
Abstract
Mesenchymal stem cells (MSCs) were initially identified as progenitors of skeletal tissues within mammalian bone marrow and cells with similar properties were also obtained from other tissues such as adipose and dental pulp. Although MSCs have been extensively investigated, their native behavior and in vivo identity remain poorly defined. Uncovering the in vivo identity of MSCs has been challenging due to the lack of exclusive cell markers, cellular alterations caused by culture methods, and extensive focus on in vitro properties for characterization. Although MSC site of origin influences their functional properties, these mesenchymal progenitors can be found in the perivascular space in virtually all organs from where they were obtained. However, the precise identity of MSCs within the vascular wall is highly controversial. The recurrent concept that MSCs correspond to pericytes in vivo has been supported mainly by their perivascular localization and expression of some molecular markers. However, this view has been a subject of controversy, in part, due to the application of loose criteria to define pericytes and due to the lack of a marker able to unequivocally identify these cells. Furthermore, recent evidences indicate that subpopulations of MSCs can be found at extravascular sites such as the endosteum. In this opinion review, we bring together the advances and pitfalls on the search for the in vivo identity of MSCs and highlight the recent evidences that suggest that perivascular MSCs are adventitial cells, acting as precursors of pericytes and other stromal cells during tissue homeostasis.
Introduction
O
However, until recently and despite the huge research effort directed to MSCs, little progress was made in understanding their in vivo identity and function. The available data are still controversial and their exact role in vivo is a subject of much debate.
Moreover, as critically addressed previously [1], the rationale for the use of MSCs in some clinical trials seems to be inconsistent with their ascribed native biology. This may explain why although the number of clinical trials using MSCs has expanded over the years, the distribution of trials by phase remains practically unaltered [2], suggesting that many trials are not progressing through the clinical pipeline. These drawbacks further highlight the need for a research effort to comprehend the native identity and function of MSCs to refine and thus improve their clinical application.
In this opinion review, we revisit the controversial issues involved on the search for the in vivo identity of MSCs and present a model that accommodates the recent evidences regarding their putative perivascular identity.
The Evolution of MSC Concept
In 1968, Tavassoli and Crosby were the first to report that autologous fragments of rodent bone marrow (BM) were able to generate de novo heterotopic marrow and bone upon transplantation [3]. Two years later, Friedenstein et al. identified a subpopulation of stromal cells responsible for the BM osteogenic potential. Following culture of BM cells, they observed that some rare stromal cells had a high replicative potential and therefore were able to form colonies of fibroblastic cells in vitro, rendering their denomination of colony-forming unit–fibroblast (CFU-F) [4]. When the progeny of a single CFU-F was implanted either subcutaneously or beneath the kidney capsule, it was able to form ectopic BM comprising trabecular bone, chondrocytes, adipocytes, and hematopoiesis-supporting stroma [5]. Due to their differentiation capacity, these cells were named as osteogenic stem cells [6] or BM stromal stem cells [7]. Few years later, Arnold I. Caplan coined the term mesenchymal stem cell (MSC) [8] to designate the CFU-F identified by Friedenstein et al. based on the premise that such cells would behave as unique progenitors for all connective tissues of mesodermal origin.
Since then, the designation, mesenchymal stem cells, has been widely adopted by the scientific community, especially after the demonstration of their multilineage potential in vitro [9]. Subsequently, many groups reported the isolation of MSC-like cells from several tissues and organs in addition to BM, such as umbilical vein [10], saphena vein [11], bone [12], skeletal muscle, lung, deciduous teeth [13], adipose tissue [14], and placenta [15].
In addition to the isolation of MSCs by adherence to plastic, several cell surface markers have been used to prospectively isolate cell populations enriched with MSCs [16 –18].
