
Abstract
Pluripotent stem cells act as an excellent cell source for disease therapy because of its specific characteristics of self-renewal and differentiation. Pluripotent stem cells are heterogeneous, consisting of naive stem cells as well as primed epiblast stem cells. However, the strategies and mechanisms of maintaining naive pluripotent stem cells remain unclear. In this study, we found that folic acid (FA) sustained mouse embryonic stem cell (ESC) pluripotency and enabled long-term maintenance of the naive state of ESCs under CHIR99021 conditions. Mechanistic experiments showed that STAT3 pathway partially mediated the effect of FA after which the interaction between STAT3 and importin α5 was enhanced. Meanwhile, MEK/ERK signaling also acted downstream of FA in maintaining ESC pluripotency. Furthermore, FA significantly promoted mouse somatic cell reprogramming. Overall, our study identified an effective chemical condition for maintaining homogeneous ESCs and highlighted the important roles of LIF/STAT3 and MEK/ERK signaling in naive ESC pluripotency.
Introduction
S
Interestingly, somatic cells can be reprogrammed to become pluripotent via nuclear transfer with oocytes or through the ectopic expression of defined factors [6 –8]. Induced pluripotent stem (iPS) cells generated by ectopically expressing defined factors exhibit the same properties as embryonic stem cells (ESCs), including morphology, proliferation, epigenetic status, and differentiation potential. However, previous reprogramming strategies have raised concerns regarding their clinical applications [9]. Strikingly, chemical-iPS cells have tended to meet the needs of regenerative medicine, highlighting the important roles of small molecules in reprogramming [10,11]. However, the time course of the presently reported chemical-induced reprogramming procedure is long and the efficiency is low; further exploration of the role of small molecules in pluripotency and reprogramming is of great interest.
Pluripotency, or the ability of a single cell to give rise to all cell types in an entire organism, attracts great biological interest in terms of understanding developmental mechanisms as well as the medical potential of pluripotent stem cells [12]. During early mouse development, successive differentiation events lead first to the segregation of the trophectoderm, and second, to the hypoblast on the blastocoelic surface of the inner cell mass (ICM), from which ESCs derive [13,14]. ESC differentiation in vitro faithfully mimics early embryonic development; the systematic study of ESC differentiation provides opportunities to deeply understand the cell biology of early development. Interestingly, folic acid (FA), also known as vitamin B9, has been shown to play a critical role during early embryonic development [15 –17], suggesting that FA may participate in regulating ESC pluripotency.
ESCs are characterized by three distinguishing features, pluripotency, self-renewal (maintenance of an undifferentiated state), and limitless proliferation, which can be maintained, in part, by the cytokine leukemia inhibitory factor (LIF) in ESCs [18,19]. Moreover, the ground state of ESCs reflected in the homogeneity of ESC populations can be sustained in the presence of 2i (CHIR99021, an inhibitor of glycogen synthase kinase-3β; PD0325901, an inhibitor of MEK/ERK) and LIF [20,21]. In addition, chemical combinations such as 2i were reported to act as a candidate culture condition to promote iPS cell reprogramming [22].
In the present study, we demonstrated that FA supports robust long-term ESC pluripotency under CHIR99021 conditions. FA maintains ESC pluripotency by activating the LIF/STAT3 pathway and inhibiting ERK signaling. Furthermore, FA significantly promotes somatic cell reprogramming. Our findings reveal a new set of culture condition for maintaining ESC pluripotency and facilitating reprogramming.
Materials and Methods
Animals
Male nonobese diabetic/severe combined immunodeficient (NOD-SCID) mice (4 weeks of age) were purchased from the Shanghai SLAC laboratory Animal Co., Ltd. (Shanghai, China). All procedures were performed according to protocols approved by the Tongji University Institutional Animal Care and Use Committee.
ESC culture and generation of iPS cells
All mouse ESCs were cultured in Gibco-DMEM [15% FBS (Gibco, NY), 1 × penicillin/streptomycin solution (P/S) (Hyclone), 1 × nonessential amino acids (NEAA) (Thermo), 1 ×
In vitro ESC differentiation and teratoma formation
To form EBs, ESCs were trypsinized into single cells and differentiated in KOSR medium without LIF and other chemical compounds using the hanging-drop method. Single ESCs were cultured at 1 × 103 cells/20 μL in a hanging drop for 3 days. EBs were collected onto gelatin-coated 48-well plates and cultured for another 6 days. For teratoma assay, ESCs were trypsinized, suspended in a medium without LIF and other chemical compounds, and injected into NOD-SCID mice at a concentration of 2 × 106 cells/150 μL. Following injection, tumor formation was examined every week for up to 3–4 weeks. Tumors were harvested and fixed in a formaldehyde solution before paraffin embedding and subsequent HE staining.
