
Abstract
An increasing number of studies have detected mesenchymal stromal cells (MSCs) and mesenchymal progenitor cells (MPCs) in the peripheral blood (PB). This study aimed to systematically review the possibility of using the PB as a source for chondrogenic progenitors. PubMed, the Web of Science, and Embase were searched for relevant articles. The findings of the studies were reviewed to evaluate the biological characteristics of PB-derived MSCs, chondrogenic MPCs, and their applications in cartilage repair. Thirty-six articles were included in the final analysis, 29 of which indicated that PB is a potential source for chondrogenic progenitor cells. Thirty-two studies reporting in vitro data, including 79.2% (19/24) of studies on PB MSCs and 75% (6/8) of studies on chondrogenic PB MPCs, confirmed the existence of PB MSCs and PB MPCs, respectively; all in vivo investigations showed that using PB as a cell source enhanced cartilage repair. PB MSCs were found in most of the animal studies (12/13), whereas 7 of 11 human studies described the presence of PB MSCs. This systematic review strongly indicates the existence of MSCs in the PB of animals, whereas the presence of MSCs in human PB is less clear. Although the presence of both MSCs and chondrogenic MPCs in the PB, as well as a few favorable outcomes associated with the use of PB-derived progenitors for cartilage repair in vivo, suggests that the PB is a potential alternative source of chondrogenic progenitor cells for cartilage repair, the efficacy of these cells has not been compared to those from other sources, such as bone marrow or adipose tissue in controlled studies.
Introduction
A
Mesenchymal progenitor cells (MPCs) from a mesoderm origin contain appealing adult stem cells, whose multipotency and low immunogenicity suggest that these cells may be broadly used in the field of cell-based tissue regeneration [1]. Traditionally, bone marrow (BM) has been regarded as the richest reserve of MSCs, and BM MSCs are the most widely used cells in regenerative medicine. MSCs have also been isolated from a variety of other tissues, such as adipose tissue, trabecular bone, cartilage, cord blood (CB), the umbilical cord, the amniotic membrane and fluid, and cervical cancer cells [2 –7].
Since Fernandez et al. [8] identified MSCs in mobilized peripheral blood (PB) from breast cancer patients, the use of the PB instead of the BM as an alternative source of cells for tissue engineering has been extensively explored due to the minimally invasive harvesting procedures and large quantities of sample. Some researchers have successfully identified MSCs in the PB [9 –25], and others have reported PB progenitor cells with MSC-like multiple differentiation potential [26 –29] or a capacity to differentiate into only one or two mesenchymal lineages [30 –37]. In contrast, several research groups have challenged the presence of circulating multipotent progenitors with contradicting evidence [38 –42].
Despite the controversy regarding the existence of MSCs in the PB, some studies have achieved satisfactory outcomes using PB as a source of chondrogenic cells for animal and even human cartilage repair [14,25,43 –47]. These encouraging results indicate that the PB might have potential applications in cartilage regeneration because it contains MSCs or chondrogenic mesenchymal progenitor cells (MPCs), and blood sample collection is easy and minimally invasive in clinical settings.
To date, systematic reviews describing MSCs in the PB have been lacking, and the chondrogenic potential of the PB-derived progenitor cells remains debatable. This review aims to assess the chondrogenic potential of progenitor cells derived from PB and conclusively determine whether the PB contains MSCs.
To better understand the current literature in this field and to avoid ambiguity and misunderstanding, the scientifically accurate and standardized term “multipotent mesenchymal stromal cell” proposed by the International Society for Cellular Therapy (ISCT) [48] will be used instead of the currently commonly used term “mesenchymal stem cell.” Although chondrogenic PB MPCs, as reported in some publications, did not meet the strict ISCT standard of MSCs due to a partial mesenchymal-lineage differentiation potential or the lack of immunophenotyping data, their chondrogenic differentiation capacity suggested that these cells could be a potential cell source for cartilage repair.
In the current review, we critically evaluated published data on the presence of MSCs and MPCs in the PB, with a focus on the application of PB-derived MSCs or chondrogenic MPCs in cartilage repair. The term PB MSCs was adopted to specifically describe human PB-derived multipotent primitive cells that meet the minimal criteria proposed by the ISCT [48]. Because the surface markers for MSCs from other species have not been universally characterized, the term PB MSCs was also assigned to nonhuman PB-derived, plastic-adherent, and fibroblast-like cells that can differentiate into mesenchymal trilineages and express nonhemopoietic immunophenotypes, whereas the label chondrogenic PB MPCs represented PB-derived primitive cells that are chondrogenic with or without a capacity to differentiate into other lineages but do not meet the ISCT MSC standard or lack immunophenotyping data.
