
Abstract
The development of a mammalian brain is a complex and long-lasting process. Not surprisingly, preterm birth is the leading cause of death in newborns and children. Advances in perinatal care reduced mortality, but morbidity still represents a major burden. New therapeutic approaches are thus desperately needed. Given that mesenchymal stem/stromal cells (MSCs) emerged as a promising candidate for cell therapy, we transplanted MSCs derived from the Wharton's Jelly (WJ-MSCs) to reduce the burden of immature brain injury in a murine animal model. WJ-MSCs transplantation resulted in protective activity characterized by reduced myelin loss and astroglial activation. WJ-MSCs improved locomotor behavior as well. To address the underlying mechanisms, we tested the key regulators of responses to DNA-damaging agents, such as cyclic AMP-dependent protein kinase/calcium-dependent protein kinase (PKA/PKC), cyclin-dependent kinase (CDK), ataxia-telangiectasia-mutated/ATM- and Rad3-related (ATM/ATR) substrates, protein kinase B (Akt), and 14-3-3 binding protein partners. We characterized WJ-MSCs using a specific profiler polymerase chain reaction array. We provide evidence that WJ-MSCs target pivotal regulators of the cell fate such as CDK/14-3-3/Akt signaling. We identified leukemia inhibitory factor as a potential candidate of WJ-MSCs' induced modifications as well. We hypothesize that WJ-MSCs may exert adaptive responses depending on the type of injury they are facing, making them prominent candidates for cell therapy in perinatal injuries.
Introduction
P
In immature infants at risk, no individual protective agent has proven safe and effective so far [7 –9]. Magnesium sulfate reduces the incidence of CP in the preterm population, but identification of pregnant mothers with anticipated preterm labor is difficult [10]. New therapeutic approaches are thus desperately needed. In recent years, mesenchymal stem/stromal cells (MSCs) emerged as a promising candidate for cell therapy targeted against perinatal brain injury [11]. MSCs isolated from the stroma of the umbilical cord (Wharton's Jelly [WJ]-MSCs) [12,13] are of special interest as they are easy available around the time of birth, an autologous source, and pose no ethical concerns [14,15]. In addition, WJ-MSCs possess a low immunogenic capacity and a multipotent differentiation potential [13]. MSCs were shown to modulate innate and adaptive immune responses, to have antiapoptotic effects, to decrease inflammation, and to enhance tissue repair, mostly through the release of paracrine factors [16,17].
The development of a mammalian brain is a complex and long-lasting process, which follows specific time- and space-dependent sequences with interactions of various cell types. One of the common denominators is the dynamic equilibrium of cellular proliferation/differentiation and apoptosis/necrosis, which is especially evident in responses to aggressors in the immature brain. Not surprisingly, both inflammation and hypoxia ischemia result in the predilection for white matter injury due to aggressors ranging from oxidative stress to excitotoxicity [14,18]. Generally, in response to DNA damage, the cell cycle checkpoints' activation at the G1/S and G2/M transitions may stop cell cycle progression to repair the damaged genetic material. In case of irreparable DNA damage, the cells will induce apoptosis [19]. The key regulators of responses to DNA damaging agents are cyclic AMP-dependent protein kinase/calcium-dependent protein kinase (PKA/PKC), cyclin-dependent kinase (CDK), ataxia-telangiectasia-mutated/ATM- and Rad3-related (ATM/ATR) substrates, protein kinase B (Akt), and 14-3-3 binding protein partners [20 –26]. For example, activation of PKA/PKC signaling results in the protection of an immature brain [21] and CDK inhibition proved to be protective in stroke [19]. Activation of ATM/ATR signaling results in neuronal apoptosis [26], and Akt signaling is a well-defined prosurvival pathway [20]. Furthermore, 14-3-3 proteins have been shown to be neuroprotective in a variety of neurological disorders [27,28]. 14-3-3 proteins represent a homologous family, which bind their substrates through an amphipathic binding cleft leading to a variety of effects, including stress response, apoptosis, transcriptional regulation, and cell cycle regulation [24]. Together, targeting these cell fate pathways represent a potential neuroprotective strategy in an immature brain.
In this report, we show the effect of WJ-MSCs in a clinically relevant murine model of immature brain injury. WJ-MSCs transplantation exerts protective effects at both histological and behavioral levels. We provide evidence that WJ-MSCs target pivotal regulators of the cell fate as well. Our result supports the notion of WJ-MSCs as an attractive cell graft in immature infants at risk.