Quantitative real-time polymerase chain reaction
RNA was extracted using the TRIzol Reagent (Qiagen) according to the manufacturer's instructions. Quantitative real-time polymerase chain reaction (qRT-PCR) analyses were performed on an MX3000P Stratagene PCR machine with iTaq™ Universal SYBR Green Supermix (BIO-RAD). Relative gene expression was normalized to endogenous Actin. Primer sequences are listed in Supplementary Table S1 (Supplementary Data are available online at
Western blot and immunostaining
Cell lysates were separated by SDS-PAGE and transferred onto nitrocellulose membranes. The membranes were probed with the following primary antibodies: P-ERK1/2 (1:1,000; Cell Signaling), ERK1/2 (1:1,000; Santa Cruz), p-STAT3 (1:2,000; Sigma), STAT3 (1:1,000; Cell Signaling), PIM1 (1:1,000; Cell Signaling), TFCP2L1 (1:1,000; Sigma), TUBULIN (1:1,000; Cell Signaling), and TBX3 (1:1,000; Cell Signaling). Membranes were washed three times and then incubated with secondary antibodies (1:5,000; Cell Signaling) for 1 h at room temperature. Signals were visualized by enhanced chemiluminescence (ImageQuant LAS 4000 mini). Immunostaining was carried out as described [23]. The antibodies used in immunostaining were as follows: Nanog (1:1,000; Abcam) and SSEA1 (1:500; Santa Cruz).
Clonogenicity assay
ESCs were seeded on gelatin-coated tissue culture dishes at 600 cells per well of six-well plates. Cells were cultured in LIF medium for 6 days. After culturing for 6 days, the cells were fixed and then stained for AP activity according to the manufacturer's protocol (Sigma). The stained colonies were counted and imaged.
Statistical analysis
Data are presented as mean ± standard error of the mean of three independent experiments. Statistical comparisons were performed using unpaired two-tailed Student's t tests. *P < 0.05, **P < 0.01, ***P < 0.001.
Results
FA sustains ESC pluripotency and self-renewal for several passages
To explore the effect of FA on ESC pluripotency, we dosed ESCs with FA in the absence of LIF for 48 h. Consistent with a previous report [18], ESCs adopted a fibroblast morphology after LIF withdrawal (Fig. 1A and Supplementary Fig. S1A). Interestingly, FA significantly inhibited ESC differentiation and restored the morphology of undifferentiated ESC colonies in a dose-dependent manner (Fig. 1A and Supplementary Fig. S1A). However, high concentration of FA treatment actually decreased undifferentiated colony formation by male ESCs (Fig. 1A, B) and female ESCs (Supplementary Fig. S1A, B). To test whether high-concentration FA would affect cell propagation, the 80,000 ESCs or MEFs were seeded and cultured for 48 h, and then, the cell numbers were counted. Our data showed that high concentration of FA, 3.0 mM, significantly inhibited the cell proliferation in two cell lines, probably due to the toxicity (Supplementary Fig. S2A, B). qRT-PCR analyses showed that FA caused a dose-dependent increase in the expression levels of the naive pluripotency markers Nanog, Oct4, Sox2, Esrrb, Rex1, Klf4, and c-Myc (Fig. 1C and Supplementary Fig. S1C). Folate receptor alpha (FRα) has been demonstrated that acted as a transcription factor regulates the expression by trans locating to the nucleus. Recently, a study found that FRα can upregulate Oct4, Sox2, and Klf4 in cranial neural crest cells [24,25]. Moreover, colony formation assays revealed that 1 mM FA notably increased the number of undifferentiated colonies to the level of LIF treatment (Fig. 1D). These data suggested that FA plays a critical role in maintaining ESC pluripotency and self-renewal.
