
Abstract
Autologous mesenchymal stem cells (MSC) are widely used cell source for cell-based tissue repair and regeneration, but replicative senescence and the associated loss of cellular activity during in vitro expansion limit their therapeutic potential. How to preserve or even enhance the proliferation and differentiation ability of MSC from aged donors without genetic modification remains a challenge to meet clinical need. MSC isolated from human fetal tissues (hFMSC) exhibit higher proliferation and differentiation activities even in prolonged in vitro culture, which might be modulated by autocrine/paracrine action. In the present study, we hypothesized that the bioactive factors secreted by hFMSC, collectively named as hFMSC secretome (HFS), could possess beneficial effect on human adult MSC (hAMSC) undergoing replicative senescence, thus promoting their capability of proliferation and differentiation. HFS was prepared by centrifugation of hFMSC conditioned medium, followed by column-based concentration, and the total protein content of the HFS was quantified to standardize treatment concentration. When compared with hAMSC secretome (HAS), HFS treatment significantly reduced senescence associated-β-galactosidase expression and activity (senescence marker) and enhanced cell proliferation and osteogenic differentiation potential of hAMSC in prolonged in vitro culture. Cellular studies revealed concomitant activation of sirt1 and foxo3a in hAMSC after HFS treatment, which was associated with upregulation of p21 and downregulation of bax and p53. The changes of these senescence associated markers suggested that HFS, but not HAS, could ameliorate replicative senescence of hAMSC in vitro. In nude mice, HFS pretreatment restored the osteogenic ability of senescent hAMSC. Tumor xenograft model revealed that HFS did not promote tumor growth. In conclusion, this study suggests that HFS could be an effective and safe method to overcome replicative senescence and facilitate the therapeutic potential of hAMSC.
Introduction
I
Mesenchymal stem cells (MSC) have been the limelight in tissue regenerative medicine because of reliable supply, multiple differentiation potentials to replace damaged tissues, migration ability toward injured sites, and immunomodulatory effect [9]. However, with no exception, MSC also undergo senescence process during in vivo culture with the aim to obtain sufficient cell numbers for cell therapy, which is manifested as enlarged and flatten cell morphology, reduced proliferation and differentiation activities, and altered genetic and secretory profiles [10]. On the contrary, fetal MSC isolated from first-trimester fetal tissues exhibited higher proliferating activity and stronger stemness phenotypes even after prolonged culture, implying that fetal MSC were less susceptible to replicative senescence, which might be due to their higher telomerase activity and longer telomeres [11]. Recent evidence suggested that the biological activities of MSC could be attributed to their secreted bioactive factors. It was reported that MSC secreted hepatocyte growth factor (HGF) and insulin-like growth factor 1 (IGF-1) for antiapoptosis [12] and fibroblast growth factor 2 for antifibrosis [13]. The immunomodulatory effect of MSC on T cells or NK cells could be partly attributed to the presence of prostaglandin E2 in secretome [14,15]. Leukemia inhibitory factor (LIF) and platelet-derived growth factor (PDGF) in MSC secretome have drawn more attention because they play important role in maintaining stemness [16] and senescence [17], respectively. There is no consensus on the standard method to track MSC senescence upon prolonged culture. Increase of senescence associated-β-galactosidase (SA-β-gal) expression and activity is one of the well-reported indicators of senescence [18]. Sirtuin 1 (sirt1) is a conserved NAD+ dependent class III histone deacetylase, which has implication in aging and cellular senescence [19]. Recent study suggested that genetic modified aged MSC overexpressing Sirt1 enhanced the therapeutic outcome for myocardial infarction in rat model [20].
To the best of our knowledge, it remains unclear whether MSC senescence could be modulated by autocrine/paracrine action, however, it is often perceived that nonsenescent MSC will secrete more bioactive factors and result in better outcome of tissue repair and regeneration. In this study, we hypothesized that secretomes from human fetal MSC (HFS) might consist of bioactive ingredients, which could ameliorate replicative senescence and, thus, enhance the proliferation and osteogenic ability of aged adult MSC.
Materials and Methods
Cell isolation and cultivation
Human adult bone marrow aspirate was used for the isolation of human adult MSC (hAMSC). The use of human adult samples was approved by Joint CUHK-NTEC Clinical Research Ethics Committee (ethical approval code CRE-2010.248). In brief, 5–10 mL bone marrow aspirate was collected from three aged donors in the operating theatre and subjected to gradient centrifugation with Ficoll-Paque Premium (GE Healthcare) as previously reported in clean room (International Organization for Standardization class 7) [21]. The cells were cultured in complete CTS™ KnockOut™ DMEM (Life Technologies, Cell Treatment Therapy grade) supplemented with 10% fetal bovine serum (FBS), 1% penicillin and streptomycin (P/S), and 1% GlutaMAX. At 70%–80% confluence, the cells were subcultured at 5,000 cells/cm2. Human fetal MSC (hFMSC) were recovered from our cryopreserved stock [21]. Human prostate cancer cell lines, PC3, were obtained from the American Type Culture Collection. PC3 were cultured in RPMI-1640 medium supplemented with 10% FBS and 1% P/S. Cell line authentication was done before the experiments. Chemicals and reagents were purchased from Sigma-Aldrich unless otherwise specified.
MSC characterization
The stem cell properties of hAMSC were determined by multilineage differentiation potentials under defined conditions and flow cytometry for surface phenotypes [22]. MSC seeded at 5,000 cells/well in six-well plate were subjected to either culture medium (control group) or induction medium (induction group). The osteogenic induction medium consisted of complete KO-DMEM supplemented with 100 nM dexamethasone, 50 mM L-ascorbic acid-2-phospate, and 20 mM b-glycerol phosphate; the adipogenic induction medium consisted of complete KO-DMEM supplemented with 500 nM dexamethasone, 50 mM indomethacin, 0.5 mM 3-isobutyl-1-methylxanthine, and 10 mg/mL insulin. The cells were fixed in 4% paraformaldehyde (pH 7.2) at day 14. Calcium nodules stained with 0.5% (w/v) Alizarin red S (pH 4.1 adjusted using ammonia) indicated osteogenesis, while adipogenesis was shown by the presence of intracellular oil droplets stained with 0.3% (w/v) fresh filtered Oil red O. For chondrogenic induction, chondrogenic medium (induction group) consisted of plain KO-DMEM supplemented with 100 nM dexamethasone; 283.9 mM L-ascorbic acid-2-phospate; ITS Premix (Becton Dickinson); 90.88 mM sodium pyruvate; 34.75 mM L-Proline; 500 ng/mL bone morphogenetic protein (BMP-2); 10 ng/mL transforming growth factor beta 3 (TGF-β3) (both from R&D Systems); and 1% P/S. The accumulation of extracellular matrix (proteoglycan aggrecan) was stained with Alcian blue (1% w/v) overnight. Surface phenotypes, including CD14, CD45, CD34, CD44, CD90, and CD105 (BD Biosciences), were used to determine the purity of MSC.