For the vast majority of these studies, MSCs had been identified based on in vitro properties such as cell morphology, immunophenotypic profile, and in vitro multilineage potential. Due to the diversity of methods employed by distinct laboratories to culture and identify MSCs, the International Society for Cell Therapy published a set of criteria as an attempt to standardize the characterization of MSCs [19]. Unfortunately, the extensive use of in vitro properties to characterize MSCs has hindered the elucidation of their native identity. One obvious reason for this is that so far there is no specific marker that distinguishes cultured MSCs from other stromal cell types. Besides, the expression of some cell surface molecules is affected by culture conditions. For example, cultured human BM-derived MSCs (BM-MSCs) upregulate the expression of CD146 in normoxia and downregulate in hypoxia [20]. Furthermore, some authors postulate that not only the immunophenotypic profile but also the differentiation potential used to define MSCs in vitro is shared with other connective tissue cells [21]. Accordingly, we have shown that human BM-MSCs, human retinal CD146+ pericytes, and human fibroblast cell lines share cell morphology, the expression of 22 cell surface molecules, and also the capacity to differentiate toward osteogenic, adipogenic, and chondrogenic lineages in vitro [22], leading to the anticipated conclusion that they are identical cells. However, by using serial analysis of gene expression, we showed that fibroblasts have a global transcriptional profile distinct from that of MSCs and pericytes and that these cell types are not coincident.
Only years after the discovery of BM-MSCs, characterization studies using stringent in vivo assays paved the way to elucidate their native identity and to confirm their stemness. Indeed, in the vast majority of studies, MSCs are referred as the population of fibroblastic cells obtained after culturing unfractionated cells from dissociated organs and tissues. Therefore, despite its global usage, the term, mesenchymal stem cells, has been criticized since it could lead to misconceptions regarding the biology of this unfractionated cultured cell population that do not meet the generally accepted criteria for stem cell activity [23,24]. Stem cells are characterized by functional assays and, accordingly, a bona fide MSC should be able to self-renew and to differentiate in vivo as the hematopoietic reconstitution in serial transplantation is used to define functional hematopoietic stem cells. The ability of BM-MSCs to self-renew in vivo was only demonstrated recently in serial transplantation assays in mice. These experiments showed that the progeny of a single murine CFU-F is able to generate bone-containing cartilage, adipocytes, fibroblasts, and hematopoiesis-supporting stroma in primary and secondary recipients [25,26]. These cells, therefore, fulfill the criteria of stem cells of generating fully differentiated descendants in vivo (multipotency) and of reconstituting in vivo cells identical in phenotype and potency to those explanted (self-renewal).
Such in vivo studies and stringent in vitro characterization have also contributed to dismantle the concept of MSCs as a population of progenitor cells endowed with identical biological properties regardless of their native microenvironment. For instance, human adipose-derived MSCs display reduced osteogenic and chondrogenic potentials, secrete higher amounts of angiogenic molecules such as vascular endothelial growth factor (VEGF)-A and VEGF-D, and display a distinct transcriptional profile in comparison with their BM-derived counterparts [27 –30]. Thus, a more realistic view is the existence of not one but several MSC populations whose biological properties are affected by their tissue of origin. Interestingly, however, in situ analyses have been demonstrating that human and mouse MSCs can be obtained after culturing perivascular cells, regardless of their anatomical source.
As discussed previously, the phenotype of MSCs is altered during culture and it is unclear whether a cell population identical to cultured MSCs exists in vivo. Thus, in this review, we refer to in vivo MSCs as genuine stromal stem cells capable of self-renewing and of originating a differentiated progeny during cellular turnover and regeneration. When cultured, these in vivo MSCs would give rise to their cultured counterpart as we know.
MSCs Reside at Perivascular Sites
The first experimentally based proposition for the in vivo niche of MSCs came out after the work of Shi and Gronthos in 2003 [31]. Gronthos’ laboratory first demonstrated that selection of stromal cells based on the high expression of STRO-1 antigen enriches all detectable CFU-Fs in human BM samples and select cells with in vivo osteogenic capacity [32]. Later, they showed that expression of STRO-1 was confined to the vascular wall in both human BM and dental pulp histological sections. These findings led to the proposition that MSCs reside at perivascular sites.