Materials and Methods
All procedures have been approved by the local institutional review board (Ethics Committee and Veterinary Department of the Canton of Berne, Switzerland). We obtained a written informed consent from all umbilical cord donors.
Cell graft: WJ-derived MSCs
Human WJ-MSCs were isolated and characterized as previously published [12,29]. Briefly, umbilical cords were chopped, digested in 270 U/mL collagenase II (Worthington Biochemical Corporation, Lakewood, NJ) at 37°C/5% CO2, and cultured in Dulbecco's modified Eagle's medium (DMEM)/F12 10% containing fetal calf serum (FCS), 2 mM GlutaMAX, and 100 U/mL penicillin/100 mg/mL streptomycin/250 ng/mL amphotericin B (Thermo Fisher Scientific, Waltham, MA). The WJ-MSCs were characterized by fluorescence-activated cell sorting as follows: Antibody stainings were performed for 30 min at 4°C. Antibodies were diluted in 1× phosphate-buffered saline (PBS), 1% FCS. FITC-conjugated mouse monoclonal antibodies against the following markers were used: human CD105 (AbD Serotec, Oxford, United Kingdom), human CD90 (Acris Antibodies, San Diego, CA), human CD45 (BD Pharmingen, Franklin Lakes, NJ), human CD34 (BD Pharmingen), human CD14 (Millipore, Billerica, MA), and human HLA-DR (BD Pharmingen). The unconjugated mouse monoclonal antibodies against human CD73 (BD Pharmingen) and CD19 (Millipore) were detected with Alexa Fluor 594-conjugated anti-mouse IgG antibody (Thermo Fisher Scientific). The positive fraction was determined using unstained cells control. After staining, cells were fixed in 1% paraformaldehyde (PFA) and 10,000 events were acquired on an LSR II flow cytometer (BD Biosciences, Franklin Lakes, NJ). Analysis was done using the FlowJo software (Tree Star, Inc., Ashland, OR).
Animal model: perinatal brain injury
We induced immature brain injury as we had previously described [21,30]. Briefly, neonatal Wistar rats were randomly divided into three groups (Sham, Injury, and Injury+WJ-MSCs; n = 19). The Injury and Injury+WJ-MSCs groups received lipopolysaccharides (LPS: 0.1 mg/kg BW, i.p.) at postnatal day 3 (P3). On the following day, the left common carotid artery was doubly ligated using a stereomicroscope and animals were subjected to mild hypoxia (8% O2/92% N2; 40 min). The Sham Group (n = 6) consisted of uninjured animals (0.9% NaCl instead of LPS i.p., exposure of carotid artery without ligation, no hypoxia and no further treatment). The Injury+WJ-MSCs Group (n = 7) received a single cell graft transplantation into the lateral ventricle. We transplanted 250,000 of WJ-MSCs into the left ventricle at postnatal day 11 as published previously [29]. Briefly, anesthetized animals were fixed on a block heated to 37°C in a small animal stereotaxic frame (David Kopf Instruments, Tujunga, CA). We injected a volume of 5 μL over ∼6 min into the lateral ventricle using a Laboratory Animal Studies Injector (Hamilton, Bonaduz, Switzerland) with a 32G needle. The needle was left in place for 2 min after injection and then withdrawn slowly. The Injury Group (n = 6) received no WJ-MSCs treatment, but the needle was placed into the ventricle without vehicle injection.
Functional tests
To assess the functional outcome after brain injury, we performed a walking pattern analysis [31]. Briefly, rats with stained hind paws were placed on an inclining gangway and walked up the gangway and into a dark box. Paper strips with footprints were scanned and stride length (the distance between steps of the same side) and toe distance (distance between the first and fifth toe) were measured for the left and right hind paws.
Perioperative care of the animals
See Supplementary Data (Supplementary Data are available online at
Tissue harvesting and immunohistochemistry
We performed brain harvesting on P39 (n = 19; Fig. 1C). Briefly, we deeply anesthetized animals with sodium thiopental (100 mg/kg body weight i.p.; Inresa, Freiburg, Germany) and sacrificed by cardiac perfusion with PBS followed by formaldehyde (4%; Merck, Darmstadt, Germany). Animals were decapitated and we removed brains surgically and fixed in formaldehyde solution (4%) for 2–4 h at room temperature (RT) followed by 4°C for a total time of 24–48 h. Fixed brains were embedded in paraffin and sectioned into 7 μm slices.
