
Abstract
Osteoporosis is a disease that affects 35% women and 20% men aged more than 65 years. Reduction in bone formation and increased bone resorption are known factors that drive osteoporosis, but recent studies suggest a positive correlation between bone marrow adipose tissue (MAT) and osteoporosis. Previously, we have observed that tonsil-derived mesenchymal stem cells (T-MSCs) reduce MAT in a mouse model of bone marrow depletion. That prompted us to investigate on the senile osteoporosis to characterize the bone-forming effect, as well as MAT-reducing effect of T-MSCs. In a mouse model of senescence-accelerated mouse prone 6 (SAMP6), we injected T-MSCs or T-MSC conditioned medium (CM) through tail vein and examined changes in bone microstructure using micro-CT scan and hematoxylin & eosin (H&E) staining. Biochemical markers of osteoporosis, deoxypyridinoline (DPD) and osteocalcin, were examined by ELISA. Results demonstrated attenuation in the progression of osteoporosis, in part, by sustaining osteocalcin production and by blocking MAT accumulation. Increase in matrix mineralization was determined using in vitro culture of murine preosteoblast cell line by treatment of T-MSC CM. Interestingly, T-MSC CM induced continuous weight loss and selectively reduced visceral adipose tissue mass. Finally, antiadipogenic effects of T-MSC CM were determined in vitro. In conclusion, regulation of bone together with MAT could be considered as a new therapeutic option for the treatment of senile osteoporosis and this report may provide a framework for future cell therapy using T-MSCs.
Introduction
T
Expansion of MAT is found with aging, anorexia nervosa, and diabetes [6 –8]. Association between MAT and bone health has been suggested, and advances in imaging technology have established MAT as a novel noninvasive biomarker for the osteoporosis risk assessment [9]. Imbalance between adipogenesis and osteoblastogenesis is one of the underlying mechanisms that results in MAT expansion [10]. This is because both adipocytes and osteoblasts are generated from mesenchymal stem cells (MSCs) and are reciprocally regulated by several common factors such as bone morphogenetic proteins, Wnt/β-catenin, and peroxisome proliferator-activated receptor-gamma (PPARγ) [11,12]. Emerging investigation on the novel factors and signaling pathways, which regulate bone marrow (BM) osteoblastogenesis and adipogenesis [13 –15], may provide a framework for the development of novel therapeutic strategies for osteoporosis targeting bone and fat in BM milieu.
MSCs are a good source for regenerative medicine due to their self-renewal and multipotent properties. These cells can be isolated from many sources, including umbilical cord blood, BM, and adipose tissue. Our group has been using human palatine tonsil as a novel tissue source for MSCs (termed T-MSCs). We have demonstrated that T-MSCs adhere to plastic culture surface, express common MSC surface markers (CD73, CD90, and CD105), and can be differentiated into osteoblasts, adipocytes, and chondrocytes [16]. Emerging reports from our laboratory and others also suggest that T-MSCs exhibit tissue regenerative [17 –19] and immunomodulatory effects [2,4,5]. Previously, therapeutic potential of MSCs isolated from BM and adipose tissue on osteoporotic bones was demonstrated [20,21]. Those works have reported an effectiveness of MSCs for the treatment of osteoporosis, although some barriers exist to be developed as a cell therapy. First, they injected MSCs directly into bone marrow cavity, which limits its practice without operation. Next, an effectiveness of human MSCs has not been confirmed in a preclinical model. In addition, previous observation was limited to osteoblasts and osteoclasts, never been extended to MAT regulation.
We have had an interesting observation previously that T-MSCs reduce MAT in mice (in press). Induction of BM ablation using busulfan and cyclophosphamide [22] resulted in massive expansion of MAT in mouse femurs. Treatment of T-MSCs improved BM cellularity, as well as decreased MAT. These observations prompted us to investigate the potential of T-MSCs or T-MSC conditioned medium (CM) to be used for treating osteoporosis.
We used senile osteoporosis mouse model, senescence-accelerated mouse prone 6 (SAMP6), to examine changes in bone and MAT. SAMP6 is a substrain of senescence-accelerated mouse [23] strain, which was developed through the selective inbreeding of AKR/J strain mice [24]. Among the strain, SAMP6 was reported to show a lower peak bone mass compared to SAMP1 or SAMP2 and suggested to be equivalent to the senile osteoporosis in humans based on microdensitometric, chemical, and histologic findings [25]. SAMP6 had been used in many studies on senile osteoporosis, and the usefulness of this mouse strain has been verified [20,26,27].
In this study, we aimed to identify the effectiveness of T-MSCs or T-MSC CM for the treatment of senile osteoporosis. To address this, we examined changes in bone microstructure and production of biochemical markers of osteoporosis following treatment. MAT accumulation was analyzed in parallel. In addition, the effects of T-MSCs on differentiation of osteoblasts or adipocytes were determined in vitro.