Materials and Methods
We conducted a systematic review of the available English language literature according to the PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-analyses) standards and a PRISMA checklist. The initial citation search terms were “mesenchymal AND circulating,” “ cartilage AND blood AND mesenchymal,” “cartilage AND blood AND peripheral,” or “mesenchymal AND blood AND peripheral.” We performed the search in PubMed, EMBASE, and the Web of Science on September 1, 2015.
Meeting and conference abstracts were excluded. All references within the included studies were searched for articles of interest that had not been identified by the initial search criteria. Any basic English language scientific studies of PB-derived primitive cells that exhibited chondrogenic or multipotent mesenchymal differentiation abilities were included. Furthermore, in vivo animal or human studies using the PB as a source of chondrogenic progenitor cells for cartilage regeneration were also included. Any studies of primitive cells that were not chondrogenic or not derived from the PB and in vivo studies that only used non-PB sources were excluded.
Results
Study characteristics
The literature search retrieved 855 studies (Fig. 1). Five hundred twenty studies were reviewed further for eligibility after excluding 75 non-English studies and 260 reviews and proceedings. Furthermore, 475 duplicate or unrelated studies and 9 in vitro studies in which a chondrogenic induction assay was not performed were excluded. This strategy produced 36 articles published between 1997 and 2015 (Table 1). The final data from the 36 studies were analyzed, including 29 in vitro studies, 3 studies involving both in vivo and in vitro experiments, and 7 in vivo studies (Tables 2 –4). The experimental subjects included humans, small animals (ie, mouse, rat, and rabbit), and large animals (sheep, pig, and horse).
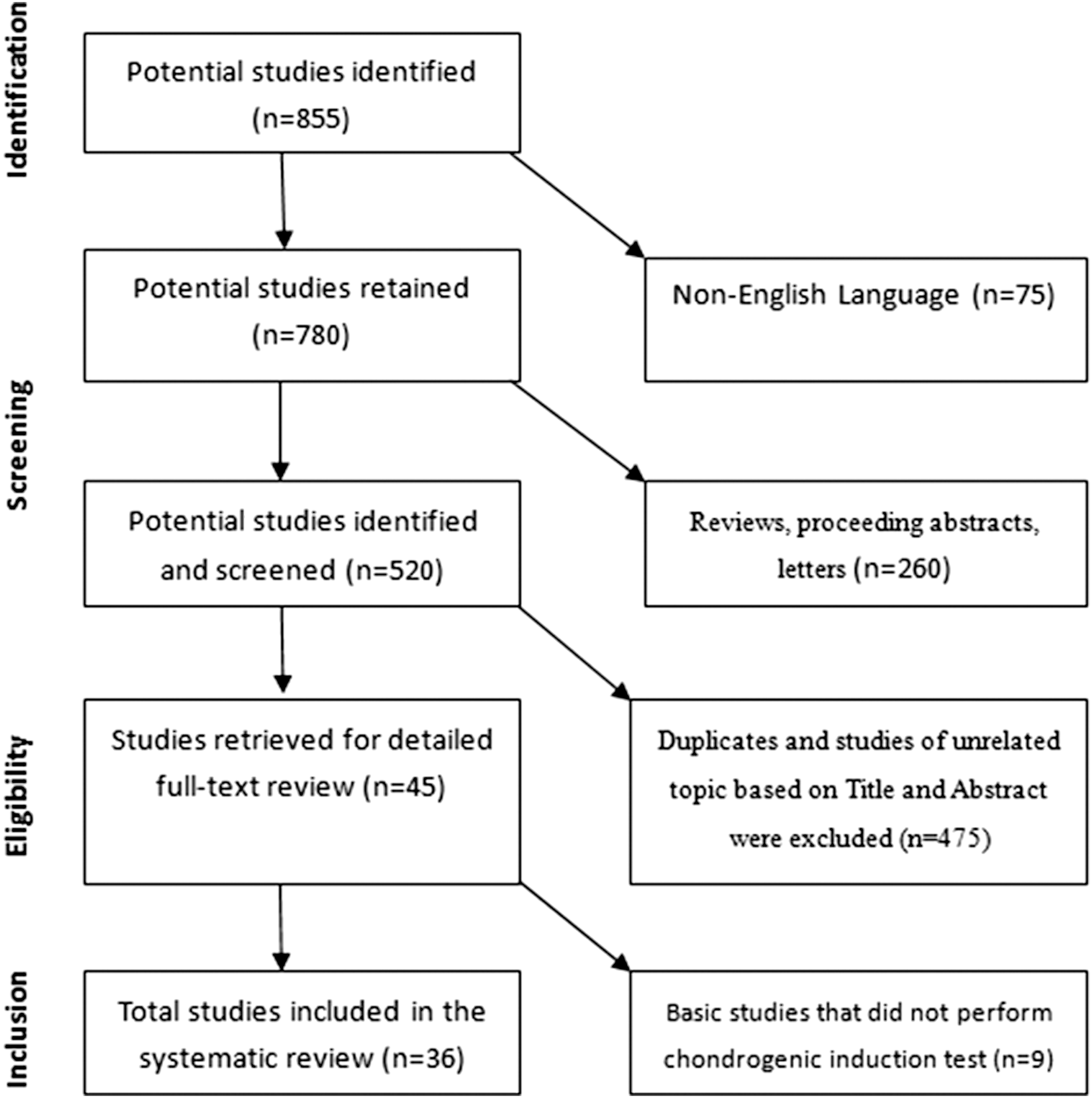
Flowchart of the literature search.