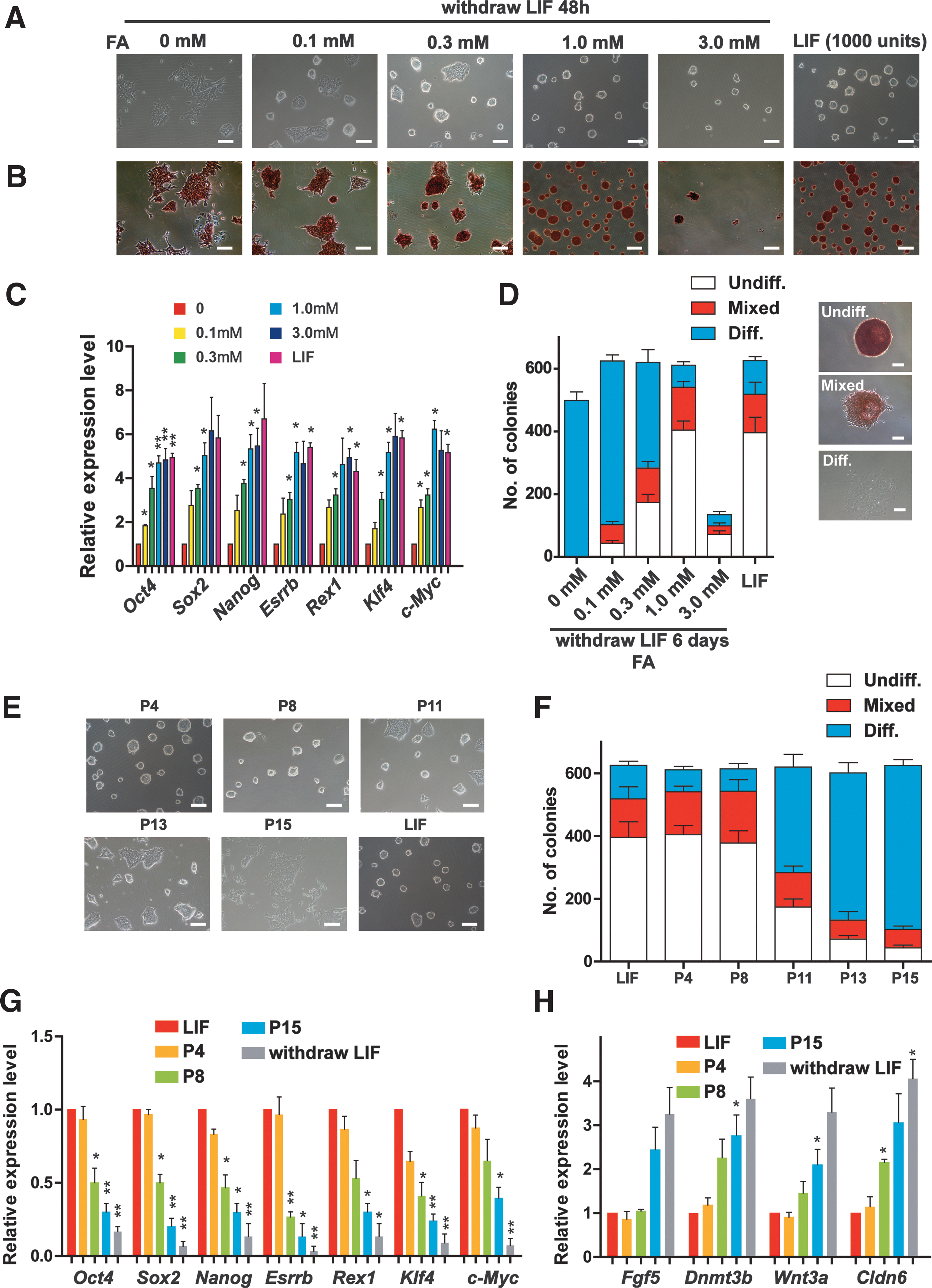
Folic acid maintains ESC pluripotency and self-renewal for several passages.
Next, we determine how long FA can maintain ESC pluripotency and self-renewal. ESCs treated with FA effectively retained the morphology of undifferentiated ESC colonies and the capacity for clonogenicity for eight passages (almost 20 days), whereas ESCs older than eight passages gradually lost these properties (Fig. 1E, F, and Supplementary Fig. S1D). On the contrary, the expression levels of naive pluripotency genes Nanog, Oct4, Sox2, Esrrb, Rex1, Klf4, and c-Myc in ESCs treated with FA gradually decreased with increasing passage number (Fig. 1G), while the levels of the differential epiblast marker genes Fgf5, Dnmt3b, Wnt3a, and Cldn6 were notably upregulated in ESCs older than 8 passages (Fig. 1H), indicating that high-passage ESCs treated with FA partially differentiated. X chromosome inactivation (XCI) has been shown to be accompanied with the differentiation of female ESCs [26,27], indicating that XCI is incompatible with the ground pluripotent state of ESCs. Importantly, FA maintained the expression of X-linked genes (Xist and Jpx) and Tsix (transcribed antisense to Xist) to similar levels as LIF treatment (Supplementary Fig. S1E), suggesting that FA inhibited XCI. Collectively, these data showed that FA maintained ESC pluripotency and self-renewal for several passages.
FA facilitates somatic cell reprogramming
Somatic cell reprogramming is accompanied by the acquisition of pluripotency and eventually leads to the generation of iPS cells [7,28]. Based on our results, we postulated that FA might play a role in iPS cell generation. To generate iPS cells, four factors (OSKM; Oct4, Sox2, Klf4, and c-Myc) were introduced into Oct4-GFP MEF cells. Compared with the control group, as expected, FA treatment resulted in a twofold increase in the numbers of AP- and Oct4-GFP-positive colonies (Fig. 2A–D). Importantly, compared with the FA group, combining FA and CHIR99021 caused a fourfold increase in the number of AP-positive colonies, or a threefold increase in the number of GFP-positive colonies (Fig. 2E–H). These results suggested that FA also played important roles in iPS cell generation.

FA promotes somatic cell reprogramming.