Materials and Methods
Animals
Male SAMP6 animals (Japan SLC, Shizuoka Prefecture, Japan) were maintained at 21–23°C and 51–54% humidity with a 12-h light/12-h dark cycle under conventional conditions. Single mouse was housed in a single cage and food and water were supplied ad libitum. Experiments and procedures were approved by the Animal Ethics Committee at Ewha Womans University School of Medicine (ESM 14-0278), and all experiments were performed in accordance with relevant guidelines and regulations. At 7 months of age, when the linear decrease in bone mass was examined [26,27], 24 mice were divided into four groups (6 mice per treatment group) by matching the average body weight to minimize weight variation across treatment groups. Control (200 μL DMEM), T-MSCs (0.8 × 106 cells in 200 μL DMEM), or T-MSC CM (CM harvested from 48-h incubation of 0.8 × 106 cells followed by concentration to 200 μL) was infused through tail vein injection twice a week for 2 weeks. A synthetic PPARγ antagonist, bisphenol-A-diglycidyl ether (BADGE; Sigma-Aldrich, St. Louis, MO), which has been shown to reduce MAT [28,29], was administered through intraperitoneal injection (30 mg/kg) for 14 days. Body weight changes were measured every 2 weeks. Blood and urine were collected at 7, 8, and 9 months of age after fasting for 6 h. At 9 months of age, mice were sacrificed by cervical dislocation and organs were harvested for further analysis.
MSC culture and preparation of CM
Previously isolated and maintained T-MSCs [16,17] were cultured in DMEM high-glucose medium (Welgene, Gyeongsan, Korea) supplemented with 10% FBS, 100 IU/mL penicillin, and 100 μg/mL streptomycin (Welgene). Culture medium was changed every 3–4 days. To collect CM, T-MSCs passaged less than 10 times [30] were cultured until 80% confluency was reached, after which the culture medium was replaced with fresh medium. Cells were incubated for an additional 48 h, and CM was harvested. CM was concentrated 10-fold using a 3-kDa Amicon Ultra centrifugal filter unit (Millipore, Darmstadt, Germany) by centrifugation at 3,800 g for 30 min. Concentrated CM was added to the cells during their differentiation into adipocytes or osteoblasts. Adipose-derived MSCs (AD-MSCs) were generously provided by RNL Bio (Geumcheon-gu, Seoul, Korea), and BM-MSCs were purchased from Severance Hospital Cell Therapy Center (Seodaemun-gu, Seoul, Korea). AD-MSCs were maintained in medium same as T-MSCs and BM-MSCs in DMEM low-glucose medium (Welgene) containing 10% FBS, 100 IU/mL penicillin, and 100 μg/mL streptomycin.
Micro-CT
Mouse femurs were fixed in 4% paraformaldehyde overnight at 4°C and a micro-CT scan was performed using the SkyScan 1172 scanner and associated software (Bruker microCT, Kontich, Belgium).
H&E staining and MAT analysis
Mouse femurs were isolated and fixed with 4% paraformaldehyde solution in PBS. Decalcified and paraffin-embedded femurs were sectioned and then subjected to H&E staining. Fields were taken from H&E-stained sections of distal femur and fat infiltration was calculated using NIH ImageJ software.
Adipocyte differentiation
The 3T3-L1 mouse preadipocyte cell line was purchased from the Korean Cell Line Bank and maintained in growth medium (DMEM high-glucose medium supplemented with 10% FBS, 100 IU/mL penicillin, and 100 μg/mL streptomycin) until 3 days after confluency was reached. Cells were then induced to differentiate in medium supplemented with 4 mg/mL insulin, 0.22 μM dexamethasone, 0.5 mM 3-isobutyl-1-methylxanthine (IBMX), and 0.1 μg/mL Biotin (Sigma-Aldrich). After 3 days, the cells were refreshed with medium supplemented with insulin only. From day 6 until experiments were completed, the cells were maintained in growth medium that was changed every 3–4 days.
Osteoblast differentiation
The MC3T3-E1 mouse preosteoblast cell line was purchased from ATCC (Manassas, VA) and cultured in ascorbic acid-free α-MEM (Gibco, Waltham, MA) containing 10% FBS, 100 IU/mL penicillin, and 100 μg/mL streptomycin. Once the cells were confluent, osteoblast differentiation was induced using osteogenic induction medium that consisted of α-MEM (Welgene) supplemented with 10% FBS, 100 IU/mL penicillin, 100 μg/mL streptomycin, 50 μM ascorbate-2-phosphate (Sigma-Aldrich), 10 mM β-glycerophosphate (Sigma-Aldrich), and 0.1 μM dexamethasone. Medium was changed every 3–4 days until experiments were completed.
RNA extraction and cDNA synthesis
RNA extraction was performed using a NucleoSpin RNA Kit (Macherey-Nagel, Düren, Germany) according to the manufacturer's instructions. The concentrations and purities of RNA samples were determined using a BioPhotometer D30 (Eppendorf, Hamburg, Germany). For cDNA synthesis, 1 μg RNA was mixed with ReverTra Ace-α reaction solution containing 1× RT buffer, 1 mM dNTP mix, 0.5 μM oligo dT primer, 0.5 U/μL RNase inhibitor, and ReverTra Ace reverse transcriptase (Toyobo, Osaka, Japan) and then incubated at 30°C for 10 min, at 42°C for 20 min, and at 99°C for 5 min.