MPC, mesenchymal progenitor cells; N/A, not available; PB, peripheral blood; PB MSC, peripheral blood mesenchymal stromal cells.
CFU, colony forming units; DGC, density gradient centrifugation; G-CSF, granulocyte colony-stimulating factor; MMC, mitomycin C; MSC, mesenchymal stromal cells; N, no; N/A, not available; PA, plastic adherence; PB, peripheral blood; PBMC, peripheral blood mononuclear cells; PB MSC, peripheral blood mesenchymal stromal cells; RBC, red blood cells; Y, yes.
DGC, density gradient centrifugation; G-CSF, granulocyte colony-stimulating factor; MSC, mesenchymal stromal cells; N, no; N/A, not available; PA, plastic adherence; PB, peripheral blood; PB MSC, peripheral blood mesenchymal stromal cells; PBMCs, peripheral blood mononuclear cells; RBC, red blood cells; Y, yes.
ACI, autologous chondrocyte implantation; APT, autologous periosteum flap transplantation; BM, bone marrow; BMS, bone marrow stimulus; DGC, density gradient centrifugation; ECMO, extracorporeal membrane oxygenation; G-CSF, granulocyte colony-stimulating factor; HA, hyaluronic acid; ICRS, International Cartilage Repair Society; IKDC, International Knee Documentation Committee; KOOS, knee injury and osteoarthritis outcome score; MSC, mesenchymal stromal cells; N, no; N/A, not available; PA, plastic adherence; PB, peripheral blood; PB MSC, peripheral blood mesenchymal stromal cells; PBMCs, peripheral blood mononuclear cells; PBNC, peripheral blood nucleated cells; PBPC, peripheral blood progenitor cells; PF, patellofemoral; RCT, randomized controlled trial; Y, yes.
Of the 32 studies with in vitro evidence, PB MSCs were successfully isolated and validated in 79.2% (19/24) of the studies, whereas chondrogenic PB MPCs were recovered in 75% (6/8) of the related studies. The remaining seven in vitro studies either failed to identify PB MSCs (4) or PB chondrogenic MPCs (2) or concluded that the PB was not a viable source of MSCs (1). In addition, seven in vivo studies evaluated the outcomes of the use of PB-derived cellular components for cartilage regeneration, including four case reports (seven in patients and one in horses), two controlled animal studies, and only one random controlled trial (RCT).
MSCs in human PB
Conflicting studies on the presence of MSCs in human PB have been reported. Eleven studies investigated the presence of MSCs in human PB, seven of which confirmed the existence of PB MSCs, whereas the other four failed to identify MSCs in the PB or concluded PB that was not a viable source of MSCs (Table 2).
Various isolation and cell culture methods have been adopted to improve the putative MSC yields. Tondreau et al. [49] claimed that human MSCs could be easily isolated from the PB using three methods: plastic adhesion, plastic adhesion supplemented with 5% BM-MSC conditioned medium, and the selection of CD133-positive cells using magnetic beads coated with an anti-CD133 antibody. The putative PB MSCs were confirmed based on the expression of MSC surface markers and the capacity to differentiate into adipocytes, osteocytes, chondrocytes, and even neuronal/glial cells.
In addition, using fibrin microbead-based isolation, Kassis et al. [22] also separated a significant number of human MSCs from peripheral blood progenitor cells (PBPCs) by mobilizing the PB of healthy adults with a granulocyte colony-stimulating factor (G-CSF). Hopper et al. [25] isolated peripheral blood mononuclear cells (PBMCs) from healthy human donors; the adherent PBMC population displayed a fibroblast-like morphology after 2 weeks of hypoxic culture, and 94% of the population expressed MSC markers (Stro-1, CD90, CD106, CD105, CD146, CD166, and CD44), whereas only 41% of cells expressed these markers under normoxic conditions.