Next, we characterized the properties of the iPS cell lines induced by the four factors plus FA and CHIR99021, which we named as CF-iPS cells. As with normal iPS cells (LIF-iPS cells), CF-iPS cells exhibited the typical ES-like morphology and expressed high levels of the pluripotency markers Nanog, Oct4, and SSEA1 (Fig. 2I and Supplementary Fig. S2C). In vivo teratoma assay also confirmed the differentiation ability of the CF-iPS cells (Fig. 2J). Collectively, these data indicated that FA enhances reprogramming efficiency.
FA maintains ESC pluripotency via LIF/STAT3 and MAPK/ERK signaling
Next, we elucidated the downstream signaling of FA. The LIF/STAT3 and MAPK/ERK pathways have been demonstrated to be critical in maintaining ESC pluripotency and self-renewal as well as reprogramming [29 –35]. STAT3 is a core component of the LIF pathway, and phosphorylated STAT3 translocates to the nucleus, where it subsequently regulates the expression of target genes such as Klfs [36 –40]. Based on our previous results in which FA maintained ESC pluripotency and self-renewal after LIF deprivation, we hypothesized that FA stimulated the LIF/STAT3 pathway. Gene set enrichment analysis (GSEA) showed that FA activated JAK/STAT3 signaling (Fig. 3A). Although FA did not lead to a time-dependent increase in JAK expression (Supplementary Fig. S3A) and STAT3 phosphorylation (Fig. 3B), FA treatment significantly decreased STAT3 phosphorylation levels in the cytoplasm and upregulated the levels of phosphorylated STAT3 in the nucleus (Fig. 3C), suggesting that FA might promote the transcriptional activity of STAT3 by relocalizing p-STAT3 in the nucleus. Then, the expression of importin proteins, which are critically involved in STAT3 relocalization in the nucleus, was examined by western blotting assay. Interestingly, we found that the expression of importin α5 was upregulated after FA treatment in the absence of LIF (Fig. 3D). We also found that FA treatment could enhance the interaction between STAT3 and importin α5, which plays important roles in STAT3 relocalization in the nucleus (Fig. 3E). qRT-PCR analyses demonstrated that FA significantly enhanced the expression of downstream transcription factors of the JAK-STAT3 pathway, namely, Pim1, Tfcp2l1, Klf2, and Klf5, but not Tbx3, Gbx2, and Socx3 (Fig. 3F). Moreover, ChIP analyses revealed that FA treatment did promote the binding of STAT3 to the promoters of STAT3 target genes such as Pim1, Tfcp2l1, Klf2, and