This hypothesis was further strengthened by other studies reporting the isolation of cells with MSC characteristics from the vascular wall of human saphena vein [11], aorta [33], and umbilical cord vein [10]. This emerging view of MSCs as perivascular progenitor cells seemed attractive since it would conveniently explain why these cells could be obtained from virtually all vascularized organs and tissues [34]. Interestingly, perivascular cells are also central for the stem cell activity in other organs such as lymphoid follicles and skeletal muscle. Perivascular cells expressing platelet-derived growth factor receptor β (PDGFRβ) were identified as precursors of follicular dendritic cells in mice [35,36], and perivascular cells positive for neuroglial 2 proteoglycan (NG2) interact with satellite cells in human and mouse skeletal muscle regulating their differentiation and quiescence through paracrine signaling [37].
Alternative experimental approaches further supported a perivascular niche for MSCs. In a comparative study of gene expression profiles and differentiation capacities conducted by our group, human MSCs derived from BM and umbilical cord were found to be more similar to human retinal perivascular CD146+ stromal cells than to other stromal cell types such as fibroblasts, smooth muscle cells, and hepatic stellate cells [22]. In the same year, Crisan et al. had reported a seminal work demonstrating that perivascular CD146+ human stromal cells obtained from different fetal and adult organs display MSC characteristics in vitro [38].
Based on the findings of this last report, some authors postulated that MSCs would correspond to pericytes in vivo [39]. However, although it is widely accepted that MSCs occupy perivascular locations in different organs, their precise identity within the vascular wall remains controversial.
Are MSCs Pericytes?
Pericytes were described 140 years ago as a population of contractile cells adjacent to the endothelium of small blood vessels [40]. This definition was refined after the observation that pericytes are partially or completely embedded within the vascular basement membrane [41]. Even with significant advances on the search for pericyte molecular markers, their anatomical characteristics have been frequently used to define this cell population [42].
As a corollary of this definition, the hypothesis that MSCs are pericytes in vivo implicates that these progenitor cells could only be obtained from preparations containing small vessels such as capillaries, venules, and/or arterioles. However, we and others have reported the isolation of MSCs from large human vessels such as aorta [33], saphena vein [11], and umbilical cord vein [10]. These observations compellingly demonstrate that MSC cultures cannot be exclusively derived from pericyte-containing microvessels.
At this point, however, it is important to acknowledge that the concept of pericytes as mural cells exclusively encircling microvessels has been challenged by the observation of pericyte-like cells forming a subendothelial network around large vessels [43]. This concept emerged from the studies of Langhans in 1866, who described the presence of subendothelial cells surrounding large vessels, which were distinct from smooth muscle cells [44]. In 1977, Shchelkunov also described this cellular layer under the endothelium of large arteries and veins in different chordate species [45]. He hypothesized that these subendothelial cells form a continuum layer from aorta to capillaries, where they were identified as pericytes. By using morphological studies associated with immunostaining of 3G5 antigen, Andreeva et al. further strengthened this hypothesis by demonstrating the presence of a continuum network of pericyte-like cells intimately associated with the endothelium in vessels of different types and caliber [46].
The putative ubiquitous distribution of pericytes throughout the vasculature implicates that the confirmation of a pericytic phenotype for MSCs would require a careful fractionation of perivascular cell populations, followed by the investigation of MSC traits in these cells.
In line with this experimental strategy, the majority of evidences supporting the pericytic phenotype of MSCs rely on the prospective isolation of cells using molecular markers expressed by pericytes, followed by the investigation of MSC traits in the isolated cells. For instance, the first suggestion that human MSCs would be pericytes in vivo was based on selection of cells whose immunophenotypic profile is similar to that of pericytes (CD146+CD34−CD45−CD56−), followed by the demonstration that these cells could give rise to osteogenic, chondrogenic, and adipogenic lineages in vitro [38].
Numerous studies have demonstrated that freshly isolated cells expressing pericyte markers are enriched with MSCs. In mouse BM, nestin-positive cells contain all CFU-F activity and are able to spontaneously differentiate into adipocytes and osteoblasts in vitro. Furthermore, these cells contribute to the formation of a hematopoietic niche after serial transplantation in vivo, as expected for genuine BM-MSCs [26]. In humans, expression of CD146, a cell surface glycoprotein present in pericytes, also defines a subpopulation enriched with MSCs in several adult and fetal tissues [38]. Furthermore, adipose tissue-derived cells expressing 3G5 were shown to be perivascular and contained MSCs capable to differentiate into ectopic bone following in vivo transplantation [47].