Real-time quantitative PCR
Expression of target genes was assessed using the StepOnePlus Real-Time PCR System (Applied Biosystems, Foster City, CA). For each reaction, a primer pair (0.4 μM) and 1 × SYBR Green Realtime PCR Master Mix (Toyobo) were added to the prepared cDNA. To amplify cDNA, samples were subjected to 40 cycles of 15 s denaturation at 95°C and 1 min amplification and signal acquisition at 60°C. Cycle threshold (Ct) values were obtained and the relative expression level of a target gene was determined as: 2(housekeeping gene Ct–target gene Ct). The primer sequences are available upon request.
Oil Red O staining
To measure lipid accumulation, adipocytes were rinsed with PBS and fixed in 4% paraformaldehyde for 5 min at room temperature. Fixative was discarded and the wells were dried completely. Oil Red O (Sigma-Aldrich) was dissolved in isopropanol to make an 8.5 mM stock solution that was mixed with distilled water at a 6:4 ratio and filtered through a 0.45 μm filter to make up the working solution. The working solution was added to the wells and incubated for 10 min at room temperature. The Oil Red O solution was removed, and the wells were rinsed with distilled water four times. The stain was eluted with 100% isopropanol before measuring the absorbance at 540 nm.
Alizarin Red S staining
Matrix mineralization in osteoblasts was examined by Alizarin Red S staining. After washing with PBS, the cells were fixed with 60% isopropanol for 5 min at room temperature. Cells were stained with a 2% Alizarin Red S (Sigma-Aldrich) solution prepared in distilled water (pH 4.1–4.3) by incubating them for 5 min or until matrix staining was visible. The cells were rinsed with distilled water four times. The stain was eluted with 10% cetylpyridinium chloride (Sigma-Aldrich) before measuring the absorbance at 570 nm.
Enzyme-linked immunosorbent assay
Mouse urine and plasma specimens were collected after 6 h of fasting. Quantitation of deoxypyridinoline (DPD) in mouse urine was performed using a MicroVue DPD EIA Kit (Quidel, San Diego, CA). For the correction of urine concentration, DPD values were divided by creatinine values measured by a MicroVue Creatinine Assay Kit (Quidel). Osteocalcin concentration in mouse plasma was determined using a Mouse Osteocalcin ELISA Kit (Immutopics, San Clemente, CA) according to the manufacturers’ instructions.
Statistical analysis