Coculture of hFMSC and hAMSC
Equal number of hFMSC and hAMSC (5 × 104) were plated onto the insert (0.4 μm pore size; BD Biosciences) and companion 24-well plate separately. hFMSC were treated with 2 mg/mL mitomycin C for 2 h to cease cell proliferation before indirect coculture. After washing with phosphate-buffered saline (PBS) thrice, the inserts were placed into the companion plates. The hAMSC were subjected to analysis for gene expression and cell proliferation after coculture for the designated period.
Preparation of MSC secretome
The preparation of the condition medium of hFMSC or hAMSC was modified from previous report [23]. In brief, the MSC at 90% confluence in 175 cm2 culture flasks were washed three times with PBS to remove residual FBS and cell debris. For every flask, the cells were incubated in 30 mL FBS free alpha-minimum essential medium for another 24 h. The condition media were then collected, and residual cells and debris were removed by centrifugation at 400 g for 5 min. The supernatant was concentrated with Amicon Ultra-15 Centrifugal Filter Device (Millipore) at 4,000 g and 4°C. The protein concentration of the filtrates hFMSC secretome (HFS) and hAMSC secretome (HAS) was quantified with Pierce™ BCA Protein Assay Kit (Thermo Fisher Scientific) at 562 nm. The HFS and HAS were aliquoted and stored at −80°C. hAMSC from three donors were treated with HFS or HAS at protein concentration of 50 μg/mL (Supplementary Fig. S1; Supplementary Data are available online at
Cell proliferation and metabolic activity
The cell number and metabolic activity were determined with direct cell count and Alamar blue assay (Life Technologies), respectively. Briefly, 5 × 103 hAMSC were plated onto 24-well plates and subjected to co-culture or secretome treatment. Then, the hAMSC were harvested at day 2 and 4 for cell counting with hemocytometer. Alamar blue assay was used to determine if hFMSC coculture or secretome treatment affect the metabolic activity of hAMSC, which might interfere with cell proliferation and gene expression. Alamar blue solution was added into the plates and collected after 3 h incubation for spectrometric measurement at 570 nm, with reference wavelength at 600 nm. The change in reduction between the treatment and control groups was calculated according to manufacturer manual to reflect the change in metabolic activity.
Osteogenic induction
For osteogenic induction of MSC, the induction medium was changed into well after MSC grow into full confluence with or without the presence of HFS. The concentration of HFS was controlled in 50 μg/mL. The recipe of induction medium was followed as per the previous study [24]. For staining, the cultured cells were stained by Alizarin red S to assess the mineralized matrix. After discard supernatant, cells were rinsed with PBS one time and fixed in 70% ethanol for 1 h. The fixed cells were stained with 2% Alizarin red S pH 4.6 (Sigma) and stained for 1 h. Then the plates were washed with deionized water and scanned after dry. For the concentration-dependent study, HFS was added in identical concentration (50 μg/mL) abovementioned studies and one with threefold higher concentration (150 μg/mL). Calcium nodules were stained at day 4 posttreatment.
Quantitative real-time polymerase chain reaction
Total RNA was reverse transcribed into cDNA with PrimeScript RT Master Mix (Takara). Quantitative real-time polymerase chain reaction (qPCR) was performed with Power SYBR Green PCR Master Mix (Life Technologies) in StepOnePlus Real Time PCR system. The PCR condition was denatured at 98°C for 15 s, annealing at 60°C for 30 s, elongation at 72°C for 30 s, 40 cycles. The sequence of primers are listed in Table 1.
1. Lonzaric J, T Lebar, A Majerle, M Mancek-Keber, and R Jerala. (2016). Locked and proteolysis-based transcription activator-like effector (TALE) regulation. Nucleic Acids Res 44:1471–1481.
Cell cycle analysis
The hAMSC was cultured in 1% FBS culture medium overnight to synchronize the cell cycle before treatment. After treatment, hAMSC were dissociated into single cells with trypsin and fixed with prechilled 80% ethanol. These cells were stained with Cycletest™ Plus Kit (BD Biosciences) according to the manufacturer's instruction. Cell cycle analyses were carried out with Cytomics FC 500, and data were analyzed using CXP Cytometry List Mode Data Acquisition & Analysis Software (Beckman Coulter).
Measurement of SA-β-gal
Increase of SA-β-gal expression and activity is an indicator of cellular senescent despite the underlying mechanism not well defined [25]. The expression of SA-β-gal at cellular lever was determined with Commercial Staining Kit (Cell Signaling) according to the manufacturer's instruction. In brief, cells grew on six-well plates were fixed with fixative solution for 15 min, washed twice with PBS, stained with 1 mL freshly prepared staining solution, followed by overnight incubation at 37°C incubator. The quantification of SA-β-gal activity was conducted with SA-β-gal Assay Kit (Life Technologies) according to the instruction. Cells were lyzed with two freezing and thawing cycles with liquor nitrogen and 37°C water bath. The cell lysate was allowed to react with o-nitrophenyl-β-D-galactopyranoside at 37°C and the absorbance at 420 nm indicated the relative activity of SA-β-gal.
Ectopic bone formation assay
After long-term treatment with HFS for three consecutive passages (passage 12–15), 1 × 106 hAMSC were mixed with 40 mg of porous hydroxyapatite-tricalcium phosphate (HA-TCP) scaffold (2 × 2 × 2 mm3) (Biocomposites). After 24 h incubation at humidified incubation at 37°C, the scaffolds were implanted at the dorsal site subcutaneously in 12-week-old male Nude mice (four scaffolds per mice). At week 8 post implantation, the mice were terminated and scaffolds were harvested, fixed in 10% neural buffered formalin, and followed by decalcification in 9% formic acid for 3 weeks. Paraffin embedded scaffolds were sectioned at 5 μm and subjected to H&E and immunohistochemical (IHC) staining.
IHC staining and semi quantification
The expression level of collagen type I alpha 1 (COL1A1) and osteoprotegerin (OPG) was determined to reflect the degree of osteogenic differentiation. The deparaffinized sections were subjected to antigen retrieval and quenching as previously reported [26] before overnight incubation with primary antibodies against COL1A1 (Southern-Biotech; 1310-01) and OPG (Abcam; ab73400) at 4°C in PBS supplemented with 10% goat serum (v/v) and 5% (w/v) bovine serum albumin. After incubation with secondary antibody conjugated with horseradish peroxidase (Molecular Probes®, Life Technologies; 1:200), the signal was developed with DAKO DAB and DAB plus Chromogen Solution(DAKO; K3468). All sections were counterstained with hematoxylin. The images were acquired with DM5500B (Leica) and analyzed with Image-Pro Plus (version 6; Media Cybernetics). The expression level of COL1A1 and OPG was presented in the ratio of stained area to the total area of the tissue, which have been excluded the area of HA-TCP scaffold.