Traditional methods, such as density gradient centrifugation and adherence to plastic, have been proven to efficiently isolate PB MSCs. Kim et al. [23] isolated PB MSCs and confirmed that the obtained cells are morphologically and immunophenotypically similar to BM MSCs. Chong et al. [21] recovered MSCs from nonmobilized human PB, and the PB-derived MSCs possessed trilineage differentiation potential and expressed CD105, CD166, and CD29, but did not express CD34 and CD45. Moreover, these cells exhibited similar characteristics and a similar ability to undergo chondrogenic differentiation as BM-derived MSCs.
Some studies have adopted pathologic conditions to mobilize mesenchymal progenitors in the circulation. Alm et al. [19] reported the isolation of circulating plastic-adherent MSCs from patients with hip fractures, but not from women with hip osteoarthritis. Similarly, Bui et al. [20] also detected MSCs in blood samples from 18 of 58 patients with extracorporeal membrane oxygenation. More than 95% of the MSCs displayed MSC phenotypes after Passage 3 and could be expanded for 3 to 17 population doublings. Notably, none of the six normal blood samples produced MSCs.
However, some studies have failed to detect viable progenitors in either normal or mobilized PB. In an early study conducted in 1997, Lazarus et al. [38] failed to isolate MSCs in the PBPCs collected by leukapheresis from human G-CSF-mobilized blood. Likewise, Wexler et al. [40] evaluated the frequency, phenotype, and differentiation potential of MSCs in the peripheral blood stem cell (PBSC) population of a normal donor, adult BM, and CB. Cultured cells derived from CB or PBSCs did not survive beyond Passage 1. Both the CB and PBSC cultures only produced a population of loosely adherent, nonconfluent, sparse macrophage-like cells with a hemopoietic phenotype (CD45+, CD14+).
In 2008, Lund et al. [39] succeeded in obtaining cells with a fibroblastic morphology from 3 of 6 G-CSF-mobilized human PB samples. The isolated cells exhibited an immunophenotyping profile that was similar to that of BM MSCs and able to differentiate into osteogenic and adipogenic lineages. Nevertheless, these cells possessed minimal chondrogenesis and limited expansion potential and became senescent 20 to 25 days after isolation. Furthermore, the PB-derived clone-forming cells did not exhibit telomerase activity and displayed evident telomere shortening. Recently, Hoogduijn et al. reported that MPCs or MSCs were not observed in the PB of normal individuals or severely ill patients [42].
MSCs in animal PB
The presence of PB MSCs in animals has been extensively investigated in 13 studies (Table 2). Ahern et al. [41] failed to isolate MSCs from equine PB apheresis. PBMCs successfully adhered to culture plates in only one of six samples, but did not proliferate. In contrast, the remaining 12 studies validated the existence of MSCs in the PB of both large and small animals, such as horses [9 –11,50], pigs [51], sheep [17], rabbits [14,15], rats [13,18,24], and mice [24].
Most equine studies support the existence of circulating MSCs, except for the report by Ahern et al. [41]. De Schauwer et al. [10] and Spaas et al. [50], who are from the same research group, successfully isolated putative MSCs from equine PB. These authors were the first to perform detailed immunophenotyping and functional characterization of MSCs derived from the PB of horses. Likewise, in a study by Bussche and Van de Walle [9], the MSCs isolated from the PB of horses expressed typical MSC surface markers and were able to differentiate into the trilineage. Moreover, Dhar et al. [11] isolated MSCs from the PB of 3 of 10 horses without hyperbaric oxygen treatment. Interestingly, after three consecutive hyperbaric oxygen treatments, MSCs could be isolated from three of the remaining seven horses that had not yielded MSCs.
In addition to horses, PB MSCs have been found in other large animals, such as sheep and pigs. Lyahyai et al. [17] first characterized ovine PB MSCs by confirming their osteogenic, chondrogenic, and adipogenic differentiation and transdifferentiation to neuronal-like cells. An RT-PCR analysis of the surface markers showed that five of six CD34-positive cultures were also positive for CD29, CD73, and CD90, but negative for CD45. Casado et al. [51] also successfully isolated MSCs from porcine PB with trilineage differentiation capacity and a human MSC phenotype.
Notably, PB MSCs were often present in the mobilized PB of small mammals (rabbits and rats). Fu et al. [12,14,15] combined G-CSF and AMD3100 to mobilize the PB of adult New Zealand white rabbits. The isolated progenitors were confirmed to be MSCs based on their morphological characteristics, surface markers, and mesenchymal lineage differentiation potential. Another study by Fu et al. [13] suggested that the mobilized PB may be an alternative source for MSCs due to the similar biological characteristics of MSCs obtained from the BM and PB of rats, including their mesenchymal trilineage differentiation capacity, apoptosis resistance, and cellular senescence. Furthermore, PB MSCs appeared more chondrogenic in vitro than BM MSCs, as evidenced by their more extensive collagen type II immunostaining.
