
Abstract
The use of human mesenchymal stem cells (hMSCs) has emerged as a possible therapeutic strategy for CNS-related conditions. Research in the last decade strongly suggests that MSC-mediated benefits are closely related with their secretome. Studies published in recent years have shown that the secretome of hMSCs isolated from different tissue sources may present significant variation. With this in mind, the present work performed a comparative proteomic-based analysis through mass spectrometry on the secretome of hMSCs derived from bone marrow (BMSCs), adipose tissue (ASCs), and human umbilical cord perivascular cells (HUCPVCs). The results revealed that BMSCs, ASCs, and HUCPVCs differed in their secretion of neurotrophic, neurogenic, axon guidance, axon growth, and neurodifferentiative proteins, as well as proteins with neuroprotective actions against oxidative stress, apoptosis, and excitotoxicity, which have been shown to be involved in several CNS disorder/injury processes. Although important changes were observed within the secretome of the cell populations that were analyzed, all cell populations shared the capability of secreting important neuroregulatory molecules. The difference in their secretion pattern may indicate that their secretome is specific to a condition of the CNS. Nevertheless, the confirmation that the secretome of MSCs isolated from different tissue sources is rich in neuroregulatory molecules represents an important asset not only for the development of future neuroregenerative strategies but also for their use as a therapeutic option for human clinical trials.
Introduction
O
Indeed, preclinical in vitro [2 –5] and in vivo [6] studies have shown that the molecules secreted by different hMSC populations, namely bone marrow (BMSCs), adipose tissue (ASCs), and human umbilical cord perivascular cells (HUCPVCs), are able to: (1) promote neuronal survival and neurite outgrowth; (2) increase levels of neurogenesis and angiogenesis; (3) inhibit apoptosis and scarring; (4) modulate immune response; and (5) improve functional outcomes in different models of central nervous system (CNS) injury and disease, such as brain ischemia, spinal cord injury (SCI), Parkinson's disease (PD), and Alzheimer's disease (AD). In addition, these studies also revealed that these hMSC secretome-mediated cell processes contribute to the improvement of animals' functional recovery following hMSC transplantation. Nevertheless, although these different hMSC populations share similar phenotypical characteristics [7,8] and exhibit proregenerative potential, they reside in different anatomic parts of the body and, therefore, it is most likely that they present differences in their secretome.
In fact, Ribeiro et al. [9] conducted a screening on the presence of some neuronal survival and differentiation growth factors in ASC and HUCPVC secretomes, revealing important differences in the secretome composition between these two populations. While ASC CM was positive for the presence of hepatocyte growth factor (HGF), vascular endothelial growth factor (VEGF), stem cell factor, and nerve growth factor (NGF), only NGF and VEGF were detected in HUCPVC CM. On the other hand, Nakanishi et al. [8] demonstrated that there were significant differences in the growth factors and cytokines secreted by rat-derived ASCs and BMSCs. While ASCs secreted higher amounts of angiogenic and antiapoptotic growth factors, such as HGF and VEGF, as well as interleukin-6 (IL-6), BMSCs secreted cell migration-related chemokine stromal cell-derived factor 1 alpha (SDF-1α) [8]. Hsieh et al. [10] also revealed that although both supernatants collected from MSCs derived from Wharton's jelly (WJMSCs) and BMSCs contained angiogenesis-related factors, the secreted factors were distinct.
Nevertheless, although all of the above-referenced studies have established that there are important differences among the secretomes of different hMSC populations, the fact remains that only targeted proteomic approaches have been used for this purpose. Being so, these studies provide only a narrow window of the soluble factors secreted by MSCs. Given the vast panel of functional roles attributed to hMSCs in the mediation of paracrine actions that ultimately contribute for CNS repair, an unbiased, global proteomics approach would better clarify the potential complexity of hMSC secretome. In this sense, a shotgun/discovery proteomics-based approach, in which liquid chromatography (LC) is used to identify and quantify proteins present in hMSC secretome, offers a more broader analysis of hMSC-secreted proteins. Fraga et al. [11], using LC coupled with tandem mass spectrometry (LC-MS/MS), were able to identify and quantify the expression of several proteins in the HUCPVCs secretome that were, to date, not known to be secreted by these cells. Importantly, in this study, the authors identified proteins, such as 14-3-3, ubiquitin-carboxy-terminal hydrolase 1 (UCHL1), heat shock protein 70, and peroxiredoxin-6, which are involved in the regulation of neuronal cell survival/protection, proliferation, and differentiation.
Having this in mind, in the present work, we aimed to: (1) identify and quantify the expression of neuroregulatory proteins that might be related with the hMSC secretome-mediated processes involved in neuroprotection, neural repair, and neurodifferentiation and (2) evaluate to what extent the secretome of different hMSC populations (eg, BMSCs, ASCs, and HUCPVCs) can differ in the above-referred phenomena.
Materials and Methods
Cell culture
Human bone marrow, umbilical cord perivascular cells, and adipose tissue-derived stem cells
BMSCs (n = 3, Stem Cell Technologies) were thawed and expanded according to protocols previously described in our laboratory [12]. Briefly, BMSCs were cultured in α-MEM (Invitrogen) supplemented with 10% of fetal bovine serum (FBS, Biochrom) and 1% penicillin–streptomycin antibiotic (Invitrogen). After reaching 80%–90% of confluence, cells were enzymatically dissociated with 0.05% trypsin-EDTA (Invitrogen), plated at a density of 4,000 cells/cm2, and maintained at 37°C, 5% humidified CO2, 95% air, and 90% relative humidity. The culture medium was changed every three days.
HUCPVCs (n = 3) were kindly provided by Professor J.E.Davies (University of Toronto, Canada; Ethical approval had been previously obtained from the University of Toronto/Sunnybrook and Women's College Health Sciences Centre, Toronto) and ASCs (n = 3) by Professor J.M.Gimble (Pennington Biomedical Research Center/Tulane University; All protocols were reviewed and approved by the Pennington Biomedical Research Center Institutional Research Boards before the study, Tulane University). Cell isolation from umbilical cord and adipose tissue was performed according to the protocols previously described [13,14]. As for BMSCs, ASCs and HUCPVCs were cultured in α-MEM (Invitrogen) supplemented with 10% of FBS (Biochrom) and 1% penicillin–streptomycin antibiotic (Invitrogen). After reaching 80%–90% of confluence, cells were enzymatically dissociated with 0.05% trypsin-EDTA (Invitrogen), plated at a density of 4,000 cells/cm2, and maintained at 37°C, 5% humidified CO2, 95% air, and 90% relative humidity. The culture medium was changed every 3 days.