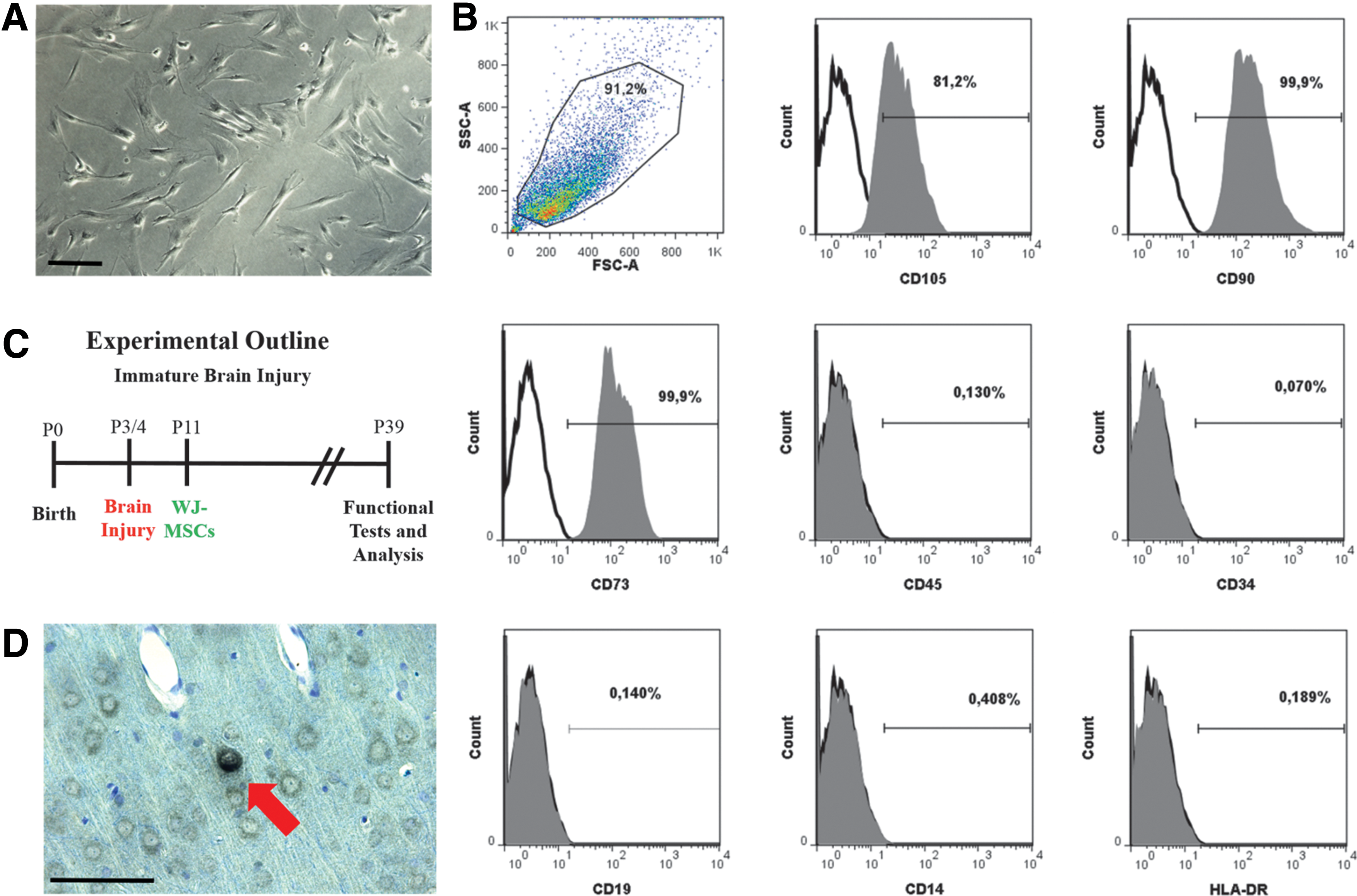
WJ-MSCs as a cell transplant.
For further details on tissue immunohistochemistry see Supplementary Data.