Klf5, but not Tbx3, Gbx2, and Socx3 (Fig. 3G). Western blotting showed that FA increased PIM1 and TFCP2L1 protein levels (Fig. 3H). Previous study reported that a downstream target of STAT3 and TFCP2L1 can replace PD0325901 in maintaining ESC self-renewal [41]. Next, we explored the function of the MEK/ERK pathway in FA signaling. In addition, FA regulated the MEK/ERK pathway, as revealed by GSEA (Fig. 3I). However, FA did not lead to a time-dependent decrease in Raf or Ras expression (Supplementary Fig. S3B). Interestingly, similar to the effect of the MEK/ERK1/2 inhibitor PD0325901, FA dramatically reduced the phosphorylation of ERK1/2 (Fig. 3J). These results indicated that FA controls pluripotency by modulating the LIF/STAT3 and MEK/ERK pathways in ESCs.
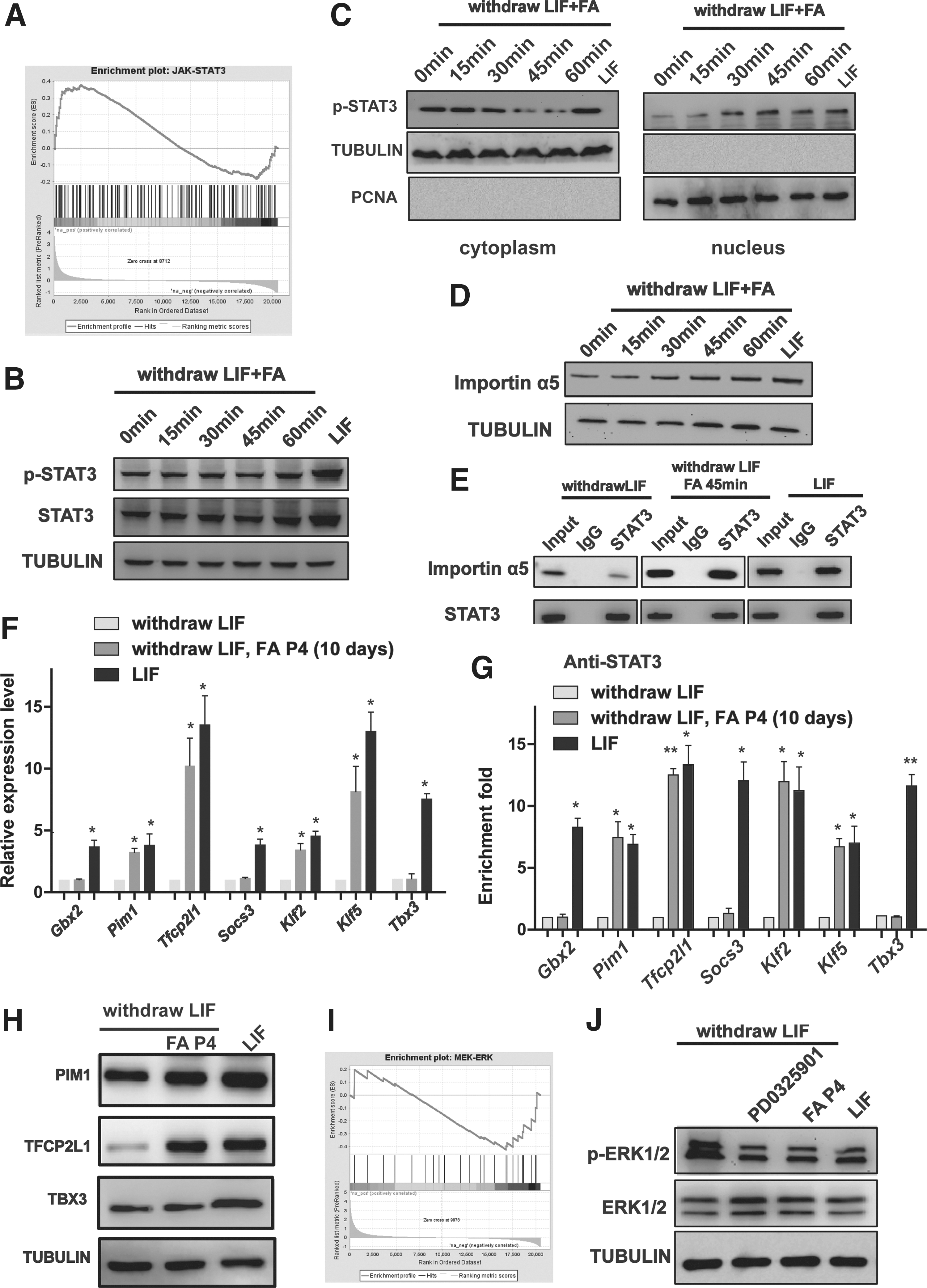
FA maintains the pluripotency of ESCs through the LIF/STAT3 and MAPK/ERK signaling pathways.
Small-molecule inhibitors rescue the effect of FA on pluripotency in ESCs
Previously, STAT3 phosphorylation is described to play critical roles in ESC pluripotency. Since the level of phosphorylated STAT3 in the nucleus was increased by FA treatment at the absence of LIF, we next tried to investigate whether the chemical inhibiting STAT3 phosphorylation can restore FA-maintained ESC pluripotency. The ESCs were treated with S31-201, which inhibits the STAT3 transcription factor by blocking the phosphorylation and dimerization events necessary for activation. Interestingly, ES-like morphology maintained by FA was suppressed by the inhibition of STAT3 activity (S31-201, 86 μM; Fig. 4A). The high expression levels of pluripotency markers were significantly suppressed by STAT3 inhibition, conversely, the low expression levels of differential epiblast marker genes slightly increased (Fig. 4B). Moreover, STAT3 inhibition abolished the effect of FA on X chromosome activation in NR3 cells (Fig. 4B). The capacity for clonogenicity was also rescued (Fig. 4C). In addition, MEK/ERK1/2 activation by insulin treatment (200 nM; Supplementary Fig. S3C) [42] was also clearly abrogated by the influence of FA in maintaining ESC pluripotency and self-renewal (Fig. 4D, E). Importantly, the combination of the STAT3 inhibition (S31-201, 86 μM) and MEK/ERK activation (insulin, 200 nM) completely abolished the effect of FA on ESC pluripotency (Fig. 4F–H). Overall, these data confirm the involvement of both the LIF/STAT3 and MEK/ERK pathways in FA function in ESCs.