The Student's t-test or one-way ANOVA in conjunction with Tukey's post hoc tests was performed as appropriate using GraphPad Prism 5.0 (GraphPad Software, San Diego, CA). Data are presented as the mean ± standard error of the mean.
Results
Administration of T-MSCs or T-MSC CM attenuated the progression of senile osteoporosis in vivo
It is previously reported that SAMP6 mice develop the characteristics of senile osteoporosis at approximately 7 months of age [27]. Seven-month-old male SAMP6 mice were divided into four groups and received treatment of T-MSCs, T-MSC CM, or BADGE, a MAT-reducing reagent. Effects of those treatments on the development of osteoporosis were determined 8 weeks following injection (Fig. 1A). To examine the changes in bone microstructure, micro-CT scans were obtained. The representative 2D and 3D images of the distal femur shown in Fig. 1B demonstrate increased volume of the trabecular bone in the MSC-, CM-, and BADGE-treated groups compared to the control group. Quantitative analysis of micro-CT images showed a significant increase in bone volume and decrease in porosity following MSC treatment. CM or BADGE injection did not affect bone volume. No changes in trabecular thickness or separation were observed (Fig. 1C).
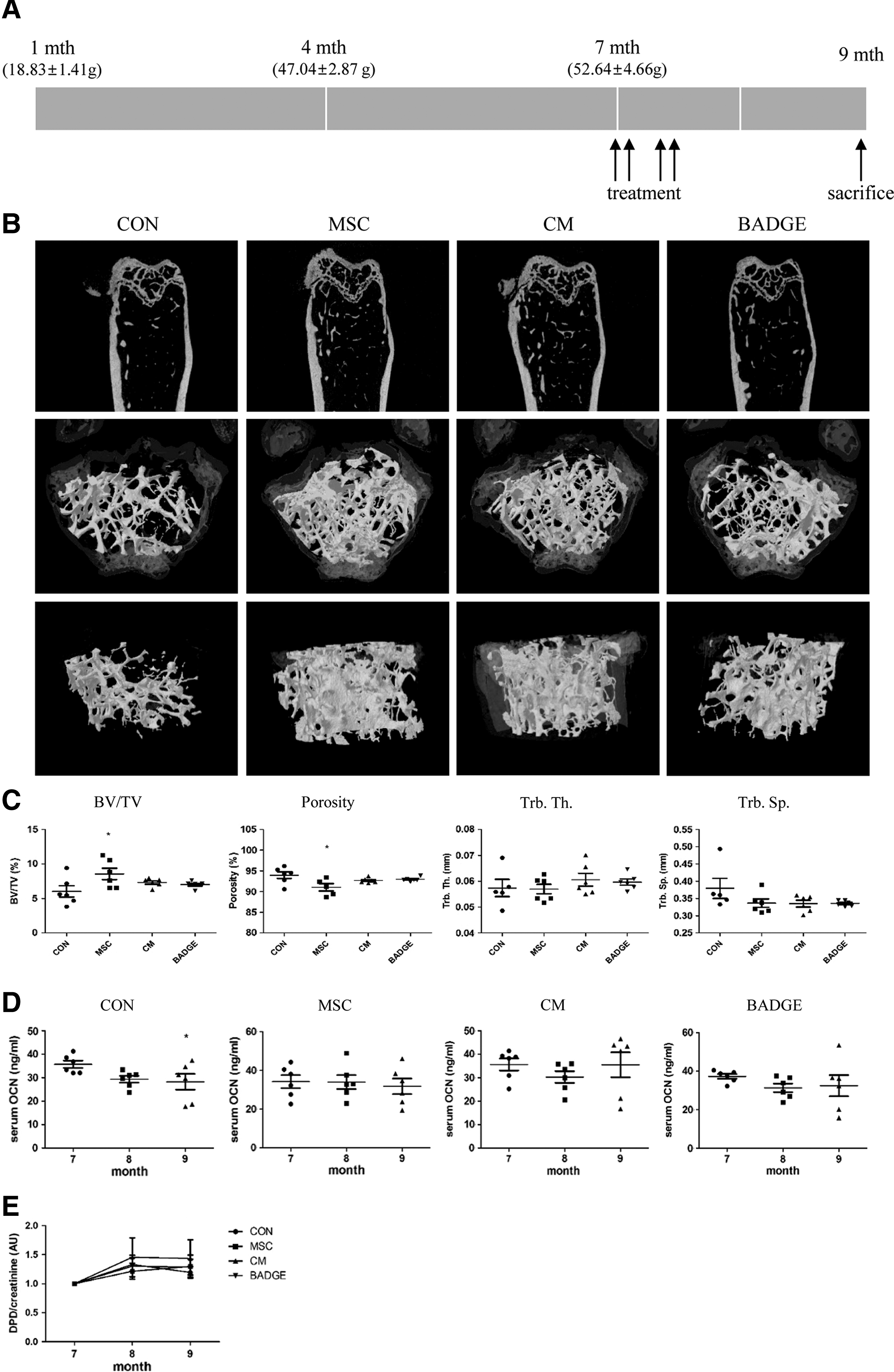
T-MSCs or CM injection attenuated progression of senile osteoporosis.
Next, we examined biochemical markers for osteoporosis. Serum osteocalcin was measured as a biomarker for bone formation at the time of injection (7 months) and at 1 and 2 months postinjection (8 and 9 months). Our data show that the serum osteocalcin level decreased with aging in control mice, but not in any of the treated animals, suggesting that reduction in bone formation with aging was attenuated by MSC, CM, or BADGE injection (Fig. 1D). Urine DPD levels, which reflect bone resorption, were also measured and normalized to the level before treatment began. However, no differences in DPD level were found in each treatment group compared to the control (Fig. 1E), demonstrating that bone resorption was not altered by MSC, CM, or BADGE treatment. These data suggest that T-MSC injection prevents osteoporosis by increasing bone volume and maintaining bone formation rather than inhibiting bone resorption. CM or BADGE treatment also maintains the bone microarchitecture to a lesser extent by attenuating the loss of bone formation associated with aging.
T-MSC CM promoted osteoblast matrix mineralization in vitro
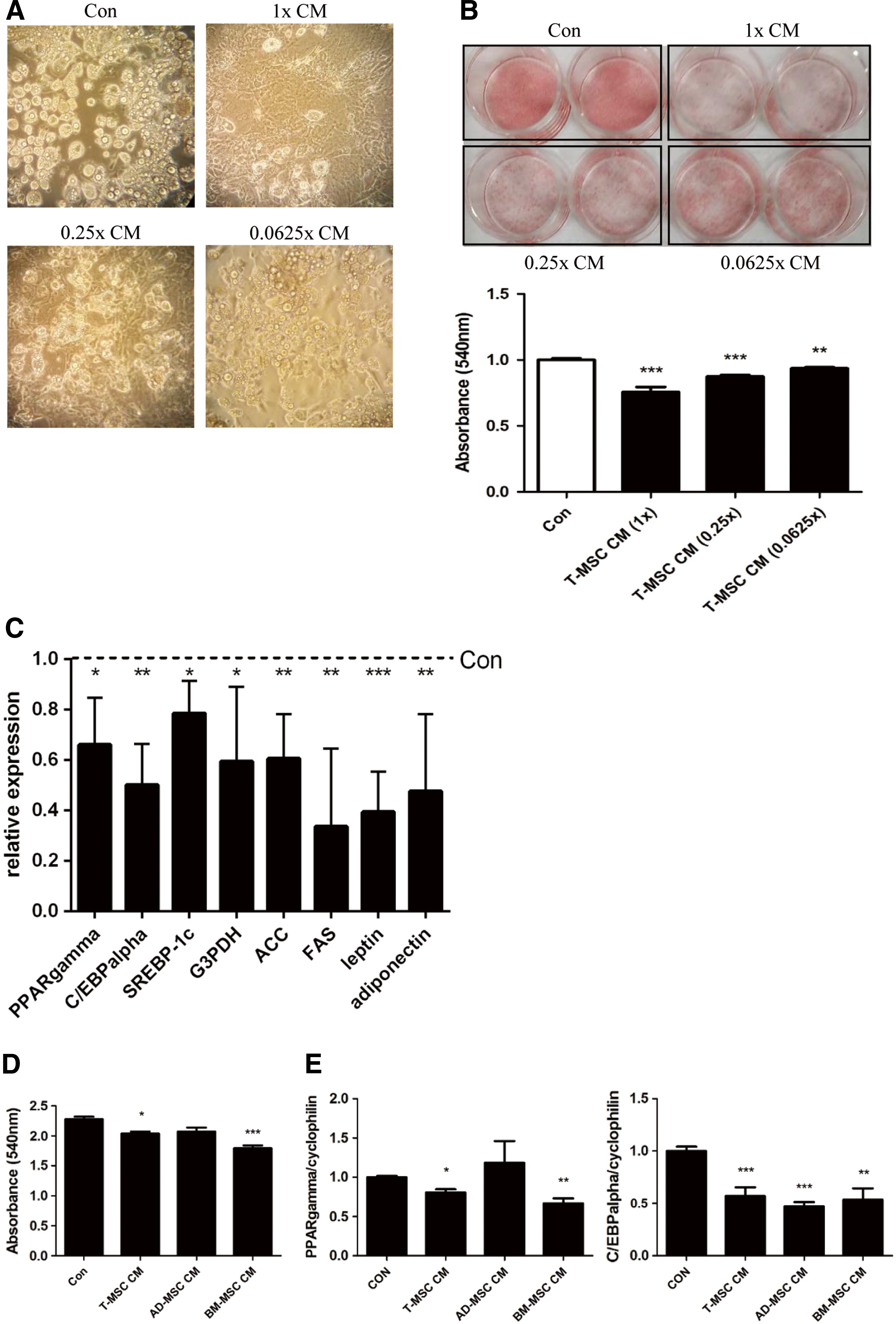