Tissue harvesting and western blots
To dissect WJ-MSCs protective mechanisms in the brain tissue (n = 3 each group), we induced brain injury as described above with following changes (see Supplementary Data for details). Whole brain lysates for kinome profiling were isolated following a protocol developed by Cell Signaling Technology (Danvers, MA). Briefly, tissue or cells were homogenized (lysate was passed through 20G needles 10 times for each sample) in a lysis buffer (20 mM HEPES pH 8.0, 9 M urea, 1 mM sodium orthovanadate, 2.5 mM sodium pyrophosphate, and 1 mM β-glycerophosphate), sonicated, and cleared by centrifugation. Protein concentration was measured using the Bradford assay and a total of 30 μg protein was loaded for each lane. Protein bands on western blots were quantified using ImageJ using β-actin as control [32]. The following antibodies were used: phospho-PKA/PKC substrate, No. 9624, 6967; phospho-CDK/tXR substrate, No. 9477, 8139; phospho-ATM/ATR substrate, No. 9607, 6966; phospho-Akt substrate, No. 9614, 10001; 14-3-3 binding partners, No. 9442; all from Cell Signaling Technology. For leukemia inhibitory factor (LIF) detection, we used an antibody from Millipore (No. MAB4306; CA).
Induction of WJ-MSCs into neural progenitor cells and gene array
To further characterize WJ-MSCs, we differentiated them at cell culture passage 5 (WJ-MSC derived from preterm birth: gestational week <37 weeks) into neurospheres [12]. Briefly, after an immediate differentiation step in DMEM/F12 medium containing 1 × N2 supplement (Life Technologies), 10 ng/mL epidermal growth factor (EGF; R&D Systems, Minneapolis, MN), 2 mM GlutaMAX, and 100 U/mL penicillin/100 mg/mL streptomycin/250 ng/mL amphotericin B, the cells were cultured in suspension in neurobasal medium (Life Technologies) containing 1 × B27 supplement (Life Technologies), 20 ng/mL EGF, 20 ng/mL basic fetal growth factor (bFGF; PeproTech, Rocky Hill, NJ), 2 mM GlutaMAX, and 100 U/mL penicillin/100 mg/mL streptomycin/250 ng/mL amphotericin B to induce neurosphere-like bodies. We extracted RNA and proteins from cell culture (WJ-MSC at passage 5 and neurospheres) using the QIAshredder and the RNeasy Plus Mini Kit from Qiagen (Hilden, Germany: according to the protocol), and proteins as previously performed [12]. RNA concentration was measured by NanoDrop spectrometry (Thermo Scientific, Wilmington, DE). Up to 5 μg of RNA were reverse transcribed using the RT2 First-Strand cDNA (Qiagen). The neurotrophins and receptors RT2 profiler PCR array (Qiagen) was used to measure the transcription of genes related to neurotrophic signaling. The plates were supplied precoated with forward and reverse primers. For one 96-well plate, 102 μL cDNA was mixed with 1,350 μL 2 × RT2 SYBR Green ROX qPCR mastermix and 1,248 μL nuclease-free water. Twenty-five microliters of the PCR component cocktail were pipetted into each well. The following cycling program was used to run the PCR on a 7300 Real-Time PCR System (Life Technologies): 10 min at 95°C, followed by 40 cycles of 15 s at 95°C and 1 min at 60°C.
Quantitative assessment of the brain
We performed quantitative assessments in the region of interest (ROI) defined as deep cerebral cortex (cortical layers V and VI in Figure 108 according to a neonatal rat brain atlas) [33] and in each hemisphere independently. Notably, ROI is the expected area of injury and hypoxia ischemia was reported to cause distinctive neuropathological alterations in these regions of the parieto-occipital cortex and white matter tracts [21,30,34]. We acquired images using a BX51 microscope (Olympus, Tokyo, Japan) with a 40× objective and equipped with a digital camera. An independent observer acquired six consecutive ROI sections visual field by visual field without overlapping per hemisphere and animal for each specific immunostaining blinded to the experimental conditions. To asses myelin loss (myelin basic protein [MBP] immunostaining) and astrogliosis (glial fibrillary acidic protein [GFAP] immunostaining), we measured the percentage of positive MBP and GFAP staining (area) in the ROI in each hemisphere as described previously [35,36]. We determined the difference between contralateral (undamaged) and ipsilateral (damaged) hemispheres using ImageJ [32].