Conditioned medium collection and concentration
The CM used was collected from BMSC, ASC, and HUCPVC cultures in passage 6 (P6) as follows: cells were plated at a density of 12,000 cells/cm2 in T175 tissue culture flasks (Nunc) and allowed to grow for 3 days. After this, the flasks were first washed five times with phosphate-buffered saline without Ca2+/Mg2+ (Invitrogen), and then washed twice with Neurobasal A medium (Invitrogen). Following this, Neurobasal A medium supplemented with 1% kanamycin (Invitrogen) was added to the cells. After 24 h, the CM was collected and concentrated (100 ×) by centrifugation using a 5 kDa cutoff concentrator (Vivaspin™; GE Healthcare). All collected CMs were frozen at −80°C until it was required.
Proteomics–mass spectrometry and SWATH acquisition
Secreted proteins were precipitated from the concentrated medium using trichloroacetic acid (TCA; Sigma)-Acetone (Sigma) procedure [15]. Briefly, samples were incubated at −80°C with TCA [final concentration of 20% (v/v)] for 30 min and centrifuged (20,000 × g) for 20 min at 4°C. Protein pellets were then solubilized in ice-cold (−20°C) acetone, aided by ultrasonication (VC750; Vibracell-Sonics&Materials), and centrifuged (20,000 × g) for 20 min. The obtained pellets were thereafter resuspended in triethylammonium bicarbonate buffer (TEAB, 1M; Sigma), aided by ultrasonication, and centrifuged (20,000 × g) for 5 min to remove the insoluble material.
For liquid digestion, samples were reduced by the addition of 4 μL of tris(2-carboxymethyl)phosphine (TCEP, 50 mM; Sigma) to 45 μL of each sample, followed by an ultrasonication step for 2 min. Then, 2 μL of the cysteine blocking agent, methanethiosulfonate (600 mM; Sigma), was added and samples were left to react for 10 min at room temperature. TEAB was added to bring the final volume of each sample to 100 μL, and the samples were digested with trypsin overnight (2 μg trypsin/sample), at 37°C, with swirling at 560 rpm. Reactions were stopped by the addition of 2 μL of formic acid (FA; Amresco) and the peptides were dried by rotary evaporation under vacuum.
After protein digestion, samples were desalted using OMIX tips containing C18 stationary phase (Argilent Technologies). Eluted peptides spiked with iRT peptides (Biognosys AG) were dried and resuspended in a mobile phase containing 0.1% FA and 2% of acetonitrile (ACN) aided by ultrasonication (20% intensity). To remove insoluble material, samples were centrifuged (14,000 × g; 5 min) before LC-MS/MS analysis. Initially, peptide samples were resolved by LC using a C18 AR reverse phase column (ChromXP, 300 μm × 15 cm, 3 μm particle size, 120 Å pore size; Eksigent) at 5 μL/min using a 25-min ACN linear gradient (from 2% to 35%) in 0.1% FA into the mass spectrometer (Triple TOF 5600 system; AB SCIEX®).
For tandem mass spectrometry (MS/MS) analysis, samples were analyzed in two phases. First, each sample was analyzed with a mass spectrometer operating in information-dependent acquisition (IDA) to detect and identify the maximal number of proteins within sample mixtures. For IDA, the mass spectrometer was set to scan full spectra of ions in the 350–1,250 m/z range, during 250 ms, followed by 20 ion fragmentation spectra (MS/MS) scans (100–1,500 m/z range), with 1 MS/MS being acquired for 100 ms before adding those ions to the exclusion list for 20 s. The selection/isolation criteria for ion fragmentation comprised intensity, where ions had to meet a minimum threshold of 70 counts/s with a charge state between +2 and +5. Ions were fragmented in the collision cell (rolling collision) using collision energy spread of 5 eV.
The library was obtained by searching against the human and bovine species from SwissProt database, using the Protein Pilot™ software (v4.5; AB SCIEX). Afterward, samples were analyzed using the SWATH acquisition method, which allows detection and an accurate quantification of the identified proteins. For this purpose, the instrument was set to isolate ions with a 26 m/z width, in a loop mode, over the precursor mass range of 350–1,100 m/z, and 30 overlapping windows were constructed. For instrument calibration, the survey scan was set to scan full spectra of peptide ions in the 350–1,250 m/z range (50 ms) at the beginning of each cycle. SWATH fragmentation spectra were collected from 100–1,500 m/z using an accumulation time of 100 ms for all fragment ion scans, which resulted in a cycle time of 3.25 s. For optimal fragmentation of precursors within the isolation windows, a 15 eV spread of collision was applied. The SWATH quantitative information was extracted from the SWATH-MS data using the SWATH processing plug-in for PeakView™ (2.0.01 version; AB SCIEX). Peak areas were extracted (in an extracted ion chromatogram window of 1.5 min) for up to five target fragment ions (automatically selected) up to 15 peptides (selected based on a false discovery rate (FDR) lower that 1%) per protein. The levels of the proteins were estimated by summing all the transitions from all the peptides for a given protein [16] and were normalized to the total intensity of the sample. Quantification results are expressed as the average protein intensity that corresponds to the relative protein intensity in proportion to the internal standard (IS, GFP).
Statistical analysis
Statistical analysis was performed using one-way ANOVA with Bonferroni post hoc test for multiple group comparison, using the program GraphPad Prism five (GraphPad Software, Inc.). Data are presented as mean ± standard deviation, and differences were considered significant when P < 0.05.