To investigate whether T-MSC CM affects osteoblast differentiation and/or maturation, murine preosteoblast cell line MC3T3-E1 was used. Cells were cultured with CM from various tissue sources for 14 days while inducing osteoblast differentiation, which was measured by Alizarin Red S staining, a method that quantifies mineral deposition in cells. Our results showed that T-MSC CM improved osteoblast matrix mineralization. CM collected from AD-MSCs was less effective than T-MSC CM, while BM-MSC CM showed comparable effects to T-MSC CM (Fig. 2A). Examination of varying amounts of T-MSC CM demonstrated a dose-dependent increase in osteoblast matrix mineralization as the amount of T-MSC CM was increased (Fig. 2B).

T-MSC CM promoted osteoblast matrix mineralization. CM was collected after a 48-h incubation of T-MSC, AD-MSC, or BM-MSC and then concentrated. While MC3T3-E1 cells (Con) were differentiating, CM secreted from the equivalent amount of MSCs (1 × ) that was diluted four times (0.25 × ) or 16 times (0.0625 × ) was added to the medium.
The expression of genes involved in osteoblast differentiation and function was also examined during the time course of MC3T3-E1 cell differentiation. Our data show that T-MSC CM did not modulate runt-related transcription factor 2 (RUNX2) expression, but it significantly reduced osterix expression. Significant increase in the expression of the noncollagenous matrix proteins, bone sialoprotein (BSP-1 and -2), was found following T-MSC CM treatment, while alpha 1 chain of type I collagen (col1a1) and alkaline phosphatase (ALP) expression remained unchanged (Fig. 2C). These data suggest that T-MSC CM may improve matrix mineralization without affecting cell differentiation.
T-MSCs or T-MSC CM injection reduced fat accumulation in the femur
Given that increased MAT is a known feature of osteoporosis [9] and BADGE treatment was shown to reduce MAT [28], we investigated whether treatment with T-MSCs or T-MSC CM could reduce the accumulation of marrow adipocytes in the femur. Histology of distal femurs by H&E staining revealed that the percentage and average size, but not the number, of marrow adipocytes were significantly reduced by T-MSCs or T-MSC CM infusion (Fig. 3A, B). However, a reduction in marrow adiposity following BADGE treatment, which has been reported previously [29], was not observed in this senile osteoporosis mouse model (Fig. 3A, B). These demonstrate that T-MSCs could ameliorate osteoporosis by reducing MAT.