Data analysis
We avoided variability resulting from tissue handling and staining, and inter-animal developmental variations by using a ratio of left (ipsilateral to carotid occlusion) to right hemisphere [37,38]. Data are represented as mean ± SEM. Single comparisons to control were made using two-tailed Student's t-test or Mann–Whitney test. We used one-way repeated measures ANOVA followed by Bonferroni's Multiple Comparison Test for multigroup design. P < 0.05 was considered to be statistically significant. No statistically significant differences were detected between the contralateral (uninjured hemispheres) in all groups. For gene array, data analysis was performed by the delta-delta Ct method using the company's software provided on their webpage and expressed as fold change in WJ-MSC derived from preterm delivery (n = 3) relative to neurospheres from preterm birth (n = 3). Data handling and statistical processing was performed using Microsoft Excel and GraphPad Prism Software.
Results
WJ-MSCs as a cell graft
We decided to use WJ-MSCs as a cell transplant as they are considered an attractive source for perinatal transplantation [11,39]. The isolated WJ-MSCs were adherent to plastic and displayed a fibroblast-like phenotype (Fig. 1A) [12]. Furthermore, after five passages, we detected characteristic MSCs cell surface markers (Fig. 1B) [40]. To test our hypothesis that our cell graft protects the immature brain, we used a well-established murine model (Fig. 1C) [21,30]. We evaluated the migration of WJ-MSCs by staining for HLA-ABC-positive cells at P39. In line with previous studies, we detected only few positive cells (Fig. 1D) [41] in the cortex cerebri, hippocampus, and striatum [29]. We did not detect any signs of differentiation in line with previous studies [42,43].
WJ-MSCs protect against immature brain injury
To test the effectiveness of WJ-MSCs treatment in immature brain injury, we evaluated the locomotor behavior first (Fig. 2A). In line with previous reports, we detected abnormal walking behavior after the brain insult, namely a significant change of step length and toe distance (Fig. 2A; compare left to right hind paws' step length and toe distance of Injury vs. Sham groups) [31]. WJ-MSCs transplantation resulted in the alleviation of spastic paresis and restored motor symmetry (Fig. 2A; compare left to right hind paws' step length and toe distance of Injury vs. Injury+WJ-MSCs groups). To determine the underlying pathophysiology, we focused on the motor cortex with emphasis on the characteristic deep cortical layers as these regions are altered as a consequence of immature brain injury [34,44] and associated with impaired locomotor behavior [31].

WJ-MSCs protect the immature brain.
We evaluated the number of NeuN-positive cells (mature neurons) in deep cortical layers first as injury, and hypoxia ischemia at this time point were reported to cause distinctive neuropathological alterations in these regions [21,30,34]. We did not detect neuronal loss (Fig. 2B; compare ipsilateral injured to contralateral intact hemisphere Sham vs. Injury) [35,45]. However, we need to consider that in contrast to our previous studies [21,30], we induced a mild injury (hypoxia reduced to 40 min). Additionally, the most common neuropathology in premature infants is not focal necrosis, but diffuse noncystic alterations [14,18]. As these alterations evolve over time to glial scars characterized by microgliosis and astrogliosis, we tested them next.
Given that the hallmark of immature brain injury is the degeneration of immature oligodendrocytes and a resulting loss of mature oligodendrocyte markers like MBP, we tested for myelin loss [46 –48]. We detected a significant myelin loss after injury (Fig. 2C: compare ipsilateral injured to contralateral intact hemisphere Injury vs. Sham groups) [38,49]. Importantly, transplantation of WJ-MSCs resulted in an amelioration of the induced MBP loss (Fig. 2C: compare ipsilateral to contralateral hemisphere Injury vs. Injury+WJ-MSCs groups). Besides demyelination, glial activation is another prominent event after immature brain injury, so we tested astroglial activation next [34,50]. We detected increased astroglial activation in the cortex following injury (Fig. 2D: compare ipsilateral to contralateral hemisphere Injury vs. Sham groups). In line with myelin protection, WJ-MSCs reduced astroglial activation significantly (Fig. 2D: compare ipsilateral to contralateral hemisphere Injury vs. Injury+WJ-MSCs groups). Collectively, our results support the notion that WJ-MSCs attenuate the induced injury in an immature brain characterized by cortical changes such as myelin loss and astroglial activation, which underlie the resulting locomotor behavioral changes [35,45].