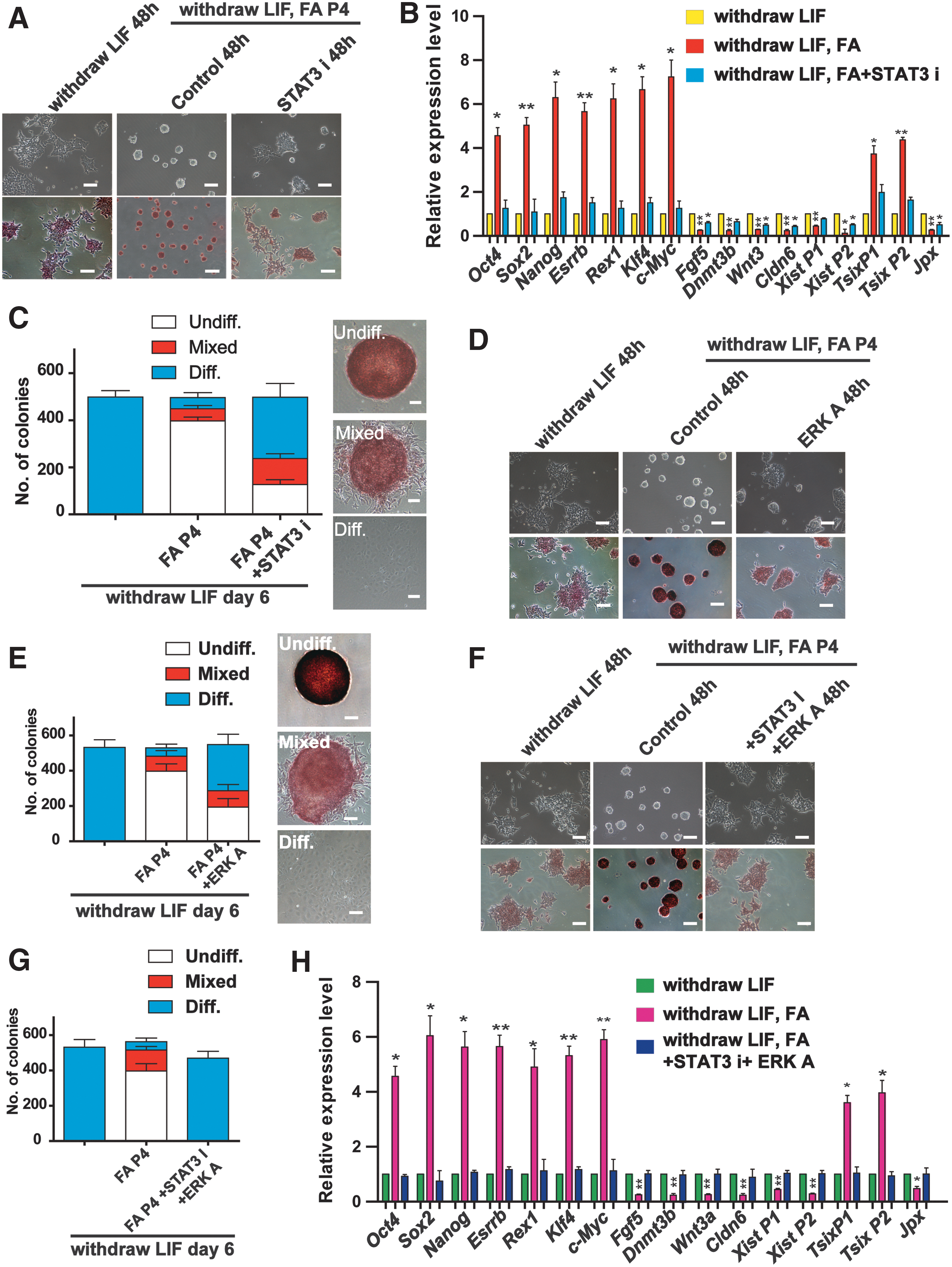
Small-molecule inhibitors rescue the effect of FA on pluripotency in ESCs.
The combination of FA and CHIR99021 promotes the ground state pluripotency
Because combining the GSK-3β inhibitor CHIR99021 with the MEK/ERK inhibitor PD0325901 (2i) is sufficient for the expansion of ESCs [21,35], we next tested whether combining of FA with either CHIR99021 or PD0325901 can support the long-term expansion of ESCs. The combination of FA with CHIR99021, but not with PD0325901, supported undifferentiated ESC expansion (Supplementary Fig. S3D) and increased the expression of TBX3 (Supplementary Fig. S3E), a critical transcriptional factor for maintaining ESC pluripotency. Interestingly, combining FA and CHIR99021 maintained the morphology of undifferentiated ESC colonies and strong AP activity of ESCs, even after 20 passages (almost 50 days), whereas CHIR99021 treatment alone had no effect on promoting ESC pluripotency (Fig. 5A). GSEA showed that FA did not affect WNT signaling (Supplementary Fig. S3F). ESCs treated with both FA and CHIR99021 had similar gene expression patterns as those of ESCs cultured under LIF condition or 2i condition, and combining of FA and CHIR99021 dramatically induced the expression of pluripotency genes Nanog, Oct4, Sox2, Esrrb, Rex1, Klf4, and c-Myc to the same levels as ESCs treated with 2i at high passages (Fig. 5B and Supplementary Fig. S4A) and stably reversed the capacity of clonogenicity (Fig. 5C and Supplementary Fig. S4B). Meanwhile, ESCs treated with FA and CHIR99021 expressed low levels of differential epiblast markers (Supplementary Fig. S4C). Next, we evaluated the differentiation potential of ESCs treated with FA and CHIR99021. ESCs treated with FA and CHIR99021 showed very similar expression patterns of three germ layer genes as LIF ESCs during in vitro embryoid body differentiation assays (Fig. 5D). In addition, we also observed all three germ layers in teratomas derived from ESCs treated with FA and CHIR99021 (Fig. 5E). Furthermore, we found that the size and weight of teratomas derived from CF ESCs were similar to those of LIF ESC-derived teratomas (Supplementary Fig. S5A, B). Moreover, the teratomas derived from CF ESCs showed very similar expression patterns of three germ layer genes as LIF ESC-derived teratomas (Supplementary Fig. S5C–E). Thus, these results revealed that ESCs treated with FA and CHIR99021 have a normal differentiation capacity in vitro and in vivo.