Tumor xenograft experiments
1 × 106 cells of Luc-GFP-PC3 suspended in 50 μL PBS with or without HFS (50 μg/mL) were injected (s.c.) at the dorsal site of Nude mice (male, 20–25 g) together with equal volume of Matrigel™ (BD Biosciences). Tumor growth was determined with IVIS 200 in vivo imaging system (Caliper Life Sciences) after intraperitoneal injection of D-Luciferin (30 mg/mL; 5 μL/g). HFS suspended in Matrigel gel was injected as control.
ELISA procedure
LIF and PDGFAA are reported bioactive factors in MSC secretome affecting stemness and senescence, while lamin B1 (LMNB1) was well reported senescence marker [27].The level of secreted LIF, LMNB1, and PDGFAA in HFS and HAS was measured with Commercial ELISA Kits (Lifespan Bioscience; Cat. No. LS-F12286, LS-F16362, LS-F28461) according to the manufacturer's instructions.
Statistical analysis
Statistical analysis between control and treatment groups was performed by Mann–Whitney U test (SPSS software, v21; IBM Corp.). All results were presented as mean ± SD. P ≤ 0.05 was considered as statistically significant.
Results
Characterization of MSC
HAMSC isolated from aged donors expressed MSC markers CD44, CD90, and CD105, but not hematopoietic stem cell markers CD14, CD34, and CD45 (Fig. 1A–F). After incubation in defined differentiation induction medium, the hAMSC could differentiate into osteogenic, adipogenic, and chondrogenic cells as indicated by the positive staining of calcium nodules, intracellular oil droplets, and proteoglycan aggrecan, respectively (Fig. 1G–I). The characterized hAMSC was used for the collection of HAS and other experiments stated below. Three independent hFMSC lines were thawed from our cryopreserved stock, which was isolated, cultured, and characterized as previously described [21]. The HAS (or HFS) from the three hAMSC (or hFMSC) was aliquoted and stored at −80°C until use.
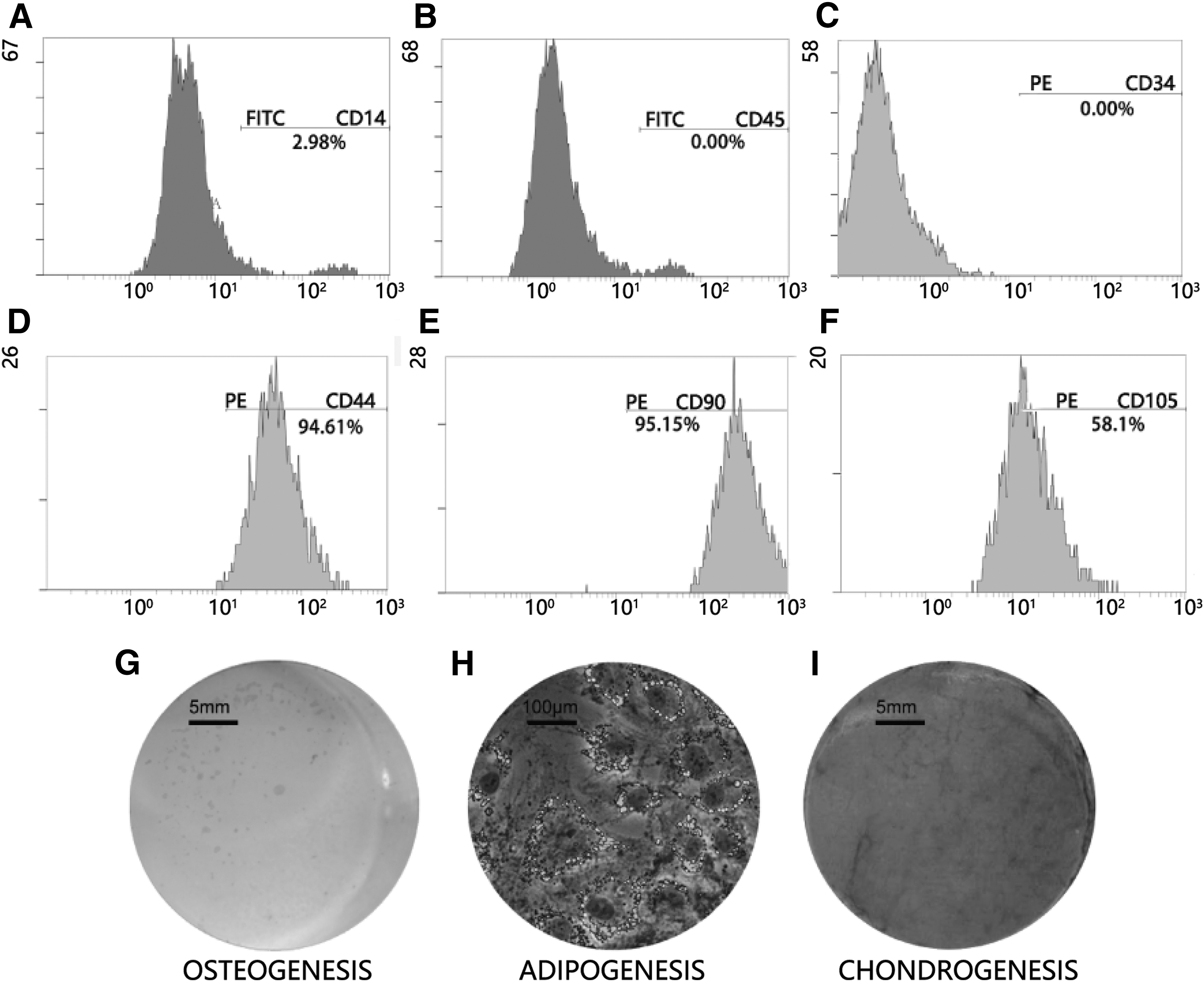
Characterization of hAMSC.
Effects of coculture of hFMSC on hAMSC
The effect of HFS on aged hAMSC was first demonstrated with indirect coculture model. The hFMSC was treated with mitomycin C in advance to cease cell proliferation and minimize its usage of nutrients during the coculture. Alamar blue assay showed slight up in reduction of hAMSC after 1 day of coculture, which was back to normal level (100% compared with hAMSC alone culture), suggesting that the hAMSC had adapted to the coculture environment at the early stage of the experimental period (Fig. 2A). At days 2 and 4, hFMSC coculture significantly promoted cell proliferation of hAMSC as indicated by more cell number per well (Fig. 2B). Meanwhile, qPCR result showed concurrent upregulation of sirt1 and foxo3a in hAMSC 1 day after coculture by nearly five- and fourfold, respectively (Fig. 2C).