In another rat study, Rochefort et al. [18] regularly observed circulating MSCs with immunophenotypes and trilineage differentiation potential similar to those of BM MSCs. The circulating MSC pool was consistently and dramatically increased (by ∼15-fold) when the rats were subjected to chronic hypoxia. The hypoxia-induced mobilization appeared specific to MSCs because the population of total hematopoietic progenitor cells did not increase significantly.
In a study of the mechanisms of PB MSC mobilization, Liu et al. [16] were able to mobilize MSCs in the PB of rats exposed to short-term hypoxia (2 days), and the mobilization efficiency increased in a time-dependent manner (2–14 days). The cultured, adherent PB-derived cells were positive for CD90, CD29, and CD44 but negative for CD34 and CD45; the induction of differentiation showed their mesenchymal differentiation potential. Recently, Li et al. [24] isolated a CD45-negative subset of the resulting fibroblast cells, which were called blood-derived MSCs due to their expression of CD105, CD73, and CD90, lack of CD45, CD34, CD11b, and CD19 expression, and mesenchymal trilineage differentiation capacity.
PB-derived chondrogenic MPCs in human and animals
Eight studies focused on the chondrogenic ability of PB-derived progenitor cells (Table 3), including four human and four animal studies. Specifically, three human and three animal studies claimed to isolate chondrogenic MPCs, whereas neither Kuwana et al. [37] nor Koerner et al. [36] detected chondrogenesis of PB MPCs from the PB of humans and horses, respectively.
Although these studies substantiated that the circulating adherent and fibroblast-like cells exhibited chondrogenic or even trilineage differentiation capacity, these cells could only be identified as chondrogenic PB MPCs, not PB MSCs, due to either partial potential toward mesenchymal differentiation or a lack of mesenchymal immunophenotyping validation, which left the identity of the progenitors in doubt [26 –29,32,33].
For instance, Proffen et al. [27] and Wan et al. [29] isolated MSCs in the PB of pigs and rabbits, respectively, but merely substantiated that these cell types had the ability to undergo osteogenic, adipogenic, and chondrogenic differentiation. Although Trivanovic et al. [28] identified fibroblast-like and clonogenic cells in human PB that exhibited multilineage (osteogenic, chondrogenic, adipogenic, and myogenic) differentiation, the atypical MSC immunophenotypes (40% CD34-positive) called the MSC identity into question. Raghunath et al. [32] cultured PB MPCs from PBMCs obtained from 10 healthy adults. Flow cytometry detected the surface markers CD14 and CD105 on the MPCs, but not CD34, CD45, or CD133. With regard to differentiation, only chondrogenesis was investigated and confirmed.
In a study using cancellous bone scaffolds seeded with G-CSF-mobilized PB-derived stem cells for cartilage regeneration, Turajane et al. found increased expression of cartilage-relevant genes and proteoglycan and glycosaminoglycan contents [33]. Unfortunately, the authors did not report their cell isolation method or osteogenic or adipogenic differentiation capacity, other than chondrogenesis.
In vivo application of PB in cartilage repair
There were seven studies with in vivo data that used PB as a source of progenitors for cartilage repair and achieved promising outcomes (Table 4). Four of the seven studies using autologous PB-derived progenitors for clinical treatment included three human studies and one veterinary patient (horse) study. The other three studies reported successful cartilage regeneration in sheep and rabbits using xenogeneic (human) and allogeneic PB, respectively. Spaas et al. [50] treated a 5-year-old horse with degenerative disease of the pastern joint with autologous MSCs that had been isolated from horse PB. Specifically, 2.5 × 106 cells were injected into the pastern joint, and visual gait evaluation and an objective pressure plate analysis confirmed the positive response to this stem cell-based treatment.
Based on this inspiring result, Fu et al. [14] investigated the feasibility of using PB MSCs for cartilage repair in a rabbit model. The authors successfully isolated and expanded MSCs from PB mobilized by G-CSF and the CXCR4 antagonist AMD3100. Both the allogeneic PB and BM MSCs were compounded with decalcified bone matrix and showed the same ability to repair cartilage defects in vivo, which was in agreement with a recent study by Hopper et al. [25], who found that human PB-derived MSC therapy was as effective as BM MSC therapy for treating osteochondral defects in sheep.