Results and Discussion
To evaluate the variation of the hMSC secretome as a function of tissue source, the secretome of hMSCs derived from the bone marrow, umbilical cord, and adipose tissue was characterized through a proteomic approach based on SWATH-MS acquisition. From this analysis, we observed that BMSC, HUCPVC, and ASC secretomes disclosed a different profile (Fig. 1A). Through the use of Venn diagrams, we were able to identify a total of 451 proteins, in which 134 proteins were common to the three MSC populations (Fig. 1B). From these, 121 were quantified (Supplementary Table S1; Supplementary Data available online at
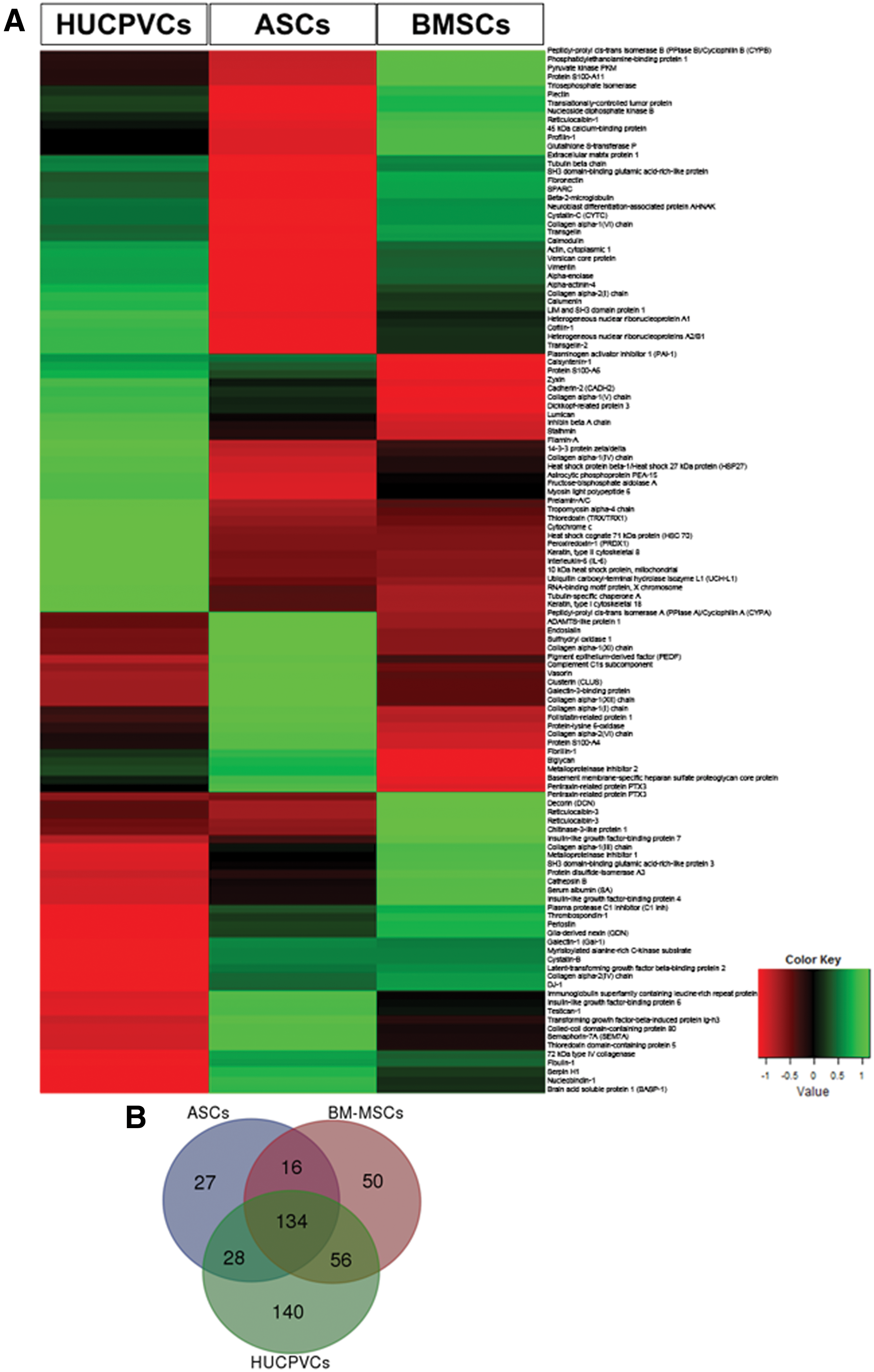
Graphical representation of HUCPVC, ASC, and BMSC CM proteomic analysis by mass spectrometry. CM analysis revealed that the BMSCs, ASCs and HUCPVCs had different secretome profiles
Protection against oxidative stress
Regarding oxidative stress, studies indicate that eight of the proteins found in the secretome of BMSCs, ASCs, and HUCPVCs, namely DJ-1, TRX, CYPA, CYPB, CYSC, PRDX1, AS, and HSP27, work as antioxidative mediators [22 –26]. For instance, DJ-1, which is a multifunctional protein from the peptidase C56 family, may become activated in the presence of reactive oxygen species (ROS) under conditions of oxidative stress, acting as a transcriptional regulator of antioxidative gene batteries [27]. Similar evidence was found in in vivo models of PD and focal cerebral ischemia after intranigral or intrastriatal injection, respectively [28]. Regarding TRX, which has been described as a cytoplasmic redox-active protein, studies have suggested that it may act as neuroprotective protein in in vitro models of PD and transient focal ischemia, through its antioxidant function upon overexpression and/or administration of human recombinant (hr) TRX [23,29]. CYPA and CYPB are proteins from the immunophilin family of peptidyl-prolyl cis-trans isomerases [30] whose role in CNS remains largely unknown. Nevertheless, they have already shown to be neuroprotective agents against amyloid beta (Aβ)-induced neurotoxicity, suppressing ROS formation [22,25]. Cys C, a cysteine protease inhibitor, was found to be a protector of cortical neurons against hydrogen peroxidase (H2O2)-induced oxidative stress upon exogenous administration of hrCys C [31]. Similar findings were also described for PRDX1, as a cytoplasmic thioredoxin-dependent peroxidase reductase, in which its overexpression in dopaminergic (DA) neuronal cell lines has shown to counteract DA cell death by acting as a ROS (superoxide anion and H2O2) scavenger in a 6-OHDA model of PD [32].