WJ-MSCs modulate PKA/PKC, CDK, Akt signaling, and 14-3-3 binding in the immature brain
Our results indicate that transplantation of WJ-MSCs protects the immature brain against injury (Fig. 2 compare Injury vs. Injury+WJ-MSCs groups). However, the underlying mechanisms remain unclear. To investigate these mechanisms, we chose a global screening approach to detect key regulators of responses to DNA damage and probed the brain tissues against well-characterized motif-specific antibodies [21]. These antibodies detect specific posttranslational modifications in the context of specific binding motif or substrate. Thus, we assess not a single target, but rather have a global overview.
We tested PKA/PKC first as they are important signaling molecules in a variety of cellular functions, including cellular response to hypoxia ischemia or oxidative stress [21,51,52]. In line with previous reports, injury to an immature brain resulted in the activation of PKA/PKC signaling (Fig. 3A) [21]. Surprisingly, WJ-MSCs did not further activate PKA/PKC, but rather decreased it significantly (Fig. 3A).
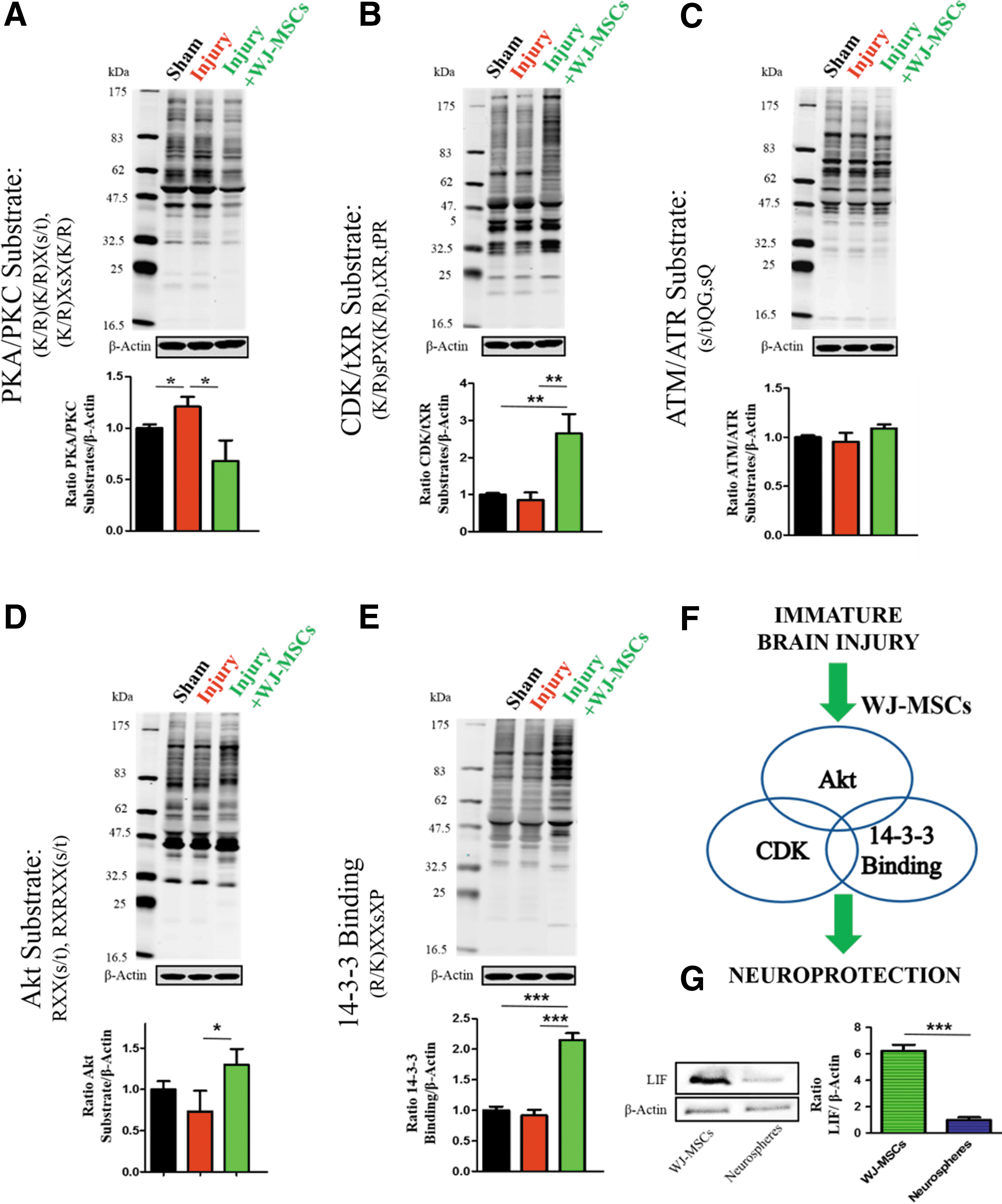
WJ-MSCs modulate crucial cell fate pathways. Proteins were extracted from frozen brain tissues (n = 3 each group) and analyzed by western blot. In upper panels representative western blots and in lower panels quantification of cyclic AMP-dependent protein kinase/calcium-dependent protein kinase (PKA/PKC)
In search for other potential mechanisms and given the emerging importance of CDK inhibitors in oncology and their potential use in stroke, we tested for CDK substrates [19]. In stroke, ischemic brain injury results in an activation of CDK signaling and CDK inhibition is protective [19]. Immature brain injury did not affect CDK signaling and we detected CDK activation after cell graft transplantation (Fig. 3B). To further elucidate the graft response, we tested ATM/ATR signaling as ATM is upstream of CDK in cell cycle arrest [26,53,54]. We did not detect any significant changes (Fig. 3C), which suggests that the observed cell graft-mediated CDK activation is ATM/ATR independent. Next, we tested phosphorylated Akt substrates and 14-3-3 binding partners as they modulate the cell cycle progression, are crucial CDK partners, and reported neuroprotective targets [20,24,55,56]. In line with CDK activation, WJ-MSCs treatment increased phosphorylation of Akt substrates and 14-3-3 binding partners (Fig. 3D, E). Together, our results provide evidence that WJ-MSCs protect the immature brain and modulate cell fate signaling pathways (Fig. 3F) involved in brain protection.
WJ-MSCs express neurotrophic factors