The other four studies intra-articularly applied a PB-derived product to treat patients with ICRS (International Cartilage Repair Society) grade III and IV lesions. Skowronski et al. [46] assessed the long-term clinical outcomes of 52 patients with ICRS grade III or IV lesions treated with microfracture in combination with autologous “blood stem cells” transplant and then covered by a collagen membrane, a procedure similar to autologous chondrocyte implantation. Approximately 90% of the cases achieved good results.
A recent case report by Fu et al. [43] showed the results of a treatment using PB MSCs for human cartilage repair. MSCs derived from mobilized PB were obtained from a 19-year-old male kickboxer with ICRS grade IV chondral lesions. The cartilage defect was addressed with an injection of autologous PB MSCs underneath a transplanted periosteum flap combined with patellofemoral realignment. Second-look arthroscopy showed a smooth surface 8 months after the surgery. After 7.5 years of follow-up, the patient had returned to competitive kickboxing. The IKDC 2000 subjective score, Lysholm and Tegner score, and CT and MRI evaluations showed a significant improvement compared with the presurgery values.
Saw et al. [44] reported articular cartilage regeneration in five patients treated with arthroscopic subchondral drilling and weekly postoperative intra-articular injections of autologous PBPCs in combination with hyaluronic acid (HA). Autologous PBPCs were collected by apheresis after G-CSF mobilization. Flow cytometry showed that most of the cells were CD105 positive. At an average of 17.6 months after the initial surgery, biopsies taken during a second-look arthroscopy confirmed cartilage regeneration and the histological sections exhibited features of hyaline cartilage. However, the outcome of the treatment may be due to the BM stimulus itself, instead of the PBPCs.
In addition, Saw et al. [45] conducted an RCT, in which 50 patients aged 18 to 50 years with ICRS grade III and IV knee cartilage lesions underwent arthroscopic subchondral drilling to further confirm the effects of PBPCs on cartilage repair. Twenty-five patients each were classified into the control (HA) and the intervention (PBSC+HA) groups. Both groups received five weekly injections 1 week after surgery. Three additional injections of either HA or PBSC+HA were given at weekly intervals 6 months after surgery. The second-look arthroscopy, biopsy, and MRI results of 16 patients from each group at 18 months showed that cartilage repair was better in the group that had received PBSCs and HA than in the group that had not received PBSCs. However, the IKDC scores were similar in each group.
Discussion
Based on the reported human and animal studies of the presence of PB MSCs, this systematic review suggested that MSCs are more often found in animals than in humans. A majority of the in vitro studies successfully isolated PB-derived MSCs (12/13) in the PB of animals. Concerning PB MSCs in humans, the current systematic review found that 7 of 11 related studies confirmed the existence of MSCs in human PB (Table 2). In addition, as shown in Table 3, chondrogenic MPCs were recovered in both human (3/4) and animal (3/4) studies, which indicated the presence of chondrogenic MPCs in the PB of both humans and animals. Because most in vitro studies reported the existence of PB MSCs or chondrogenic MPCs, and PB-derived cellular components have been used for cartilage repair in vivo with excellent outcomes, the current review concludes that the PB represents an alternative source for chondrogenic progenitors.
Several studies have been performed on humans and animals to isolate MSCs from the PB, but with conflicting results. Notably, MSCs were more frequently found in the PB from animals, rodents in particular, than in the PB from humans.
The discrepancy between the results of the majority of animal and human studies may significantly depend on the different biology of MSCs between experimental animals and humans. Many of the antigens known to be expressed on human MSCs, such as CD31, CD34, and CD45, are not expressed on MSCs from other species [52,53]. The surface molecules on MSCs are associated with their potential functions. For example, CD105 is a transforming growth factor (TGF)-beta receptor III that potentially plays a role in TGF-beta signaling during chondrogenic differentiation of MSCs [54]. The differential expression of MSC surface markers between humans and other species will certainly affect the marker-associated biology of MSCs, such as the capability to adhere to culture plates or multipotentiality.
Why are the PB progenitors more frequently found in animals than in humans? One possible reason might be that experimental animals are more homogeneous subjects and can be repeatedly tested under more extreme and controlled conditions (hypoxia and hyperbaric treatment) than humans [42].
Although MSCs have been detected in healthy nonmobilized subjects, mobilizing factors, such as growth factors (G-CSF) and pathological conditions (hypoxic or hyperbaric oxygen treatment), have been proven to increase the efficiency of isolating primitive cells from PB. The chemokines and receptors involved in MSC migration from and into the blood have been well studied [55]. AMD3100 is a bicyclam molecule that selectively and reversibly antagonizes the binding of stromal cell-derived factor-1 (SDF-1) to its receptor CXC motif receptor-4 (CXCR4), which results in the egress of hematopoietic stem cells to the PB [56]. Inspired by the fact that MSCs can be mobilized from the BM into the bloodstream, Fu et al. recovered MSCs from G-CSF- and AMD3100-mobilized PB in a series of studies.