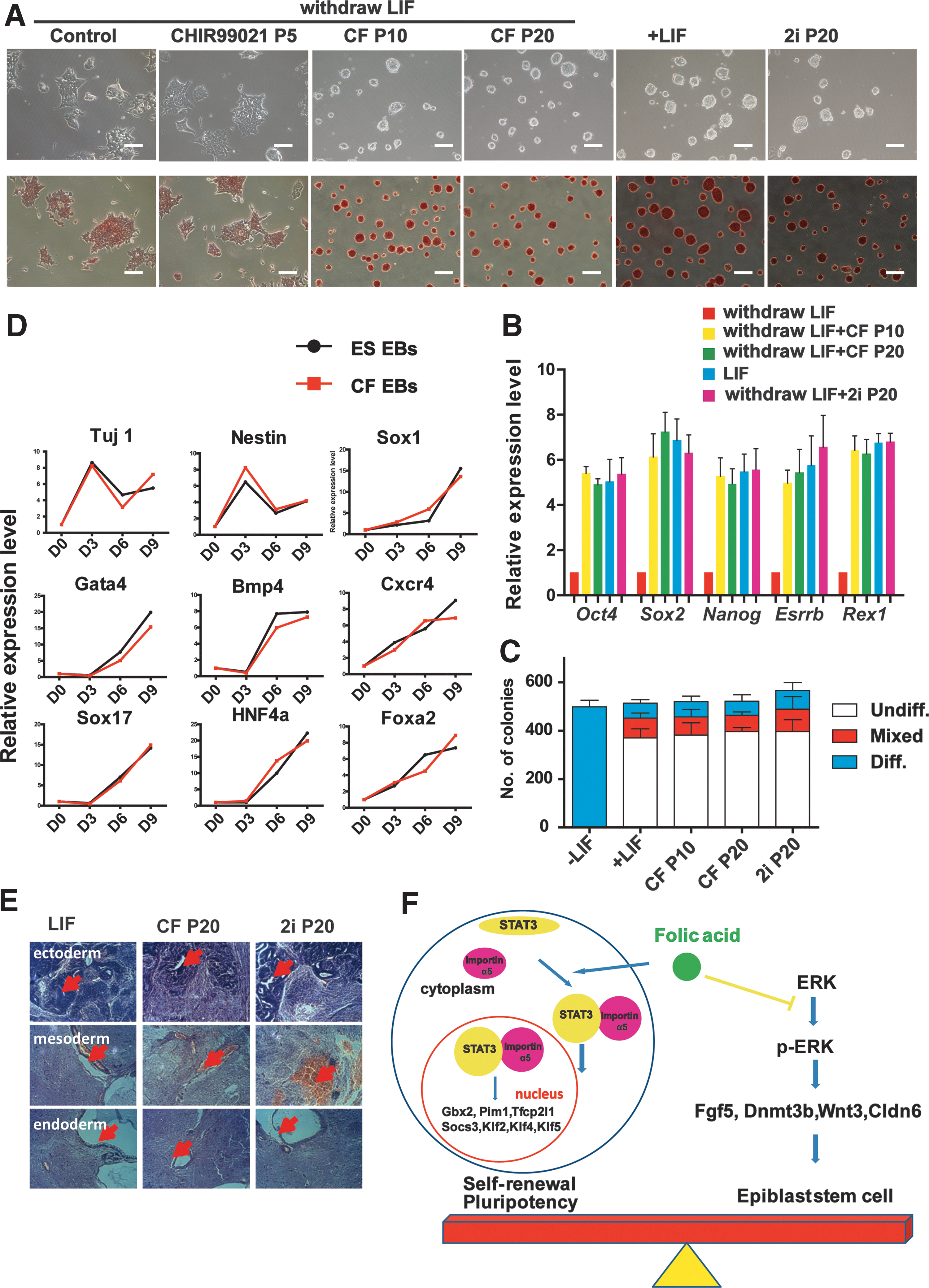
The combination of FA and CHIR99021 promotes ground-state pluripotency in ESCs.