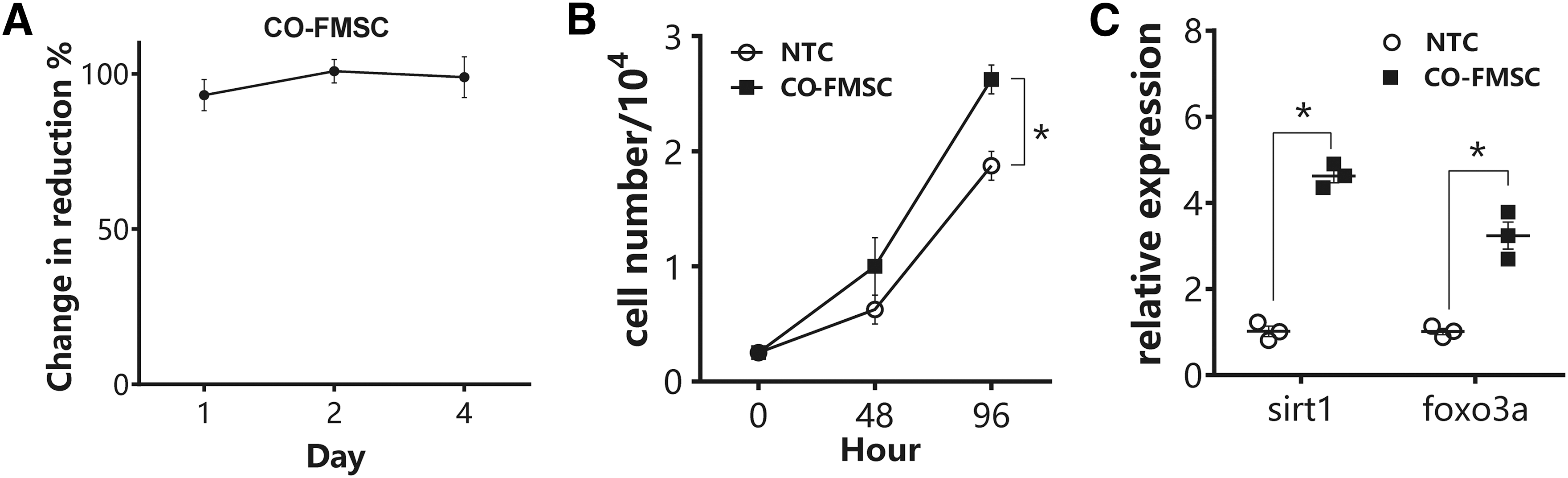
Effects of hFMSC on hAMSC in coculture model. hAMSC was coculture with or without hFMSC in transwell system.
Effects of HFS on hAMSC
The hAMSC was treated with HFS and HAS (as control) under the same culture condition as the coculture model to test whether this phenomenon is solely due to unidirectional effect. Similar to coculture results, HFS enhanced cell proliferation of hAMSC, which reached statistical significance at day 4 (Fig. 3A), and induced the expression of sirt1 and foxo3a significantly at day 1 (Fig. 3B). Furthermore, HFS promoted osteogenic differentiation as indicated by more calcium nodule formation in a time- and concentration-dependent manner (Fig. 3C). qPCR analysis showed consistent result to Alizarin red staining. HFS treatment induced the expression of osteogenic genes, including alp, runx2, and rex1 (Fig. 3D) [28].
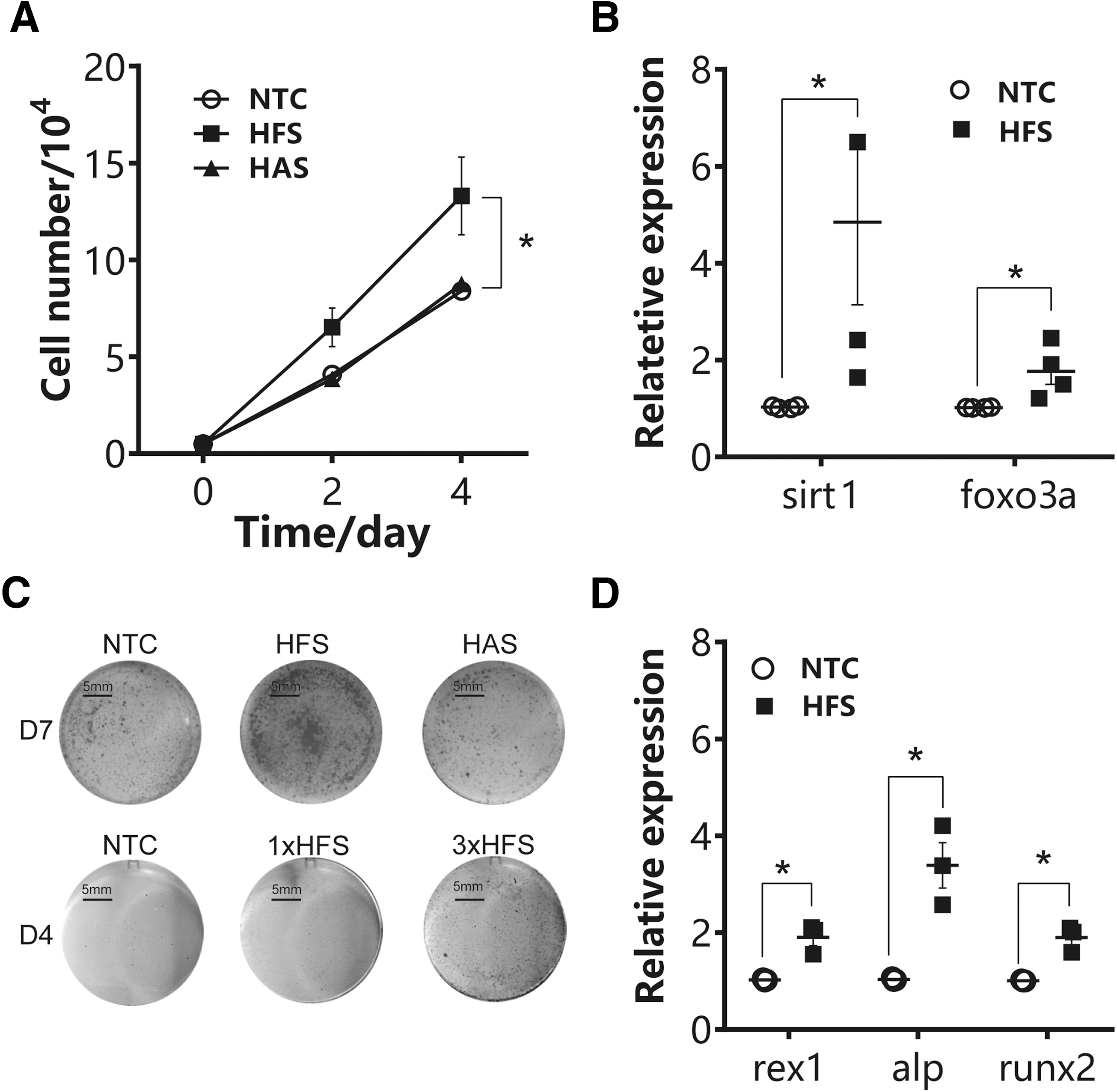
HFS treatment promoted hAMSC proliferation and osteogenic differentiation. hAMSC were treated with either HFS, HAS, or vehicle control.