All these prospective isolation approaches contributed to support a perivascular niche for MSCs. However, a more precise determination of the in vivo phenotype of MSCs based solely on expression of cell surface molecules is limited. This is because none of the abovementioned markers (nestin, CD146, 3G5), for instance, is expressed exclusively by pericytes. As an example, 3G5 antigen is also expressed by cells present in the media and adventitia layers of large veins and arteries in human samples [46].
So far, a molecular marker able to unequivocally identify pericytes is still unknown. This not only limits the interpretation of flow cytometry data but also precludes the development of robust in vivo lineage tracing experiments to demonstrate that cells with MSC traits can be obtained exclusively from pericytes. For example, using a Cre-loxP mouse model for lineage tracing, it was suggested that BM osteoprogenitors are derived from cells expressing alpha smooth muscle actin (αSMA) [48]. Despite being expressed by some pericytes [49], αSMA is also present in myofibroblasts and vascular smooth muscle cells. In another study, PDGFRβ was used as a marker to demonstrate the perivascular origin of mouse white adipocyte progenitors [50]. Subsequently, constitutive activation of endogenous PDGFRβ by targeted mutagenesis was shown to suppress adipogenic differentiation while maintaining progenitor potential in vivo, indicating that this pericyte marker identifies adipocyte progenitors and is essential for adipogenesis [51]. However, PDGFRβ is also expressed by some fibroblasts, vascular smooth muscle cells, and capillary endothelial cells [52].
The absence of a molecular marker profile shared exclusively by MSCs and pericytes precludes a definitive association between these two cell types. However, are there molecular markers commonly expressed by pericytes that are not found in MSCs? It was previously demonstrated that capillary pericytes are consistently positive for desmin in vivo and after culture [53,54], a characteristic that is also shared by vascular smooth muscle cells around large blood vessels [55]. However, cultured MSCs isolated from mouse, rat, and human tissues are all negative for desmin [56 –58], indicating that these cells are not the ex vivo counterpart or derived from mature pericytes. However, as a note of caution, it remains unclear whether the conditions used for culturing MSCs reduce desmin expression.
In face of these uncertainties, which perivascular cell type is likely to represent the native identity of MSCs? As we shall discuss bellow, dissection techniques and the use of molecular markers already allow the fractionation of perivascular cell populations. As a result of these approaches, several studies postulate that MSCs are adventitial cells occupying the outermost perivascular cell layer. Of note, both pericytes and adventitial cells share the expression of nearly all markers used for the identification of MSCs in lineage tracing and prospective isolation studies (Table 1).
It refers to the expression of markers in situ. Both pericytes and adventitial cells express most of the markers used to identify MSCs in situ.
MSCs, mesenchymal stem cells.
MSCs as Adventitial Cells
The tunica adventitia is the outermost cell layer that surrounds blood vessels. In large vessels such as arteries, it consists in a heterogeneous network of connective tissue comprising vasa vasorum, nerve endings, inflammatory cells, collagen fibers, and fibroblasts, among others [59]. However, as discussed in more detail below, adventitial fibroblastic cells were also identified around capillaries in adipose tissue, where they did not coexpress pericyte markers, but share the perivascular space with pericytes [60]. Therefore, these adventitial fibroblastic cells might be present in all extensions of the vascular system, not only in the outermost cell layer of large vessels.
The pioneering work of Hu et al. [61] suggested that adventitia contains cells expressing stem/progenitor markers (eg, Sca-1+, c-kit+, and CD34+) that differentiate into vascular smooth muscle cells in a mouse model of atherosclerosis. This evidence was followed by several studies from different groups demonstrating that the adventitia may be a perivascular reservoir of stem/progenitor cells with critical role in vascular remodeling and disease [62].