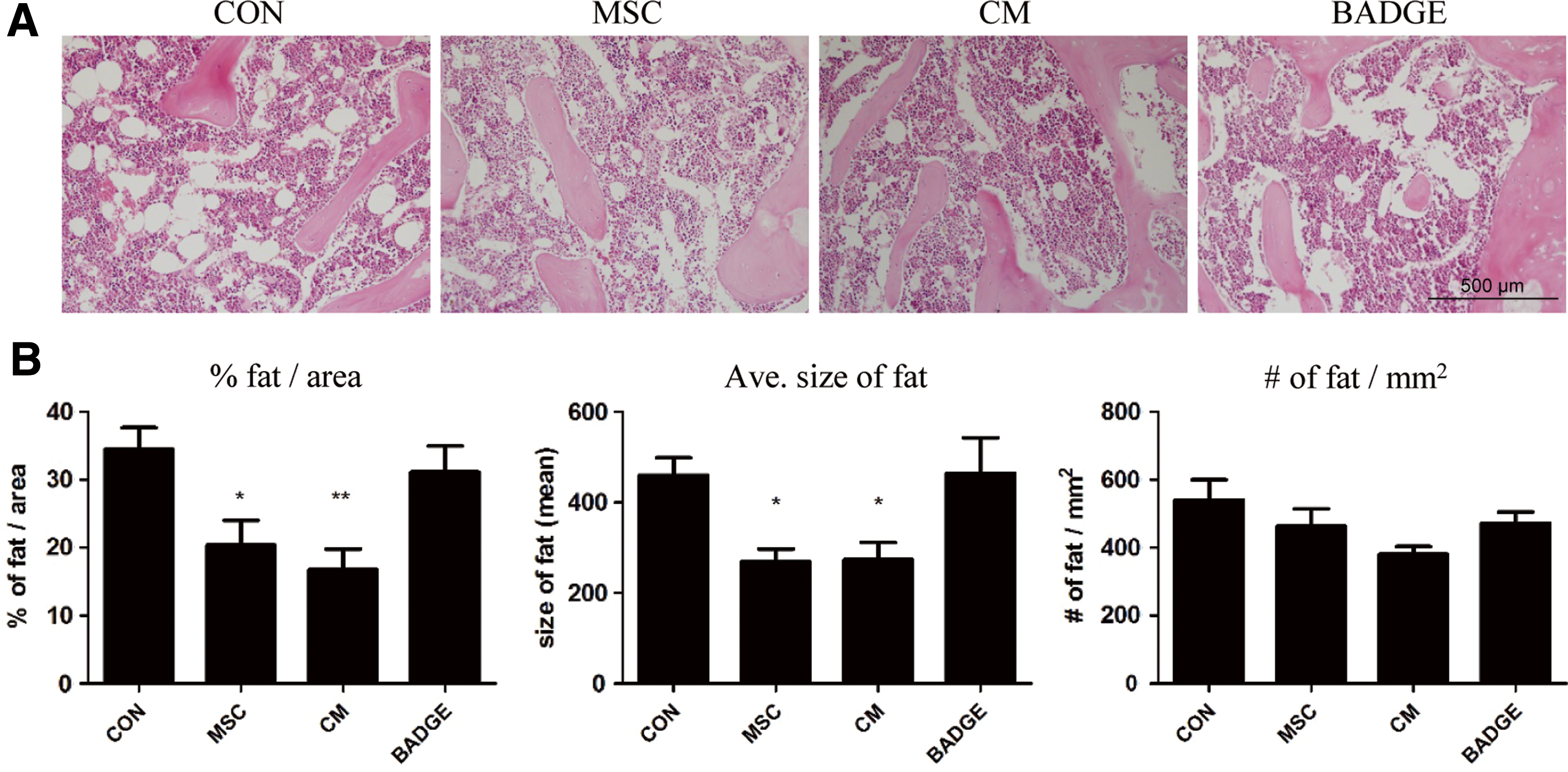
T-MSCs or CM injection reduced adiposity in bone marrow.
T-MSC CM promoted weight loss in SAMP6 mice
To determine whether the treatments raised any whole body effects, SAMP6 mouse body weight was measured every 2 weeks. Due to a big variation in mouse weights as shown in Fig. 4A, we examined the percent of body weight changes in each group (Fig. 4B). BADGE treatment resulted in a significant reduction in body weight compared to the control group at 9 months (Fig. 4B). We also found that CM treatment resulted in a continuous reduction in mouse body weight over time that started at approximately 1.5 months after treatment initiation. Weight reducing effect of BADGE seems rebound at some stages, but remained significant at termination of the experiment. A decrease in body weight was also observed in control and T-MSC-injected mice, but these were not significant and more likely due to injection-related artifacts (Fig. 4C). To determine the changes in adipose tissue weights, tissues were isolated and weighed from four different adipose depots. A significant decrease in the accumulation of fat was observed in visceral areas, namely mesenteric (mAT), epididymal (eAT), and retroperitoneal (rAT), following T-MSC CM administration. BADGE treatment reduced fat accumulation in rAT only. Subcutaneous fat, liver, and skeletal muscle weights were not affected by the treatments (Fig. 4D). These data suggest that selective weight reduction in visceral adipose tissue (VAT) by T-MSC CM treatment may produce beneficial effects in whole body metabolism.