Human SA, which is the most abundant protein in plasma, was found to be a powerful antioxidant mediator, being able to reduce the neuronal death induced by H2O2 or copper/ascorbic acid oxidant stimulation [33]. Finally, HSP27 from the subfamily of small HSPs, which is mostly known by its role in providing thermotolerance to cells [34] and chaperone activity in proteins [35], has demonstrated to be a modulator of ischemic brain damage in vivo, inhibiting the levels of oxidative stress [24,26]. In addition to these functions, we have observed that the relative expression levels of the above-referred proteins were significantly different in the CM of BMSCs, ASCs, and HUCPVCs (Fig. 2). Specifically, CYPB was the only protein upregulated in BMSC CM when compared with HUCPVC (P < 0.05) and ASC (P < 0.01) CMs. CYPA, on the other hand, was just significantly more expressed in BMSC CM when compared with ASC CM (P < 0.05). Additionally, it was also found that DJ-1 was just upregulated in the BMSC CM when compared with HUCPVC CM (P < 0.05). On the other hand, upon comparison of protein profile between HUCPVC and ASC CMs, results showed that TRX and CYPA expression was significantly elevated in HUCPVCs, whereas the opposite was observed regarding Cys C expression (P < 0.05). Taken together, results indicate that BMSC secretome might have a more prone antioxidative profile when compared with HUCPVCs and ASCs. In fact, we have also identified gelsolin, which was only present in the BMSC secretome, and that has been described as a stronger antioxidative molecule in neurodegenerative disorders such as AD [36], thereby supporting the prominent antioxidative properties of BMSCs.
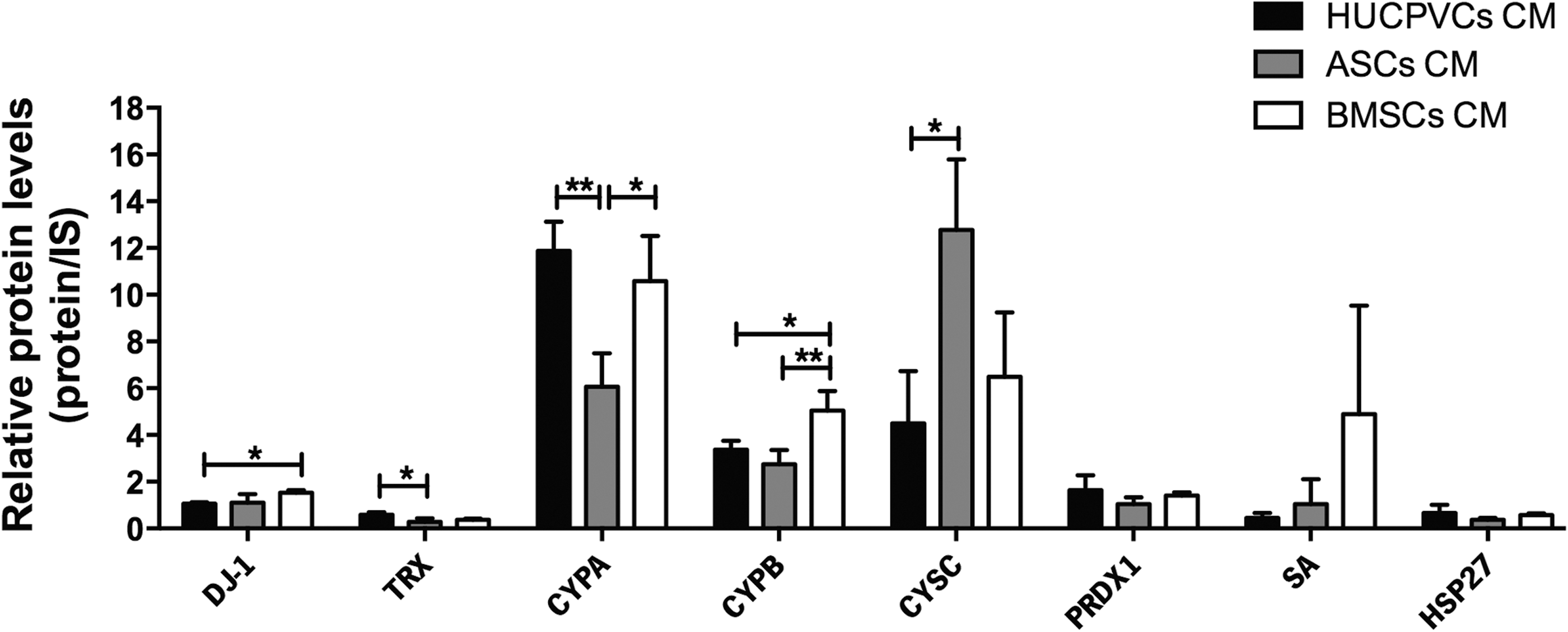
HUCPVC, ASC, and BMSC CM identified proteins with potential antioxidative functions on CNS physiology. Proteomic analysis revealed that CYPB was significantly more expressed in BMSC CM when compared with HUCPVC and ASC CMs. CYPA and CYPB expression was significantly elevated in BMSC CM when compared with the ASC CM. Similarly, comparisons between BMSC CM and HUCPVC CM showed that CYPB and DJ-1 were significantly more expressed in BMSC CM. On the other hand, statistical differences between HUCPVC CM and ASC CM revealed that expression of TRX and CYPA was significantly increased in HUCPVC CM, whereas CYSC was significantly upregulated in ASC CM. Data are expressed as mean ± SD. *P < 0.05, **P < 0.01. CNS, central nervous system; IS, internal standard; SD, standard deviation.
Apoptosis
CYPA, CYPB, Cys C, IL-6, Gal-1, and HSP27 have been described as antiapoptotic proteins [24 –26,31,37,38]. CYPA, CYPB, and Cys C have shown to have a dual role in CNS, playing as antioxidative and antiapoptotic modulators in in vitro models of AD and PD. For instance, the administration of hrCYPA or CYPB revealed to prevent PC12 cell death against Aβ-mediated apoptosis when exposed to aβ peptides [22,25]. On the other hand, Cys C was demonstrated to be a neuroprotective agent against Aβ and 6-OHDA-induced apoptotic neuronal death [31,38,39]. IL-6 has also shown to have a protective role within the CNS. For instance, intracerebral administration of hrIL-6 was demonstrated to be an antiapoptotic agent, able to reduce neuronal cell death in transient and permanent rodent models of cerebral ischemia [40]. Concerning Gal-1, described as an endogenous soluble mammalian lectin, it was already demonstrated that its infusion into the cerebello-medullar cistern led to the reduction of neuronal apoptosis in a rat model of focal cerebral ischemia by inducing the expression of brain-derived neurotrophic factor [21], also known as an antiapoptotic factor [41]. Similar outcomes were also described for HSP27, respectively [24,26].