The hypothesis that adventitia contains MSCs was first explored by Hoshino et al. in 2008 [63]. In their work, the authors dissected the adventitia from pulmonary artery and established explant cell cultures. After incubation in specific inductive media, these adventitial cells were able to differentiate into adipocytes, osteoblasts, and smooth muscle cells and also expressed MSC markers after culture.
Studies in BM also indicate that MSCs are adventitial cells in vivo. Sacchetti et al. [25] demonstrated that perisinusoidal human CD146+ cells can differentiate in bone, self-renew, and also organize a hematopoietic niche in vivo after subcutaneous transplantation in mice. Later, Tormin et al. [20] demonstrated that BM CFU-F activity is exclusively present in the stromal CD271+ cell population, which can be either CD146+ or CD146-/low. A recent study confirmed that BM-MSCs are highly enriched in stromal CD271+ cell population [64]. All single-cell-derived colonies generated from CD271+ population were able to differentiate at least into osteocytes in vitro. When subcutaneously transplanted into immunodeficient mice, CD271+C45- cells differentiated in ectopic bone containing host hematopoietic cells. These cells were also found at the perivascular space following intrafemoral injection both in primary and in secondary recipient mice. Importantly, when the authors stained human BM samples with anti-CD271 antibody, they observed that perivascular CD271 expression was exclusively confined to the cells composing the tunica adventitia.
Immunohistochemical studies using human adipose tissue and internal thoracic artery specimens demonstrated that adventitial cells and pericytes can be discriminated based on the expression of CD34 and CD31 [65,66]. Staining large blood vessels with anti-CD34 antibody revealed two concentric rings comprising CD34+ cells. The internal ring displayed a contiguous staining for CD34, which overlapped with CD31 expression. These were endothelial cells. Interestingly, the outer ring consisted in the adventitia and comprised CD34+CD31− cells. A subsequent work carefully evaluated the distribution of these CD34+CD31− adventitial cells in blood vessels from human adipose tissue samples [60]. The authors demonstrated that CD34+CD31− adventitial cells did not express αSMA and were located at the vascular wall of arteries and veins (diameter of 75–150 μm) and of arterioles and venules (diameter of 25–75 μm). Interestingly, CD34+αSMA−CD31− adventitial cells were also observed around capillaries (diameter of 10 μm), sharing the perivascular space with PDGFRβ+ pericytes.
The elucidation of molecular markers differentially expressed by adventitial cells and inner perivascular cells (including pericytes) paved the way for prospective isolation, culture, and characterization of cell populations enriched with either of these two cell types. Taking advantage of this strategy, Corselli et al. provided experimental evidences that human adipose-derived MSCs are adventitial cells within the vascular wall [67]. The authors demonstrated that CD34+CD146−CD31− adventitial cells from adipose tissue expressed the MSC markers, CD44, CD73, CD90, and CD105, in situ. In addition, these cells were able to differentiate toward adipogenic, chondrogenic, and osteogenic lineages in vitro. When comparing the phenotype of adventitial cells with that of CD34−CD146+CD31− pericytes, the authors observed that cultured pericytes were consistently positive for other pericyte markers such as αSMA, PDGFRβ, and NG2, besides CD146 used for their isolation. In a sharp contrast, cultured adventitial cells did not express any of these proteins. However, adventitial cells treated with angiopoetin-2 for 72 h upregulated all the abovementioned pericyte markers. Based on these observations, it was suggested that MSCs are adventitial cells and precursors of pericytes. The concept that MSCs are not mature pericytes, but only share the perivascular space with them, was previously speculated by our group [22], and the experimental evidences provided by Corselli et al. [67] further strengthen this hypothesis.
Indeed, Howson et al. [68] previously demonstrated the existence of pericyte progenitors within the vascular wall of rat aorta specimens. The authors isolated a population of CD34+CD31−aSMA− perivascular cells that formed nonadherent spheroids upon cultivation in serum-free conditions. However, following incubation in fetal bovine serum, these cells downregulated the expression of CD34 and started to express pericyte markers such as αSMA, calponin, and desmin. These resulting pericytes were differentiated cells that lost the capacity to form new spheroids and were able to exhibit perivascular homing in neovessels sprouting from explanted fragments of rat aorta.