Effects of HFS on hAMSC senescence-associated cell cycle arrest
Sirt1 is an antiaging gene, which can regulate the expression of SA-β-gal, a representative senescence marker. The overexpression of sirt1 could delay replicative senescence in MSC [29]. Sirt1 was also reported to maintain the differentiation ability of MSC [30]. The data from indirect coculture and secretome treatment studies suggested the presence of the bioactive factors released from hFMSC, which could enhance MSC proliferation through sirt1 signaling. We further analyzed how HFS treatment affected cell cycle pattern of senescent hAMSC. Flow cytometry showed that senescent hAMSC were predominantly arrested at G1 phase. HFS and HAS treatment activated cell division as indicated by increase of hAMSC in G2 and M/S phases by 10.23% and 23.69% respectively (Fig. 4A). HFS was more effective than HAS in promoting cell division. So, we further investigated the expression levels of senescence-related genes in hAMSC with or without HFS treatment (Fig. 4B). HFS treatment downregulated tumor suppressor genes p16 and p53 and proapoptotic gene Bax, but upregulated p21 expression. Furthermore, both staining and enzymatic reaction showed significant reduction of SA-β-gal expression and activity (Fig. 4C, D). Flatten and large senescent hAMSC became more spindle-like after HFS treatment.
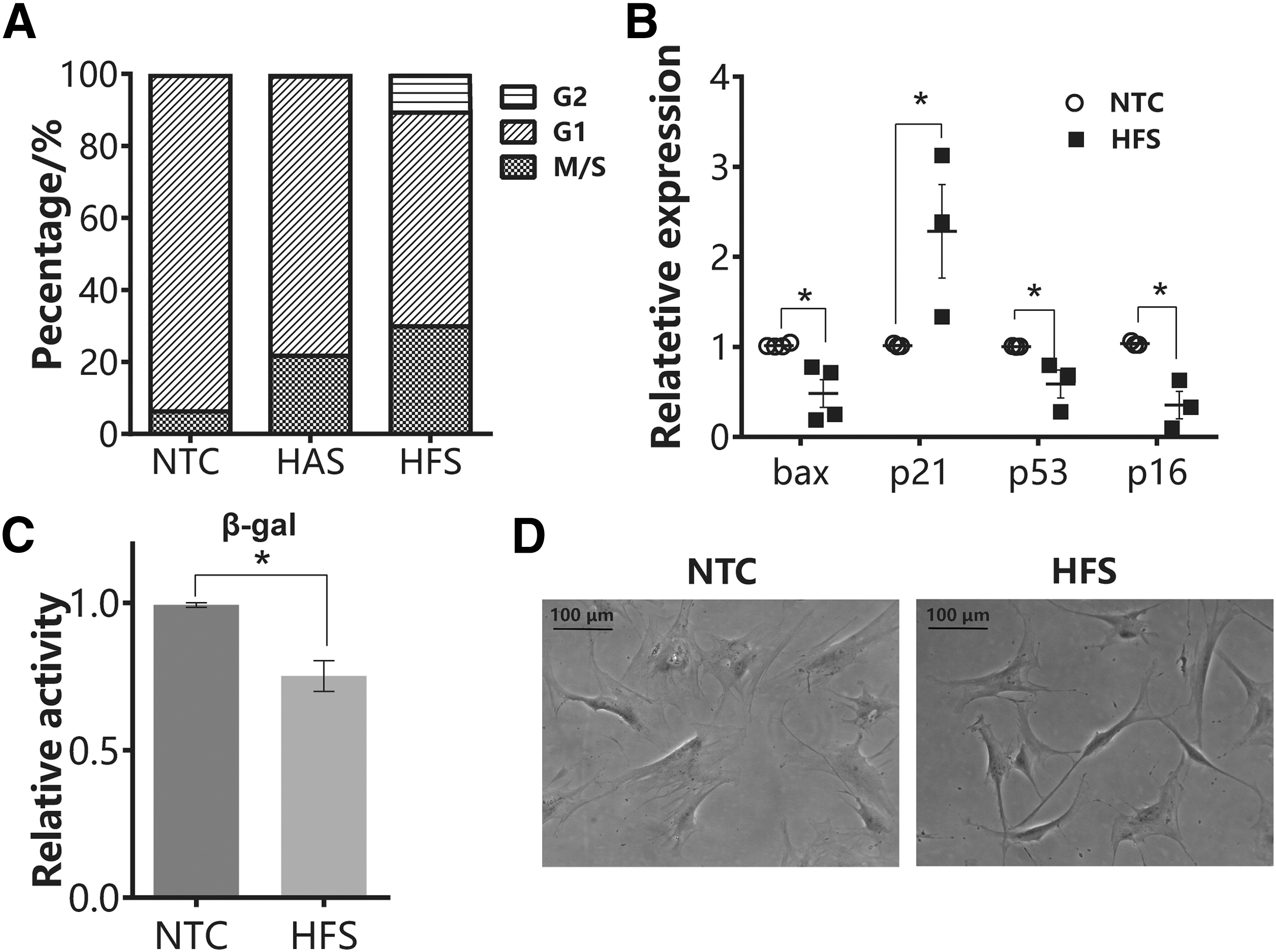
HFS treatment activated cell division and ameliorated SA-β-gal expression and activity. hAMSC were treated with either HFS or vehicle control.
HFS promoted osteogenic potential of hAMSC in vivo
The ultimate goal of MSC therapy is achieved through the injection or implantation of MSC to facilitate regeneration of damaged tissues. Our in vitro data have suggested HFS as a preconditioning alternative to enhance the differentiation ability of MSC. To provide more solid evidence, the aging hAMSC was pretreated with HFS for three consecutive passages before being implanted into nude mice. At 8 weeks post implantation, H&E staining showed that hAMSC (passage 15) had higher tendency to differentiate into adipocyte tissues, while hFMSC (passage 15) exhibited higher osteogenic ability. Pretreatment of HFS reversed the aging property of hAMSC to more hFMSC like as shown by more pinkish fiber matrices and organized collagen fibers (Fig. 5A). The immune histochemistry result of osteogenic-related marker was detected on paraffin sections. The represented picture shows differential expression of COL1A1 and OPG as shown in Fig. 5A. Simultaneously, the semiquantification result, which was measured by Image-pro plus 6, is shown in Fig. 5B. COL1A1 and OPG were highly expressed in HFS group. Col1A1 is usually highly expressed at the early phase of osteoblast differentiation [31].OPG is usually secreted by mature osteoblast to suppress the osteoclast differentiation [32]. Taken together, our immunochemistry result suggests that HFS treatment group is undergoing faster osteogenesis process in vitro than hAMSC group as much higher late-stage marker in osteoblast differentiation, OPG, was detected.