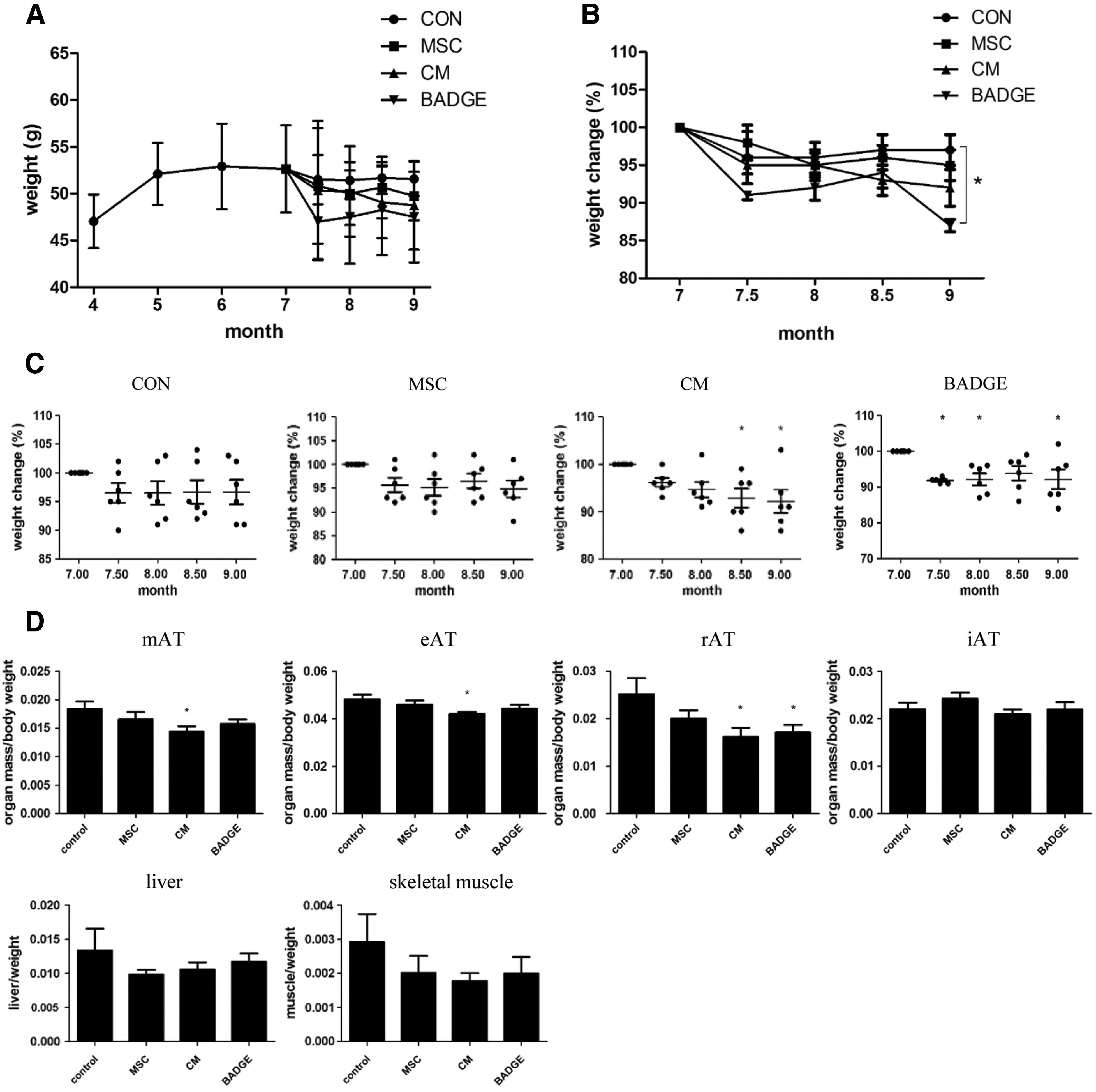
T-MSC or CM injection reduced visceral fat accumulation.
T-MSC CM produced antiadipogenic effects in vitro
Reduction in MAT by T-MSCs or T-MSC CM treatment prompted us to investigate the effects of T-MSC CM on adipocyte differentiation using the murine preadipocyte cell line 3T3-L1. Throughout the 10 days of differentiation, varying doses of T-MSC CM were added. The effects of treatment with T-MSC CM were examined by changes in adipocyte morphology using phase contrast microscopy and comparing inhibition of adipocyte differentiation between T-MSC CM-treated cells and control cells (Fig. 5A). Lipid droplet formation was measured by Oil Red O staining and indicated that T-MSC CM inhibited lipid accumulation in adipocytes in a dose-dependent manner (Fig. 5B).
T-MSC CM inhibited adipocyte differentiation. While 3T3-L1 cells (Con) were differentiating, CM secreted from the equivalent amount of MSCs (1 × ) that was diluted four times (0.25 × ) or 16 times (0.0625 × ) was added into the medium. Adipocyte differentiation and lipid accumulation were evaluated by
The expression of genes involved in adipogenesis was examined using real-time quantitative PCR. Transcription factors (PPARγ, CCAAT-enhancer-binding protein-alpha [C/EBPα], sterol regulatory element-binding protein-1c (SREBP-1c), lipogenic enzymes (glycerol-3-phosphate dehydrogenase [G3PDH], acetyl-CoA carboxylase [ACC], fatty acid synthase [FAS]), and adipokines (leptin and adiponectin)) were all expressed at significantly lower levels than in control cells following incubation with T-MSC CM (Fig. 5C). We also confirmed that antiadipogenic effect of T-MSC CM was mediated by factors secreted from T-MSCs rather than FBS contained in culture medium (Supplementary Fig. S1A; Supplementary Data are available online at
The antiadipogenic effects of T-MSC CM were compared with CM collected from AD-MSCs and BM-MSCs by examining lipid accumulation and adipogenic transcription factor expression. Our data show that the antiadipogenic effect of T-MSC CM was superior to that of AD-MSC CM, but similar to BM-MSC CM (Fig. 5D, E). These results demonstrate that T-MSCs secrete factors that inhibit adipogenesis.