Given the modifications after transplantation of WJ-MSCs (Fig. 3) and WJ-MSCs' protective effects (Fig. 2), we aimed to further characterize the cell graft using a gene array [12]. We decided to compare WJ-MSCs to differentiated neurospheres derived from WJ-MSCs as WJ-MSCs have a multi-differentiated potential. It is the potential to express factors from other tissues before any differentiation. Thus, we aimed to detect changes unique to the relatively heterogeneous cell population of WJ-MSCs [12]. In line with this potential, we detected increased expression of neurotrophic factors and receptors involved in brain development and response to injury such as nerve growth factor receptor (NGFR) and GDNF family receptor alpha 3 (GFRA3) [57,58] (all genes summarized in Table 1). We detected increased expression of prostaglandin E receptor 2 (PTGER2), which in stroke protects against oxidative stress in a PKA-dependent manner [59]. Interestingly, we detected increased expression of LIF as well. LIF is an important factor contributing to a variety of cellular processes such as neuronal development, astroglial responses to injury, and oligodendrocyte survival [60 –63]. Importantly, LIF exerts its biological effects partially through Akt, CDK, and 14-3-3 signaling pathways [61,64,65], which makes it a potential candidate of the WJ-MSCs' induced modifications (Fig. 3). We confirmed increased LIF expression in WJ-MSCs on protein level as well (Fig. 3G).
We examined gene expression using specific profiler polymerase chain reaction array. The table shows significantly expressed genes of WJ-MSCs compared with neurospheres derived from WJ-MSCs in culture. We used an arbitrary cutoff of more than twofold change and P < 0.05.
WJ-MSC, Wharton's Jelly-derived mesenchymal stem/stromal cell.
Together, we hypothesize that WJ-MSCs may exert adaptive responses depending on the type of injury they are facing, making them prominent candidates for cell therapy in perinatal injuries.
Discussion
This report documents that WJ-MSCs transplantation exerts protective effects against immature brain injury. Using a clinically relevant model, we demonstrate that our cell graft (Fig. 1) is able to ameliorate myelination loss and astroglial activation while preserving proper functional behavior (Fig. 2). Most importantly, we are the first to report that WJ-MSCs target crucial signaling pathways involved in cell cycle regulation and apoptosis (Fig. 3F), while expressing growth and differentiation factors (Table 1). Together, we provide evidence that WJ-MSCs are potential candidates for treatment of immature infants at risk.