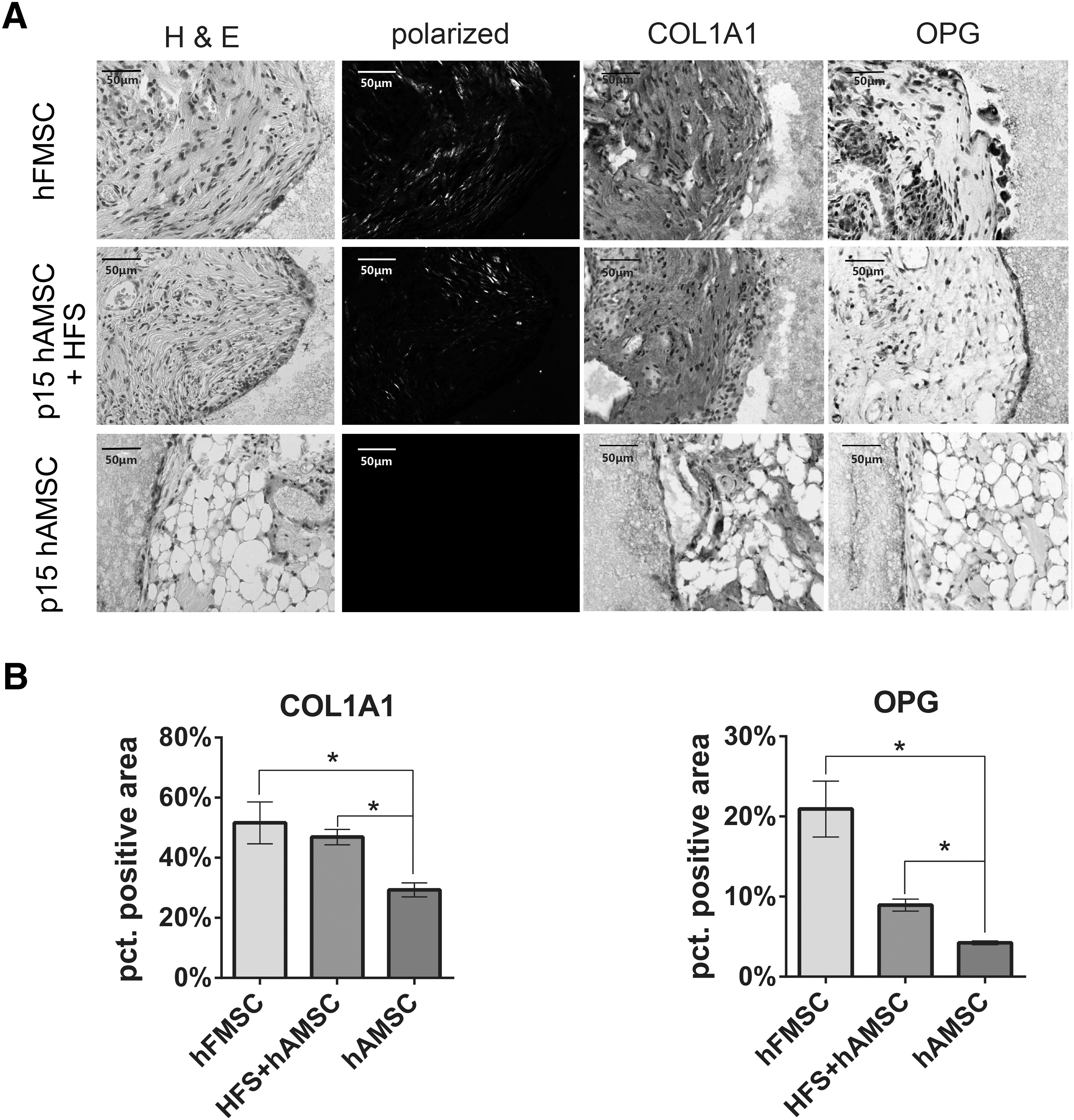
HFS treatment promoted ectopic bone formation of hAMSC in nude mice.
Effects of HFS on tumor growth
There is no doubt that the complex and diverse nature of MSC secretome might result in unknown side effects in addition to beneficial outcomes. Promotion of tumor growth is of particular concern as shown in our previous study [33]. The transduction of luciferase into PC3 allowed longitudinal monitoring of luciferase activity, equivalent to tumor mass, in nude mice (Fig. 6A). In this study, concurrent injection of HFS did not promote tumor growth as indicated by similar luciferase activity between the PC3 group and PC3 plus HFS group throughout the 1-month observation period. Injection of HFS suspended in Matrigel did not result in observable tumor growth (Fig. 6B).
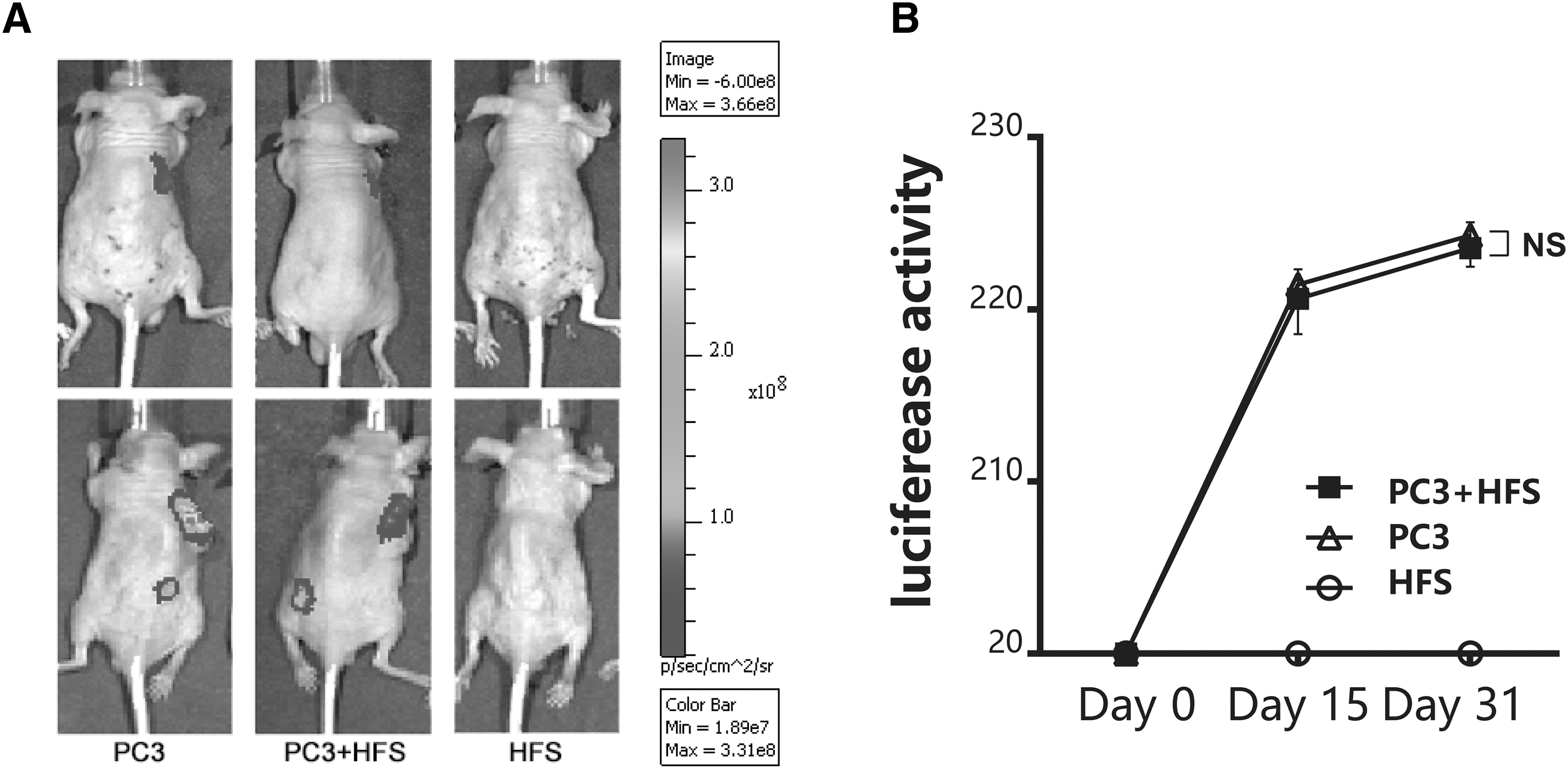
Effect of HFS treatment on tumor growth in vivo. The growth of human prostate tumor cell line (PC3) overexpression luciferase was used to test if HFS promoted tumor growth.