Concerning the relative expression levels of the above-referred proteins in the CM of BMSCs, ASCs and HUCPVCs, the results revealed that CYPA, CYPB, CYSC, and IL-6 were differently expressed, while Gal-1 and HSP27 were not (P > 0.05; Fig. 3). CYPB was found to be significantly upregulated in BMSC CM (P < 0.05; P < 0.01), whereas IL-6 was highly expressed in HUCPVC CM (P < 0.05). Concerning differences between HUCPVC and ASC CMs, IL-6 and CYPA expression was significantly superior in HUCPVC CM when compared with the ASC CM (P < 0.05), while the opposite was observed regarding Cys C expression (P < 0.05). Therefore, these results indicate that both BMSC and HUCPVC secretomes may exhibit a similar antiapoptotic profile, in which the sole expression of SDF-1α and gelsolin by BMSCs and CSF-1 by HUCPVCs (defined as neuroprotective and antiapoptotic agents [36,42 –44] supported even more the above-referred outcomes.

Specific HUCPVC, ASC, and BMSC CM proteins with potential antiapoptotic effects on CNS physiology. Proteomic analysis revealed that CYPB was significantly upregulated in BMSC CM when compared with HUCPVC CM and ASC CM. In contrast, IL-6 was highly expressed in HUCPVC CM when compared with BMSC and ASC CM. CYSC was found upregulated in ASC CM when compared with HUCPVC CM. In opposition, CYPA was downregulated in ASC CM when compared with both HUCPVC and BMSC CMs. Data are expressed as mean ± SD. *P < 0.05, **P < 0.01. IL-6, interleukin-6; IS, internal standard.
Excitotoxicity
Excitotoxicity represents a pathological process by which neurons are damaged and killed by the overactivation of excitatory neurotransmitter receptors involved in stroke, traumatic brain injury, and neurodegenerative diseases [45]. The IL-6, PEDF, and PAI-1 identified in the proteomic analysis are described as important neuroprotective mediators against glutamate-induced excitotoxicity [46,47]. In addition to its antiapoptotic action, IL-6 has also the ability to reduce neuronal damage induced by glutamate in hippocampal neurons in vitro [48] as well as in striatal cholinergic neurons in vivo [49]. Similarly, PEDF, which has been described as a multifunctional protein and a noninhibitory member of the serine protease inhibitor (SERPIN) gene family, was found to be a neuroprotective protein of cerebellar granule cells and hippocampal neurons against glutamate cytotoxicity in vitro [46]. In addition, in neurodegenerative disorders such as PD, PEDF revealed to be a neuroprotective agent against 6-OHDA-induced excitotoxicity [50]. Finally, PAI-1, also a member of the serine protease inhibitor (Serpin E1) superfamily [51], in the recombinant form, has been described as a protective player against the N-methyl-D-aspartate (NMDA)-induced excitotoxicity in cortical neurons through modulation of NMDA-Ca2+ influx upon exogenous stimulation [47]. In terms of the relative expression levels of the above-referred proteins, the proteomic results revealed that IL-6 and PEDF were differently expressed in HUCPVCs (upregulated, P < 0.05) and ASCs (upregulated, P < 0.01; Fig. 4), which indicates that their secretome can play a role in mediating neuroprotection induced by excitotoxicity phenomena.
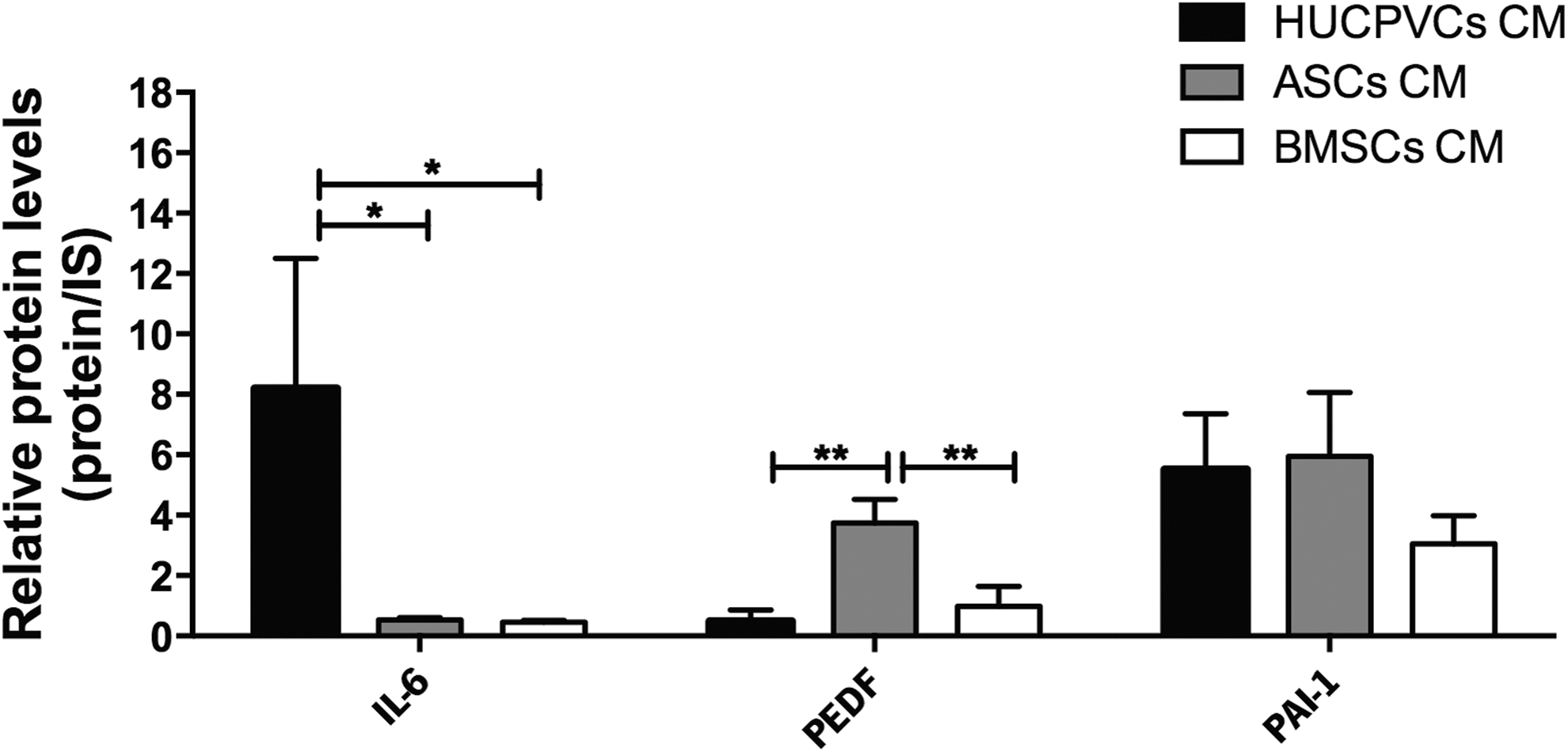
Specific HUCPVC, ASC, and BMSC CM proteins with potential antiexcitotoxicity effects on CNS physiology. Proteomic analysis revealed that IL-6 was significantly more expressed in HUCPVC CM when compared with ASC and BMSC CMs. Results also showed that PEDF expression was significantly elevated in ASC CM when compared with HUCPVC and BMSC CMs. Data are expressed as mean ± SD. *P < 0.05, **P < 0.01. IS, internal standard; PEDF, pigment epithelium-derived factor.
