the blood-testis barrier [30,31]. Apart from these functions, they also participate in paracrine and endocrine signaling. For instance, fibronectin is a basement membrane protein known to be secreted in the seminiferous tubule by the peritubular cells [31]. In addition to fibronectin, peritubular cells secrete TGF-α, TGFβ, and IGF-1 [32]. However, in the peritubular cell population, the activity of the individual cell types (presumably Leydig cells, endothelial cells, smooth muscle cells, and perivascular stromal cells) cannot be discriminated from one another. Cellular and molecular analysis of pure populations of different cell types is necessary to clarify the role of these cells in the development and maintenance of testicular function. For this purpose, the combination of CD49a, CD49f, CD144, and HLA-ABC may serve as attractive biomarkers to prospectively isolate perivascular stromal and endothelial cell compartments. Though we have reported CD49f+CD49a−CD164− as the expression profile to isolate Sertoli cells in the testis [7], markers to isolate other cell types in the testis, including Leydig cells and smooth muscle cells, have yet to be established. Experiments to identify novel markers for the isolation of other cell types in testis are in progress, and our initial experiments indicate that SUSD2 may be an ideal target to isolate Leydig cells from the peritubular region of testis.
The continuous production of male gametes requires the precise regulation of the differentiation and replication of spermatogonial stem cells. These critical processes occur in a niche that is created in part by Sertoli cells, the peritubular cells including TMSCs, myoid, endothelial, and Ledyig cells. As yet, the in vivo mechanisms by which TMSCs regulate spermatogonial stem cell differentiation and maintenance have not been elucidated. In this study, based on the expression of growth factors, their receptors, and downstream signaling molecules, we propose that spermatogonia interact with TMSCs through paracrine signaling. The possible interactions of spermatogonia and TMSCs are depicted in Supplementary Fig. S3. INHBA and KIT ligand (KITLG, stem cell factor) produced by Sertoli cells are known to be involved in the differentiation of spermatogonia [33,34]. In addition, FGF2, which is also produced by Sertoli cells, is known to be involved in the maintenance of undifferentiated spermatogonia [35]. In our study, we show that TMSCs express the growth factors FGF2, INHBA, KITLG, and FLT3LG and spermatogonia express the corresponding receptors of these factors. Our results indicate that apart from Sertoli cells, TMSCs may interact with spermatogonia to control their proliferation and differentiation through the secretion of specific factors, thus contributing to the niche microenvironment of spermatogonia.
Although FLT3 and KIT are expressed at mRNA levels in SPCs, we did not find the expression of these receptors on the cell membrane at the protein level, as determined by flow cytometry. This might be due to translational repression of mRNA of these receptors in undifferentiated spermatogonia. It has been shown that NANOS encodes for a zinc-finger RNA-binding protein and gives rise to translational repression activity of many different mRNAs, including KIT [36,37]. On differentiation of SPCs, translational repression might be suppressed by downregulation of the expression of repressors, leading to translation of KIT and FLT3 mRNA.
Several studies have shown that the meiotic gatekeeper NANOS, an RNA-binding protein, silences genes that are essential for spermatogonial differentiation and meiotic entry by interaction with Pum2 [36,37]. FGF9 is known to be secreted by Sertoli cells and acts as a meosis-inhibiting substance by increasing NANOS levels in premeiotic spermatogonia. The autocrine signaling of FGF9 in spermatogonia is not known. In this study, we report that FGF9 and its receptor FGFR3, as well as its target NANOS2 and its interaction partner PUM2 are expressed by human spermatogonia (Supplementary Raw Data 1). This indicates that FGF9 may act via both autocrine and paracrine pathways for spermatogonial stem cell maintenance (Fig. 5).
Markers for combinatorial sorting are needed to improve the specificity of enrichment of different cell types from adult human testis. The antibodies described in this study provide refining cell enrichment strategies for SPCs, TMSCs, and TECs. In addition, our data provide novel insights into likely interactions between spermatogonia and TMSCs. Based on these interactions, we propose that TMSCs act as niche components of spermatogonia. In future, the novel enrichment strategies of SPCs may serve as a basis for an improved treatment of male infertility. Besides its application for therapeutic purposes, isolation of pure populations of different testicular cell types will help in defining the cues that support development and differentiation of spermatogonia. Finally, the knowledge about the composite phenotype of defined testicular cell subsets will contribute to a more accurate discrimination between healthy and malignant cells in the human testis.
Footnotes
Acknowledgments
The authors would like to thank Sabrina Grimm for the technical assistance in cell sorting and cell culture. This work was funded by the Forschungsprogramm der Tübinger Medizinischen Fakultät—Fortüne program (Project number: F1235109).
Ethical Conduct of Research
The authors state that the investigation was conducted after obtaining approval of the Ethics Committee (nos. 273/2006 and 493/2008A) of the University of Tübingen. Informed consent was obtained from individual patients whose samples were involved in the investigation.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.