Then, we investigated the influence of FA plus CHIR99021 on XCI. Consistent with LIF treatment, the combination of FA and CHIR99021 sustained low expression levels of X-linked genes (Xist and Jpx) and high expression levels of their transcribed antisense sequence Tsix in female NR3 ESCs (Supplementary Fig. S4D), suggesting that combining FA and CHIR99021 significantly inhibited XCI. Taken together, our results demonstrated that FA maintained the long-term pluripotency and self-renewal of ESCs under CHIR99021 conditions.
Discussion
Pluripotent stem cells provide a promising cell resource for studying disease and for regenerative medicine [2,43]. Self-renewal and differentiation are two naturally opposite and equally important characteristics of these cells. Maintaining pluripotency and self-renewal using chemicals is of great importance for the amplification, scientific study, and potential clinical application of pluripotent stem cells. In the present study, we found that FA, especially when combined with CHIR99021, efficiently maintained ESC pluripotency and self-renewal by activating STAT3 targeting genes and inhibiting MEK/ERK signaling.
ESCs display two types of pluripotent states, naive and primed epiblast, between which they can interconvert [44]. Naive ESCs appear to have the largest role in future applications [45,46]. However, a list of mouse and human ESCs cultured in vitro do not display the naive pluripotent state and maintaining the naive pluripotent state of ESCs remains to be achieved. Identifying a chemical approach to maintaining the naive state of ESCs is a worthwhile strategy. In addition, because the efficiency of currently available chemically induced reprogramming is low, optimizing the chemical combinations used to induce reprogramming is extremely useful for mechanistic studies and the clinical application of iPS cells. FA induced multiple functions while simultaneously avoiding the cytokine-induced cross-crossing physiological processes of cultured ESCs, owing to its properties as a small molecular compound. Previous data reported that, under LIF conditions, transient folate deprivation facilitates the generation of somatic cell-derived pluripotent stem cells in mice [47]; our data suggested that, in the absence of LIF, combining of FA and CHIR99021 may be a candidate chemical condition for maintaining the naive state of ESCs and enhancing iPS cell generation.
Stem cell populations do not comprise a homogenous cellular entity and instead exhibit an intrinsic molecular heterogeneity [20,48 –50]. Several signaling pathways, including the LIF/STAT3, MEK/ERK, and GSK-3β pathways, are known to play critical roles in the induction and maintenance of the naive pluripotent state of ESCs [13,36,38,51,52]. LIF plus 2i can efficiently maintain naive ESCs by regulating the LIF/STAT3, MEK/ERK, and GSK-3β pathways [20,21]. In the present study, FA not only partially activated downstream genes of the LIF/STAT3 pathway but also inhibited ERK1/2 phosphorylation. Importantly, FA in combination with CHIR99021 stably maintained the naive state of ESCs in the absence of LIF, suggesting that FA plus CHIR99021 may substitute for the combination of LIF and 2i in maintaining naive ESCs.
The pluripotency network is inherently stable but is extremely sensitive to destabilization by exogenous signals such as MEK/ERK signaling [53]. ERK signaling is strongly activated during the lineage-specific differentiation of pluripotent stem cells and is critically associated with the initiation of trophectoderm differentiation. Our data revealed that FA inhibited the MEK/ERK pathway, suppressed differentiation initiation, and sustained naive ESCs. In addition, FA stabilized the active state of X chromosomes and blocked differentiation under specific conditions, guaranteeing the homogeneously pluripotent state of female ESCs.
Previously, it was reported that the competence of embryonic development could be affected by various factors, including the pluripotency of ICM cells and their subsequent differentiation into all cell types in the entire organism. Maternal FA intake during pregnancy is known to be significantly associated with a decreased risk of neural tube defects (NTDs) and other FA-sensitive congenital malformations [15 –17,54]. These studies strongly support our results that FA supported robust pluripotency and self-renewal in ESCs derived from ICM under specific conditions. The regulation of ESC pluripotency by FA might also be one of the mechanisms through which FA deficiency leads to NTDs and other birth defects.
In this work, we demonstrated that FA functions as a positive factor in the naive pluripotent state of ESCs by regulating the LIF/STAT3 and MEK/ERK pathways and that FA facilitated mouse somatic reprogramming (Fig. 5F). Our findings deepen our understanding of ESC naive pluripotency and reprogramming and offer a new chemical approach for naive pluripotency maintenance and iPS cell generation, which may help to improve the success of pluripotent stem cells in clinical applications. In addition, these data provide one possible mechanism for FA intake during pregnancy, which is very useful for both prenatal and postnatal care.
Footnotes
Acknowledgments
This work was supported by grants obtained from the Ministry of Science and Technology (grant numbers 2013CB967401, 2012CB966603, 2013CB967600), the National Natural Science Foundation of China (grant numbers 81530042, 31571529, 91519320, 31210103905, 31371510, 31301208, 31471250, 31401257, 31540037), and sponsored by Shanghai Rising-Star Program (14QA1403900). We thank Shaorong Gao's laboratory for providing us NR3 female ESCs.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.