MSCs and MSC-derived therapies emerged in the last decades as a potential paradigm shift for complex multifactorial diseases, such as immature brain injuries [66]. As the preterm infant faces multiple risk factors, including inflammation/infection, ischemia/reperfusion, and deprivation of placental protective factors, an optimal therapeutic should target the injuries globally (several organs) and locally (response to signals of local injury) [11]. In our study, WJ-MSCs protect the immature brain and modulate different signaling pathways. This is particularly interesting as it fuels the constant debate whether MSCs or MSC-derived products such as conditioned media or exosomes are the optimal choice for treating perinatal injuries. We hypothesize that the WJ-MSCs' ability to locally respond to signals after injury is an advantage, but the direct comparison between WJ-MSCs and their products in perinatal injury models needs to be assessed in future studies [66]. From a clinical point of view, precautions and long-term adverse consequences need to be considered as in the immunocompromised developing preterm infant, stem cells have the theoretical potential to enhance tumor growth [67].
Another important consideration is the route of stem cell transplantation and the animal model. We tested WJ-MSCs in a well-established model of immature brain injury [21,30,35]. We induced injury at P3/4, a time point that corresponds to a high-risk period of human preterm infants with immature oligodendrocyte development peak [45]. Expectantly, we detected myelin loss with motor and sensory deficits (Fig. 2A, C) [35,45]. However, preterm survivors demonstrate cognitive and behavioral deficits with higher incidence of psychiatric disorders as well [5,6]. Thus, studies identifying MSC's effects after P3/4 injury are needed, especially, as injury at P4 may result in postnatal maturation and affective disorders [68]. Multiple transplantation routes for brain injuries, including intracerebral, intraperitoneal, intranasal, or intravenous, were introduced [67]. Indeed, we chose an invasive route, but a straightforward approach, as we aimed to determine direct effects on the immature brain. Given that the graft preparation and optimal administration route remains unclear [69], further noninvasive studies are warranted, but beyond the scope of this article [29,70]. From a clinical point of view, evaluation of the optimal cell transplant is obligatory. Although we did not detect morphological changes of the cell graft over time (Fig. 1D), longitudinal assessment of the cell graft fate are still needed, especially as we used a xenograft. Additionally, preparation of the graft before transplantation such as removal of fetal bovine serum needs to be assessed before clinical utilization.
Numerous studies postulate that MSCs protective effects after perinatal injury are based on paracrine effects and immunomodulation [11]. In contrast to upcoming novel immunomodulation therapies such as preimplantation factor, WJ-MSCs did not impact PKA/PKC signaling after injury [21]. Given the diverse modes of action, a combined therapeutic approach may provide superior protection due to synergistic effects. Another surprising finding is that WJ-MSCs activate CDK signaling in the brain, whereas CDK inhibition protects neurons in stroke or traumatic brain injury [19,71]. However, when considering this opposite effect, the distinct pathophysiology of an immature brain and time point of the evaluation need to be accounted for. For example, in an immature brain, cell proliferation and differentiation, especially in the case of oligodendrocyte progenitors, depend on proper CDK activity [53,72]. Furthermore, in stroke CDK activation was detected up to 24 h of reperfusion injury only [19]. Given that CDK modulation was not detected after injury (Fig. 3B: compare Injury vs. Sham) and neuroprotective inhibition of CDKs is present in adult animals only [73], suggest an age-dependent role of CDKs. Together, we provide evidence that WJ-MSCs modulate crucial cell fate pathways in an immature brain injury setting. We hypothesize that targeting these pathways impacts the regeneration capacity of a developing brain. Additional studies will dissect which specific pathways such as AKT/CDK/14-3-3 binding and factors such as LIF may produce beneficial effects. Furthermore, studies addressing the direct comparison of different cell transplants, including adipose-derived stem cells and bone marrow mononuclear cells, will determine whether the documented effects (Fig. 3) are WJ-MSC specific as well.
In conclusion, disease-specific and age-specific considerations are necessary when assessing therapeutics for immature infants. Additional studies identifying WJ-MSCs' optimal formulation and preparation (live cells vs. cell products), transplantation routes (invasive vs. non-invasive), and design (alone vs. synergistic additives) need to be addressed before clinical translation. We hypothesize that WJ-MSCs may exert adaptive responses depending on the type of injury they are facing, making them prominent candidates for cell therapy in perinatal injuries.
Footnotes
Acknowledgments
This study was supported by Cryo-Save AG, Switzerland; Mobiliar Jubiläumsstiftung, Switzerland; and Forschungs-Grant Inselspital Bern, Switzerland.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.