However, some studies have failed to detect PB MSCs, even after administering mobilizing agents (G-CSF) or using samples from patients with end-stage organ injury [39,42]. The recent report by Hoogduijn et al. demonstrated that PB MSCs could not be detected in patients with chronic organ failure, but could be found in patients with multiple fractures [42].
Nevertheless, this study does not disprove the concept that MSCs can be mobilized and circulate, because the timing of blood harvesting in patients with chronic diseases had passed the acute phase when the MSCs were most responsive to recruiting signals. In addition, it was observed that rapid sample processing (within 4 h of collection) was a critical sample parameter that produced PB MSCs [20]; a delay in sample processing has been suggested as a contributing factor to the lack of MSC adherence [28,41].
As shown in Tables 2 and 3, MSC isolation from mobilized PB is as successful as isolation from nonmobilized PB in both human (4/7 vs. 3/6) and animal (6/6 vs. 6/7) studies. However, in addition to blood mobilization, several other factors also affect the outcomes of MSC recovery, such as isolation protocols, culture conditions, and culture media, all of which have been shown to significantly impact the developmental potential of the putative MSCs [57].
It should be noted that some of the earlier studies were performed 12 to 20 years ago. For example, the studies from Lazarus et al. [38] and Wexler et al. [40] found no evidence of PB MSCs or only isolated PB progenitors with nonconfluent fibroblast-like cells with hemopoietic phenotypes CD45 and CD14.
With the evolution of cell enrichment techniques, culture media, and the optimization of protocols in recent decades, it is reasonable to assume that these advances have contributed to the recent success in the isolation of PB MSCs or chondrogenic MPCs. For instance, Li et al. recently cocultured nucleated cells from mouse PB with mitomycin C-treated AML12 cells and identified a group of CD45-negative cells with trilineage differentiation capacity and immunophenotypes [24]. Similarly, Hopper et al. found a novel population of adherent cells with MSC phenotypes after culturing human PBMCs in an environment mimicking the oxygen tension of hypoxic tissues [25].
In addition, a variety of methods used to isolate and expand MSCs from the BM have been adopted to improve the recovery of MSCs from PB [58,59]. For instance, MPCs have been successfully enriched using immunophenotypic sorting and MACS [37,49]. A panel of surface markers has been recommended for MSC identification; nonetheless, there are currently no specific surface markers of MSCs. For example, although CD14 is a hemopoietic surface marker, it has been reported that CD14-positive monocytes from healthy human blood exhibited mesenchymal differentiation capacity [37]. It has also been suggested that surface marker-based immunoselection increases the risk of obtaining a subpopulation of cells with specific biological properties [58]. A population of CD133-positive PB MNCs has been shown to differentiate into neural/glial-like cells [49].
Despite the innovations and modifications of isolation protocols, conventional methods (ie, density gradient centrifugation and adherence to plastic) have often been used to efficiently isolate PB MSCs. It remains difficult to compare the protocols from various laboratories and find the most fruitful method for obtaining PB-derived MSCs due to the significant variance in the reported protocols and the scarcity of MSCs in PB.
Most of the currently used culture or differentiation induction assays were based on researchers’ experience with BM MSCs and might not be suitable for progenitor cells from other sources. For instance, many studies have shown that PB MSCs or PB MPCs shared the same ability to differentiate along chondrogenic lineage as BM MSCs. Koerner et al. [36] and Lund et al. [39] showed that the PB-derived progenitor cells did not exhibit chondrogenic activity. After modifying Koerner's culture and differentiation conditions [36], Giovannini et al. [26] confirmed that PB MPCs had mesenchymal trilineage differentiation capacities.
To some extent, the heterogeneous protocols used by various laboratories for MSC isolation, culture, and induction have hampered progress in the search for PB-derived MSCs and have hindered further investigation and application of these progenitor cells [60].
CD34 is considered a well-accepted negative marker for MSCs. It is not surprising that doubt has arisen about whether the PBPCs used by Saw et al. [44,45] for cartilage repair were pure MSCs, as the cellular component is a mixture of both CD105 (mesenchymal marker)-positive and CD34 (hematopoietic marker)-positive progenitors, and the majority of cells were CD105 positive. Therefore, the clinical outcome could not be solely ascribed to the MSCs in the PBPCs.