Discussion
Cellular senescence is a basic process, which contributes to age-related dysfunction through low-level chronic sterile inflammation. Recent studies revealed that some age-related dysfunction could be delayed by eliminating the senescent cells [34]. Previous study comparing the senescence progress of MSC isolated from young and aged adult donors showed associated genomic instability and differentiated global gene expression, which were also observed in MSC undergoing continuous in vitro culture irrespective of the source and culture condition [35]. MSC undergoing replicative senescence exhibited different gene expression profile compared with chronologically aged MSC [36], but both senescent scenarios also result in the secretion of SASP, which could affect the activities of neighboring cells. In addition to our previous report to enhance MSC proliferation with low seeding density and hypoxia [22], we in this study demonstrated that HFS treatment could ameliorate replicative senescence of adult MSC, resulting in the promotion of cell proliferation and osteogenic differentiation with no significant associated side effect of promoting tumor growth. Our data suggest the presence of senescence regulators in HFS, which might be involved in the maintenance of fetal MSC activities through autocrine/paracrine action and served as effective preconditioning approach to facilitate the in vitro culture of aged MSC for autologous transplantation.
MSC secreted bioactive factors, which can mimic the regenerative effect obtained with cell transplantation. Current understanding is that these factors, including cytokines and microRNA, are enclosed in extracellular microvesicles with the size ranging from 30 nm to 5 μm. Like adult MSC, MSC derived from fetal tissues such as umbilical cord, Wharton's jelly, and amniotic fluid also secreted bioactive factors, including TGFβ, VEGF, HGF, IGF-1, MMP9, IL-6, and IL-8. [37]. Previous studies showed that fetal MSC exhibited differential gene expression profile, which facilitated myogenic and neurogenic development [38]. Moreover, the extracellular matrices of fetal and young MSC promoted the cell proliferation and osteogenic differentiation ability of adult MSC [39,40].
The composition of MSC secretome is subjected to the type of MSC and culture condition. In this study, the hFMSC and hAMSC from the same type of tissue were cultured under the identical condition, but eliciting different responses. We suspect that the intrinsic difference between the two cell types resulted in different bioactivity of the secretome. Similar phenomenon has been observed in senescent adult MSC, which promoted tumor cell growth through the secretion of IL-6 [41] and induced premature senescent growth arrest through IGFs binding proteins 4 and 7 [42], suggesting the senescence could be one of the driving forces to produce different secretion profile. Interestingly, the HFS could ameliorate replicative senescence by overcoming the effect of hAMSC own secretome, indicating that there are potent factors in HFS, which could be a potential pharmacological intervention for aging process.
Our data showed that HFS promoted cell division as indicated by the increase of hAMSC count at G2/M phase, which was associated with the downregulation of cell cycle inhibitors p16 and p53, but upregulation of p21. Our data are in agreement with previous findings, which showed that cellular senescence was associated with p16 upregulation [43], while p21 was reported to induce cell proliferation [44,45] and was found to be a more unique developmental senescence marker in embryo [46]. Moreover, p21 was coactivated with runx2, a key regulator of osteogenic differentiation [47,48], in MSC undergoing osteogenic differentiation induced by histone deacetylase inhibitor, valproic acid [49]. This might partly explain the concurrent osteogenic differentiation potential induced by HFS. Surprisingly, indirect coculture hFMSC suppressed calcium nodule formation of senescent hAMSC (Supplementary Fig. S2). The coculture model is one of the models for us to study the effects of paracrine factors from hFMSC on the cellular activities of hAMSC. In the coculture study, we cultured the hFMSC and hAMSC in 1:1 ratio, resulting in similar findings as HFS treatment except for the suppression on osteogenic differentiation. The presence of two cells allows dynamic cell-to-cell interaction, but also might interfere with the responses. Meanwhile, in view of the enriched concentration in HFS, it is speculated that the HFS might possess biphasic effect on the osteogenic differentiation subjecting the concentration of that components.