Discussion
In this study, we identified the pro-osteogenic and antiadipogenic effects of T-MSCs or T-MSC CM in a mouse model of senile osteoporosis, which is characterized by reduced bone mass and increased fat mass in the BM. Our data demonstrate an improved microarchitecture in the femurs of mice treated with T-MSCs or T-MSC CM. Osteocalcin levels were maintained across the experiment, suggesting that the progression of osteoporosis was attenuated by the treatments. MAT expansion was inhibited by T-MSCs or T-MSC CM injection mainly by reducing the size of adipocytes. Systemic effects were observed in T-MSC CM-treated animals, which showed a moderate, but continuous, decrease in body weight and a significant reduction in fat accumulation in the abdominal area. Our findings indicate that T-MSCs secrete factors involved in preventing osteoporosis by maintaining bone formation and reducing marrow adiposity.
Human tonsil is an attractive source of MSCs [16,32]. This tissue is typically generated as medical waste from tonsillectomy procedures and yields high numbers of MSCs (8–10 × 108 cells from a 2 × 1.5 × 1.5-cm tissue, unpublished data). We demonstrated previously that T-MSCs could be used for MSC banking as they retain MSC characteristics through cryopreservation/thawing and form chimeras in mixed cultures [16]. It is confirmed by a long-term culture of T-MSCs that MSC-specific surface marker expression, doubling time, and differentiation capacity of T-MSCs maintained until passage 10 [30]. Moreover, it is also reported that T-MSCs proliferate twice faster compared to AD-MSCs [33].
Accumulating evidence from our laboratory and others has demonstrated the tissue regenerative (ie, liver, pancreas, and parathyroid gland) and immunomodulatory effects of T-MSCs [17,23,33 –36]. This report expands the therapeutic potential of T-MSCs to diseases related to bone and/or adipose tissue. Moreover, we show the effectiveness of T-MSC CM that is more easily handled and harvested than MSCs. Comparison of CM collected from different MSC sources suggests that the efficacy of T-MSC CM is superior to AD-MSC CM, but similar to BM-MSC CM.
MSCs function by homing and self-differentiation, as well as by secreting effector molecules. Results from this study provide insight into which functions are performed by the cells themselves versus those mediated by secretory factors. This is supported by our data showing that bone volume in mouse femurs was altered only in the T-MSC-treated group. In contrast, a reduction in BM adiposity was observed in both T-MSC- and T-MSC CM-injected groups. These data suggest that T-MSCs have potentially self-differentiated into bone-forming cells, resulting in a further decrease in marrow adiposity. In contrast, T-MSC CM could have affected both osteoblasts and adipocytes through secretory factors. Our in vitro data show that T-MSC CM upregulates BSP-1 and -2 expression, but not transcription factors involved in osteoblastogenesis such as RUNX2 or osterix. Evidence suggests that BSP transcription is regulated by hormones, growth factors, or cytokines [37 –39]. Further studies are required to identify effector molecules secreted by T-MSCs and to better understand their efficacy toward inducing osteoblast differentiation and treating osteoporosis.
Interestingly, T-MSC CM injection resulted in decreased visceral adiposity together with marrow adiposity. Although adipocytes from two different depots exhibit distinct gene expression profiles [31,40], both have a common function regarding lipid accumulation. Thus, T-MSC secretory factors could reduce the volume of fat in these two depots possibly through the same mechanism. The T-MSC secretome is worth investigating to identify common paracrine factors that regulate adipocytes in two adipose depots. This knowledge would further extend the application of MSC CM to antiobesity or obesity-related diseases such as diabetes, atherosclerosis, or nonalcoholic fatty liver disease.
In this study, we examined the effect of injecting MSCs through the tail vein. Previous reports that demonstrated the effects of BM- or adipose tissue-derived MSCs were performed using intra-BM injection [20,21], which might produce profound effects focused on bone. However, our study design is more relevant for the development of therapies because it is a noninvasive procedure. In addition, through a systemic injection, we could observe the effects of T-MSC CM on reducing body weight and VAT accumulation.
In summary, we have shown that T-MSCs could attenuate the progression of senile osteoporosis by sustaining osteocalcin levels in the circulation, which resulted in improved bone microarchitecture. We also demonstrated the reduction of BM adiposity following T-MSCs or T-MCS CM injection. A dual role of T-MSCs that increase bone mineralization and reduce adiposity suggests that T-MSCs may be a possible and favorable therapeutic source for treating osteoporosis.
Footnotes
Acknowledgments
This work was supported by the Bio & Medical Technology Development Program of the National Research Foundation (NRF) of Korea funded by the Ministry of Science, ICT & Future Planning (2012M3A9C6049823) and the Intramural Research Promotion Grants from Ewha Womans University School of Medicine.
Author Disclosure Statement
The authors declare no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.