Interestingly, several studies of MSCs had observed that CD34 expression disappeared when the freshly isolated MSCs were expanded in culture [61 –63]. The loss of CD34 expression has continued to be noted in endothelial cells [64] and in MSCs of various tissue origins [65,66]. Thus, although there is not a current consensus, some researchers believe that CD34 is expressed in tissue-resident MSCs and that negative expression of CD34 is a consequence of in vitro culture [67]. Given the emerging evidence showing the cell culture-associated loss of CD34 expression, one may presume that the CD34-positive cells in the blood apheresis product reported by Saw et al. might be hematopoietic progenitor cells or unexpanded PB MSCs.
However, the biological property that most uniquely identifies MSC is their capacity for trilineage mesenchymal differentiation, which was determined in functional assays [58].
Functional analyses will help clarify the identity of the progenitor cells derived from PB apheresis, which are intended to be used fresh without in vitro expansion. Although four clinical studies have obtained satisfactory outcomes with PB-derived progenitors, these cells could not be conclusively identified as MSCs, due to inadequate functional characterization of the cellular components used in the clinical studies. In addition, only two of the in vivo animal studies provided detailed immunophenotypic and functional information about the PB-derived progenitors. Some in vitro studies did not perform functional assays, which rendered the identification of the putative MSCs untenable; these studies were consequently removed from the current review [68].
As shown in Table 3, some in vitro studies seemed to focus only on the chondrogenic ability of the PB-derived progenitor cells [32,33]. These studies analyzed the surface markers and performed the chondrogenic differentiation test, but neglected to conduct the osteogenic and adipogenic assays. Some other studies only conducted the trilineage differentiation tests without providing immunophenotyping data [26,27,29].
Although Trivanovic et al. conducted a full set of immunophenotyping and functional assays, the surface markers of the PB progenitors they identified did not meet the ISCT criteria of human MSCs [28]. The authors reported that a considerable proportion (40%) of progenitors were CD34-positive, CD90-negative, and slightly positive for CD105 (6%–10%). Because the cells used for the cytometry analysis were at Passages 3 to 6 in the study by Trivanovic et al., and the MSC surface phenotype tends to change following in vitro cultivation [69], additional experiments using progenitors at the same passage are needed to clarify the identity of the chondrogenic MPCs. Based on these incomplete in vitro data, the PB progenitors can only be regarded as chondrogenic MPCs that possess the potential to differentiate into at least one of the mesenchymal lineages (ie, chondrogenesis).
Additional experiments and detailed procedures must be reported to obtain replicable results and successfully translate stem cell research from the bench to the clinic [70,71].
Enhanced cartilage regeneration was observed in rabbits, sheep, and a racehorse treated with chondrogenic PB-derived progenitors [14,25,47]. However, most clinical applications were reported in case reports, with the exception of one RCT and two controlled animal studies. Furthermore, it is problematic to attribute the enhanced cartilage repair solely to the delivery of PB-derived progenitors, because the treatment protocol in some studies involved a combination of PB progenitor injections and BM stimulus techniques [43 –46]; the latter is well known to promote cartilage healing.
Hence, a reliable clinical trial design with reproducible experimental conditions and well-characterized cellular components of the PB will be beneficial to clarify the mechanisms responsible for the favorable outcomes and promote the advancement of PB-derived cell-based treatment for cartilage engineering.
This systematic review has potential biases. First, the databases searched in this study were limited to PubMed, EMBASE, and the Web of Science. Therefore, studies that were exclusively published in other databases and non-English articles might have been overlooked.
Second, some articles were excluded due to a lack of indispensable experiments, such as immunophenotyping analyses, chondrogenic differentiation data, or full trilineage differentiation data. The removal of certain articles might have reduced the number of potential studies supporting or opposing the existence of MSCs or chondrogenic MPCs.
Third, publication bias remains a concern in meta-analyses or systematic reviews of MSCs because some journals are less likely to publish negative results [72]. Thus, relevant studies of PB-derived progenitor cells that reported negative results may have been missed in the present work. Therefore, our results must be interpreted with caution.
In summary, this systematic review suggests the existence of PB MSCs in animals based on both in vivo and in vitro data. The presence of PB MSCs in humans is less certain, due to the similar numbers of in vitro studies with positive and negative outcomes and uncharacterized PB-derived progenitor cells that were used for cartilage repair in patients. However, the majority of the studies support the presence of circulating chondrogenic MPCs in humans and animals. These results identify the PB as a potential source of chondrogenic progenitor cells. Guidelines and standardized protocols for laboratory studies and clinical trials of PB-derived progenitor cells are currently lacking, and considerable efforts are required to address this need.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (No. 51273004; No. 31200725).
Author Disclosure Statement
No competing financial interests exist.