As mentioned above, in addition to the ability of proteins present in hMSC secretome to regulate processes, such as oxidative stress, apoptosis, and glutamate-mediated excitotoxicity, proteomic analysis also revealed the presence of proteins involved in the regulation of proteasomal degradation, neurogenesis, inflammation, glial scarring, and toxic protein deposition (Fig. 5). One of these proteins was UCHL1, which is both a ubiquitin (UB) hydrolase [52] and UB-ligase enzyme mostly localized in neurons [53]. UCHL1 has been shown to be involved in regulation of proteasomal degradation [54], which has been linked with neurodegenerative diseases such as AD and PD [53]. As can be observed in Fig. 5, results show that UCHL1 was significantly upregulated in HUCPVC CM (P < 0.05). For TRX, besides its antioxidant activity, it was recently reported that it is also a promoter of hippocampal neurogenesis, leading to cognitive recovery in a cerebral ischemia model [55]. Like UCHL1, TRX was also significantly increased in HUCPVC CM (P < 0.05).
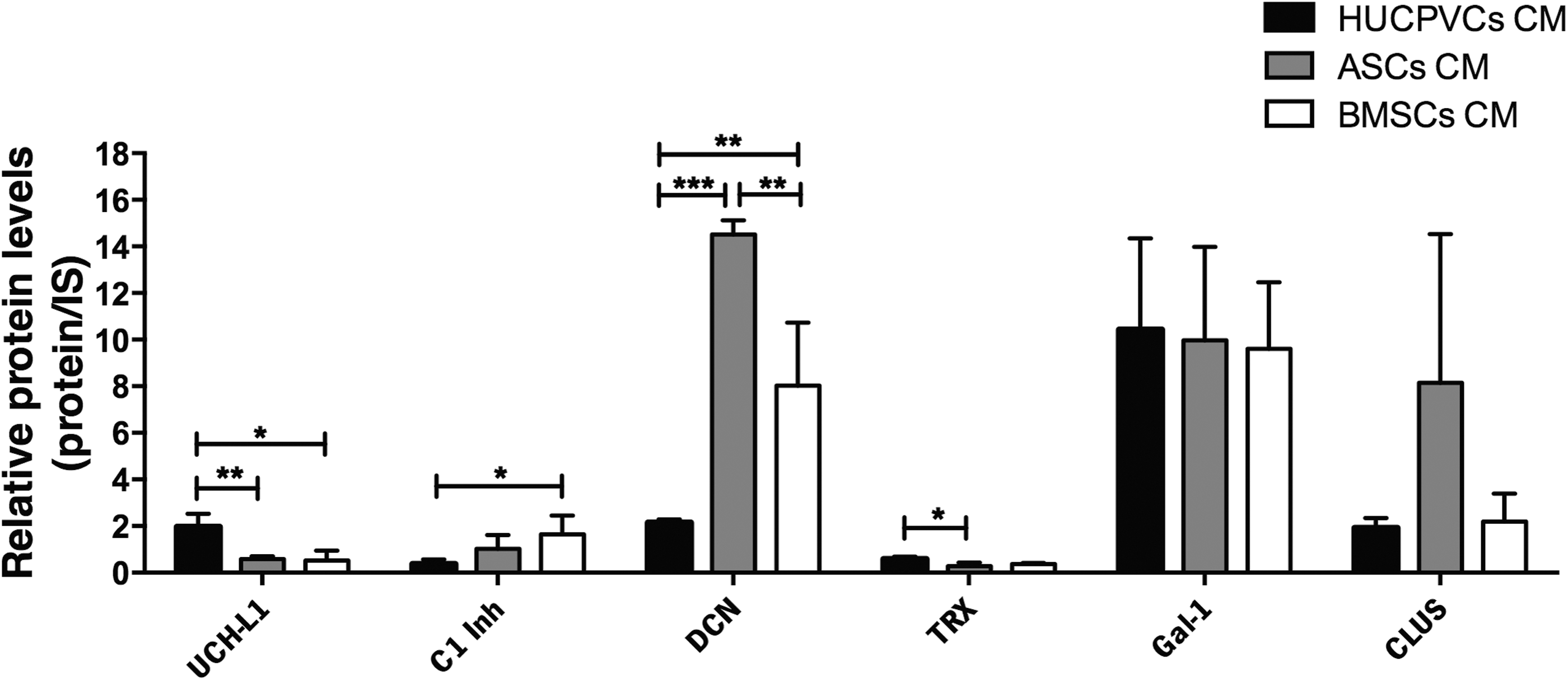
Specific HUCPVC, ASC, and BMSC CM proteins with potential actions in the regulation of proteasome degradation (UCHL1), neurogenesis (TRX, Gal-1), inflammation (c1-Inh), glial scarring (DCN), and toxic protein deposition (CLUS). Proteomic analysis revealed that UCHL1 expression was significantly higher in HUCPVC CM when compared with both ASC and BMSC CMs. TRX expression was also significantly elevated in HUCPVC CM when compared with ASC CM. C1-inh was significantly increased in BMSC CM when compared with HUCPVC CM. On the other hand, results showed that DCN expression was significantly upregulated in ASC CM when compared with both HUCPVC and BMSC CMs and also in BMSC CM when compared with ASC CM. Data are expressed as mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001. IS, internal standard; UCHL1, ubiquitin-carboxy-terminal hydrolase 1.