As a member of the sirtuin family of class III histone deacetylases, Sirt1 could prolong lifespan through multiprotective roles against cellular senescence and stress in the neural, cardiovascular, and renal systems [50], thus making it a potential target for antiaging therapy. In musculoskeletal system, sirt1 is also implicated in age-related degenerative diseases such as osteoporosis and osteoarthritis [51,52]. Sirt1 activation promoted MSC proliferation and reduced cellular senescence [30,53] and regulated the swing between osteogenesis and adipogenesis of MSC [54]. Adipogenesis of MSC is strongly associated with cellular aging [55,56]. A number of Sirt1 activators have been developed to target Sirt1 specifically for the treatment of aging and aging-related diseases. Previous study showed that Sirt1 expression in murine MSC decreased with the age of donor increase. Overexpression of sirt1 by genetic approach was shown to ameliorate age-related senescence of MSC [29]. Recently, two epigenetic modifiers, ezh2 and kdm6a, have been found to regulate the swing of osteogenesis and adipogenesis of MSC [57]. In contrast, trimethylation of H3K27 also played an important role in cellular senescence. In our study, the lower expression of p16 may indicate the inactivation of ezh2, whereas the higher expression of runx2 and p21 might suggest involvement of other epigenetic modifiers. Further mechanistic study is warranted to explore how HFS regulates cellular senescence and differentiation. The specific contains of secretome is of utmost importance for explaining the phenomenon. We have performed the ELISA to investigate the possible contains of fetal secretome. According to the literature, we select 3 factors, which may relate with the superior function of HFS as our potential targets (Fig. 7A–C). LIF is an essential factor to maintain the pluripotency of embryonic stem cell [58]. In MSC, LIF is secreted by MSC and mediate the immunosuppression function [16,59]. Although its function on MSC differentiation and proliferation is largely unknown, LIF can prevent senescence in neural stem cell [60]. In our ELISA result, LIF is much more enriched in HFS (Fig. 7A); this indicated that LIF may play an important role in regulation of stemness of MSC. Fetal MSC possess much higher stemness than adult MSC. Lamin B1 (LMNB1) is a well known senescence marker; decreased Lamin B1 expression indicates that cell is undergoing senescence [27], thus we test LMNB1 contains in HFS. Although the result shows no significant difference, we still observed the relative higher value of concentration of LMNB1 in HFS (Fig. 7B). PDGFAA (PDGFA) and PDGFBB are two isoforms of PDGF family. PDGF signaling is believed to promote the osteogenic, as well as proliferation, ability of MSC [61,62]. In our test, nearly fourfold contains of PDGFAA were detected in HFS while comparing with HAS (Fig. 7C). PDGFAA can be secreted by senescent cell through SASP, which inversely contribute to the proliferation and senescence prevention of other cells in wound healing process [17]. In summary, ELISA results provide us possible explanation to the benefit of HFS. Higher level of PDGFA and LMNB1 may partially contribute to the antisenescence function of HFS. Meanwhile, the amount of LIF in HFS is much higher than HAS. Although we did not find direct evidence to support the antiaging effect of LIF on MSC, it is very interesting that LIF may also contribute to the antisenescence property, since LIF is a well-known growth factor to maintain stemness of embryonic stem cell.
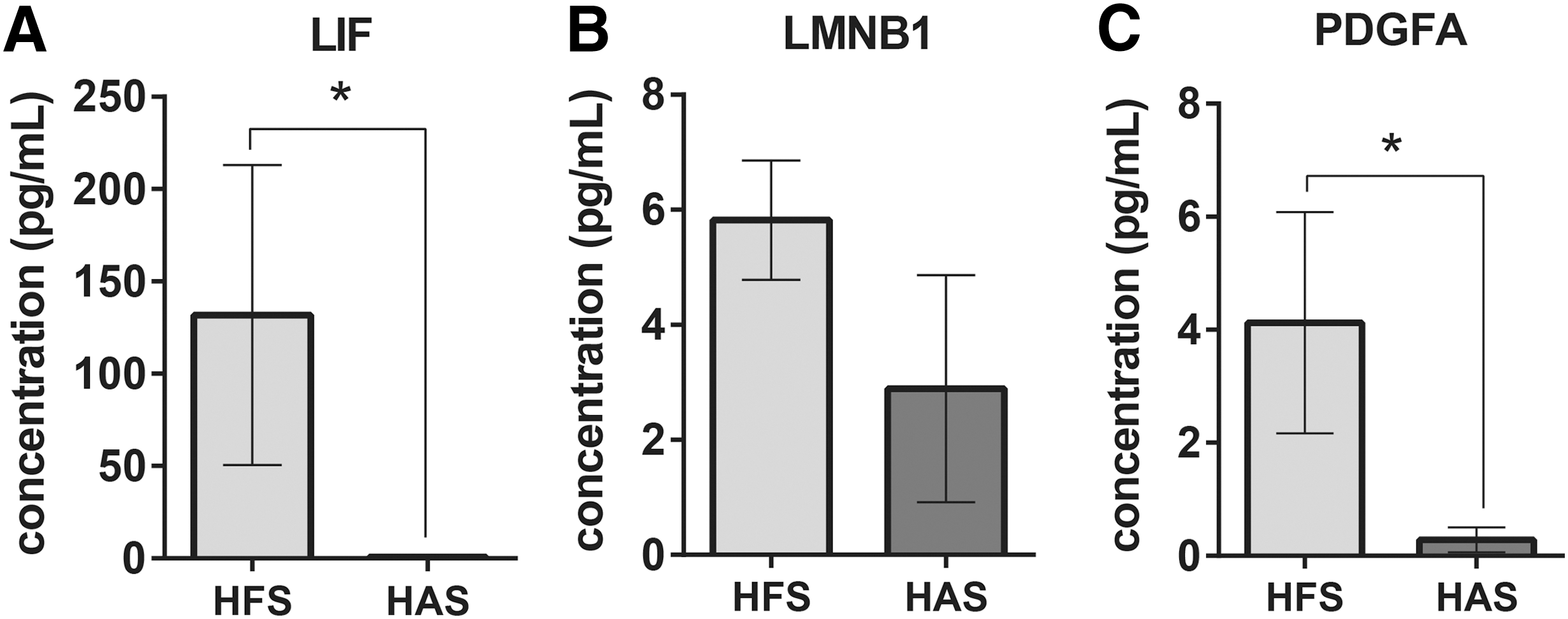
Differentiated secretion of bioactive factors in HFS and HAS. HFS consists of more
This study is largely limited by the poor understanding on the proteomic profile of adult and fetal MSC secretome, which have not been characterized yet. There is emerging evidence suggesting that the therapeutic effects of MSC therapy could be partly achieved by MSC secretome, which renders diverse biological activities, including immunomodulation, anti-inflammatory activity, and angiogenesis. [63]. However, the composition and bioactivity of MSC secretome vary with the source of MSC, culture condition for MSC, and method of secretome preparation; therefore, the control of desired factor production and release and balance of positive and negative factors remain the outstanding challenge to overcome. High throughput proteomic method is warranted to identify the composition of HFS versus HAS and to identify and validate more potent antisenescence factors in the HFS. Physiological, genetic, pharmacological, and cell–cell interaction are common preconditioning approaches, which have been tested to enhance the bioactivity of MSC secretome for the treatment of cardiovascular disease [64]. To serve as a complement to existing understanding on MSC secretome which is mainly obtained from study on adult MSC, our data on the comparison between adult and fetal MSC secretomes provided the first ever evidence showing the beneficial effect of fetal MSC secretome on cellular senescence.
Footnotes
Acknowledgments
The authors thank Li Ka Shing Institute of Health Science, The Chinese University of Hong Kong, for providing the use of clean room human cell culture facilities and Dr. Nina Chu for help in specimen collection. The work was partially supported by grants from National Natural Science Foundation of China (NSFC no. 81371946, 81272548, 81374568, 31400834) to Gang Li, Xiao-hua Jiang, and Liangliang Xu, respectively. Hong Kong Government Research Grant Council, General Research Fund (CUHK470813 and 14119115), and a project grant from China Shenzhen City Science and Technology Bureau (JGJHZ20140419120051680) to Gang Li are gratefully acknowledged. This study was also partly supported by SMART program, Lui Che Woo Institute of Innovative Medicine, The Chinese University of Hong Kong.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.