Another protein that was found in hMSC secretome was plasma protease C1-Inh, a glycoprotein that belongs to the superfamily of serine protease inhibitors (Serpin G1) [56]. At the same time, it is also an endogenous inhibitor of the complement classical pathway and the contact-kinin systems [56]. Actually, the former system is involved in a variety of immune inflammatory responses, whereas the contact-kinin system is involved not only in inflammation but also in coagulation and blood pressure control, both of which have been shown to play crucial roles in the pathophysiology of ischemic stroke [57]. As displayed in Fig. 5, C1-Inh was upregulated in BMSC CM (P < 0.05). The role of this protein in the CNS is still largely unknown, but it would be interesting to evaluate its effects on the modulation of the activity of microglial cells.
Proteomic analysis also revealed the presence of a small leucine proteoglycan protein, named DCN [58], which was the only antiscarring quantifiable molecule found in the analysis of hMSC CM. Indeed, hrDCN administration has been reported to promote axon regeneration, even across the lesions, by acting as an antiscarring agent both in vitro and in vivo [58]. Actually, DCN antiscarring effect was attributed to the reduction of inflammatory fibrosis, astrogliosis, and levels of several scar-related elements (eg, chondroitin sulfate proteoglycans). The expression of this protein was increased in the ASC secretome (P < 0.001) and also in BMSCs relative to HUCPVCs (P < 0.01).
Finally, regarding proteins involved in neuroprotection phenomena, within the hMSC secretome, we have also found CLUS, which is an extracellular chaperone found in all human fluids [59]. In fact, it has been shown that CLUS displays a protective function against Aβ-induced neurotoxicity in vitro [59]. However, although the exact mechanism by which secreted CLUS protects neuronal cells from Aβ neurotoxicity is unclear, it is thought to be related with CLUS-Aβ complex formation, leading to the attenuation of Aβ aggregation and subsequent degradation by lysosomes [59]. In terms of expression, results showed that although a much more noticeable expression of CLUS was found in ASC CM, no significant differences were found when compared with the other hMSC populations. Collectively, results indicate that HUCPVC CM seems to have a more pronounced role in mediating neuroprotective activity associated with abnormal proteasomal degradation, and also in promoting neurogenesis.
Neurite outgrowth and neurodifferentiation
Although the mechanisms by which hMSC secretome is able to modulate the behavior of neural progenitors remain unclear from the molecular point of view, studies have determined that a network of multiple signaling pathways and transcriptional regulators controls the differentiation of neural progenitors [60]. In fact, from our proteomic analysis, we have found that PEDF, CADH2, and IL-6 have been related to processes such as neurite outgrowth and neuron differentiation (Fig. 6) [61 –63]. Besides its role as a neuroprotective factor, PEDF has been described as a mediator of neuronal survival and differentiation. Exogenous addition of PEDF to human Y-79 retinoblastoma cells and embryonic chick spinal cord motor neurons not only promotes survival but also stimulates the formation of a dense meshwork of neurites [62,64]. Similarly, IL-6 has also been demonstrated to induce neuronal differentiation in PC12 cells [61]. On the other hand, CADH2, a neuronal cell adhesion glycoprotein, has been found to induce both morphological and biochemical features of differentiated neurons in embryonic carcinoma P19 cells [63]. Regarding relative expression levels, IL-6 and CADH2 were significantly increased in HUCPVC CM when compared with ASC CM (P < 0.05), whereas the contrary was observed concerning PEDF (P < 0.01). Nevertheless, these data indicate the presence of proteins that promote neurodifferentiation in the secretome of the three hMSC populations, further reinforcing the neurodifferentiation properties of the secretome of these cells. In addition to these findings, we have also found proteins documented to play roles in axon guidance and neurite outgrowth, namely SEM7A [65], GDN [66], and BASP-1 [67] (Fig. 6). SEM7A, which belongs to the semaphorin family of axon guidance proteins, has been described to enhance axon growth from olfactory bulb explants in both its soluble and membrane-bound forms [65]. GDN, a serine protease inhibitor from the serpin family [66], has also been shown to promote neurite outgrowth in neuroblastoma (NB2a) cells, and also in rat hippocampal cells [68]. Finally, BASP-1, a major protein of neuronal lipid rafts, when overexpressed in PC12E2 cells (a subclone of PC12 cells) and rat primary hippocampal neurons, was found to be a stronger stimulator of neurite outgrowth in both cell types [67]. Thus, as can be observed in Fig. 6, HUCPVC CM expression of GDN and SEM7A was significantly downregulated when compared with both BMSC CM (P < 0.01) and ASC CM (P < 0.05). Thus, from the obtained data, BMSC and ASC CMs might exhibit a similar profile in promoting neurite outgrowth when compared with HUCPVCs. In fact, in addition to the above-quantified proteins, we have also identified β4Gal-T (only in the de ASC CM) and SDF-1α (only in BMSC CM), which have been described as promoters and regulators of neurite outgrowth in the CNS [69 –72]. At the same time, despite the (apparently) less prominent profile of HUCPVCs on this topic, we have also identified important molecules (only in their CM) playing a role in the regulation of neural progenitor cell maintenance and maturation as Cyr61 and CSF-1, respectively [73,74].
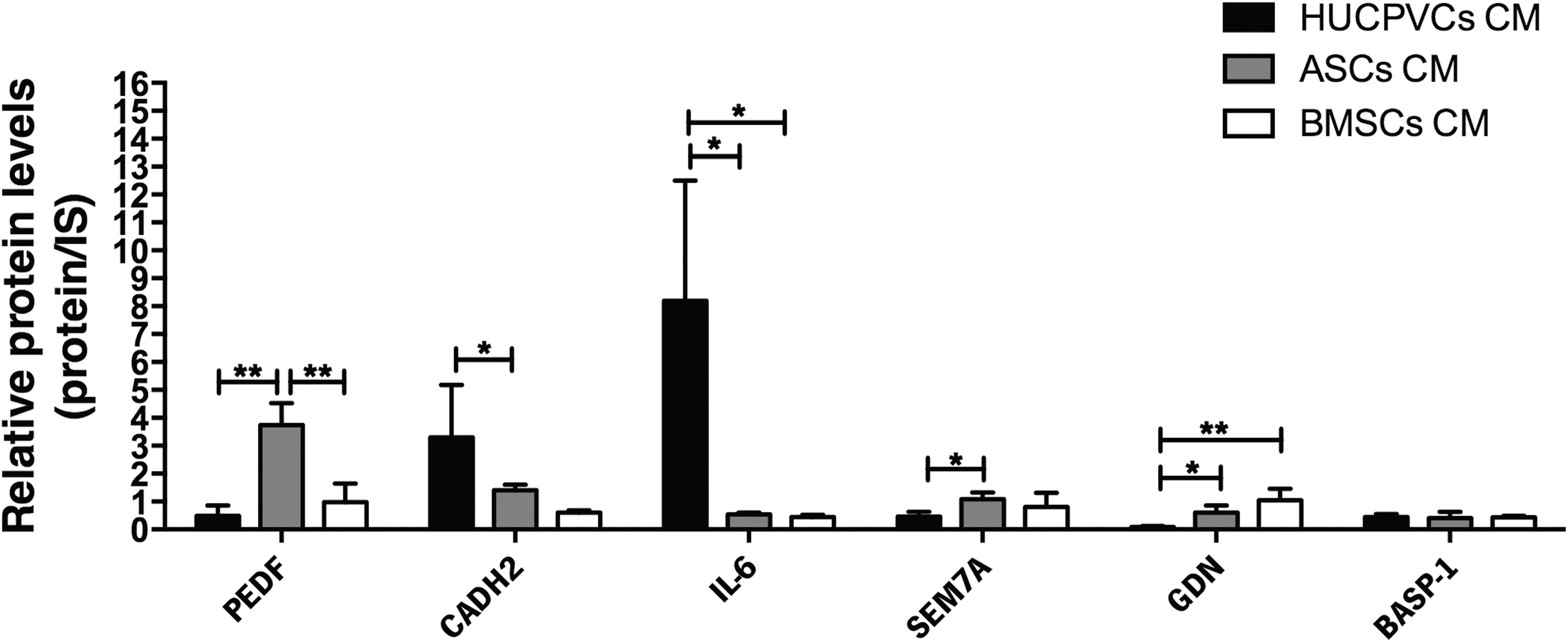
Specific HUCPVC, ASC, and BMSC CM proteins with potential effects in neurite outgrowth and neuronal differentiation. From the comparative analysis of the secreted proteins in the different MSC CM populations, we have identified proteins involved in neurodifferentiation, namely PEDF, CADH2, and IL-6. IL-6 and PEDF were significantly upregulated in HUCPVC CM and ASC CM, respectively, when compared with the other CM groups. CADH2 were significantly increased in HUCPVC CM when compared with ASC CM. In contrast, PEDF was found to be significantly downregulated in HUCPVC CM and BMSC CM when compared with ASC CM. On the other hand, LC-MS/MS analysis also revealed that proteins involved in neurite outgrowth (SEM7A, GDN, and BASP-1) and GDN expression were significantly elevated in BMSC CM when compared with both HUCPVC and ASC CMs. HUCPVC CM exhibited significantly lower expression of SEM7A and GDN when compared with both ASC and BMSC CMs. Data are expressed as mean ± SD. *P < 0.05, **P < 0.01. GDN, glia-derived nexin; IS, internal standard.
Conclusions
In the present work, we have characterized the secretome of BMSCs, ASCs, and HUCPVCs through proteomic-based analysis, identifying and quantifying the relative expression differences among these hMSC populations. Our results showed that hMSCs derived from different tissues may have distinct profiles, which could lead to different action(s) against distinct physiological or pathogenic processes involved in CNS disorders/injuries. Indeed, the evaluation of the hMSC CM secretion profile, based on the differential expression of proteins with neuroprotection character, indicated that BMSC CM might be the most advantageous choice for a therapy designed to reduce oxidative stress, while HUCPVCs and ASCs could more beneficial in the protection against excitotoxicity. On the other hand, results also suggest that HUCPVCs may be less appropriate for antiscarring phenomena, whereas it might be the most indicated for targeting abnormal proteasomal degradation. Likewise, both BMSC and HUCPVC CMs might be advantageous as antiapoptotic agents. Therefore, future studies should focus on validating the efficiency of the CM of each individual hMSC source for each of the aforementioned biological processes.
Footnotes
Acknowledgments
The authors are grateful to the Foundation Calouste de Gulbenkian for the funds attributed to A.J.S.; Portuguese Foundation for Science and Technology (FCT) for PhD fellowships attributed to A.O.P. (SFRH/BD/33900/2009) and S.I.A. (SFRH/BD/81495/2011) and Ciência 2007, IF Development Grant attributed to A.J.S., and projects PTDC/NEU-NMC/0205/2012, UID/NEU/04539/2013; cofinanced by COMPETE Programa Operacional Factores de Competitividade; and by the National Mass Spectrometry Network (RNEM) (REDE/1506/REM/2005); Prémios Santa Casa Neurociências—Prize Melo e Castro for Spinal Cord Injury Research; cofunded by Programa Operacional Regional do Norte (ON.2–O Novo Norte), ao abrigo do Quadro de Referência Estratégico Nacional (QREN), and através do Fundo Europeu de Desenvolvimento Regional (FEDER). The authors also would like to thank Professor J.E.D. (University of Toronto, Canada) and Professor J.M.G. (Tulane University) for kindly providing HUCPVCs and ASCs, respectively.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.