Another recent report provided robust evidences supporting the role of MSCs as more primitive progenitor cells capable of originating pericytes. Using Cre-recombinase models for in vivo lineage tracing, Zhao et al. [69] observed that Gli1+ cells in mouse incisor are genuine MSCs capable of originating all stromal cells during mesenchyme turnover and injury repair. These Gli1+ cells occupy the outermost layer around arterioles and, surprisingly, did not express any of the typical murine MSC markers in situ (ie, CD105, CD73, NG2, CD146, CD44, Sca-1, and nestin).
Unlike Gli1+ periarterial cells, NG2+ pericytes within tooth mesenchyme coexpressed the classical MSC markers, CD105, Sca-1, and CD146, in situ. However, these NG2+ pericytes did not give rise to other stromal cells during physiological mesenchyme turnover, but only originate some odontoblasts during injury repair [69].
Notably, permanent staining of Gli1+ periarterial cells and NG2+ pericytes using Cre-recombinase models demonstrated that Gli1+ cells give rise to NG2+ pericytes in vivo. Upon cultivation, Gli1+ cells lost the expression of this marker. Both cultured Gli1+ and NG2+ cells expressed the typical MSC markers, CD146, CD105, Sca-1, and CD73, and were capable of adipogenic, osteogenic, and chondrogenic differentiation in vitro. However, from all cultured MSCs obtained from unsorted tooth mesenchyme, 95% were derived from Gli1+ cells, while the remaining cells originated from NG2+ pericytes [69].
These results indicate that adventitial Gli1+ cells are genuine MSCs acting as mesenchyme precursors during homeostasis and tissue repair in mouse incisor. NG2+ pericytes practically do not contribute to mesenchyme turnover in mouse incisor and can give rise to other cell types only during injury repair.
MSCs at Extravascular Sites: Insights from Bone
Despite the recognition that MSCs occupy the perivascular niche, some studies have reported the identification of cells with MSC traits that do not exclusively populate blood vessel walls. So far, the most detailed evidences were reported from studies of BM.
The medullary cavity of long bones is mainly occupied by the BM, which contains hematopoietic cells, stromal cells, adipocytes, and numerous blood vessels. The periphery of the medullary cavity contains a thin layer of connective tissue known as endosteum, which lines the internal surface of the bony tissue. As referred previously, it was shown that all CFU-F activity of human BM-derived cells is exclusively present within the nonhematopoietic CD271+ population [20]. These CD271+ cells can be further classified on the basis of CD146 expression: CD271+CD146+ and CD271+CD146−/low cells. These two subpopulations not only contain all CFU-Fs in BM but also are capable of osteogenic differentiation in vivo and are found in contact with CD34+ hematopoietic stem cells, forming hematopoietic niches within the BM. However, a clear difference between these two subsets lies on their in situ localization: while CD271+CD146+ cells are perivascular, CD271+CD146−/low cells are found at the endosteum, that is, proximal to the surface of trabecular bone. These observations agree with previous findings demonstrating that CD271+ stromal cells can be either perivascular or bone lining cells [70] and that mouse BM contains both endosteal and perivascular hematopoietic stem cell niches [71].
Additional evidences of an endosteal population of MSCs were reported for mouse BM during in vivo lineage tracing of cells expressing Ebf2 [72], a transcription factor identified in osteoblast progenitors during fetal bone development [73]. In a Cre-recombinase mouse model in which the progeny of Ebf2+ cells expresses yellow fluorescent protein (YFP), the YFP+ cells coexpressing the osteoblast marker, osteocalcin, or the chondrocyte marker, aggrecan, were localized in proximity to bone surface and in articular cartilage, respectively. These findings are indicative of in vivo multilineage potential of Ebf2+ cells. In mice expressing GFP under the control of Ebf2 promoter, the authors observed that GFP/Ebf2+ cells primarily occupy the endosteum and are capable of differentiating into multiple mesodermal tissues in vitro and in ectopic bone in vivo. Furthermore, single-cell transcriptomic analysis revealed that GFP/Ebf2+ cells express several MSC-related genes, including nestin, which codifies for an intermediate filament whose expression was previously identified in mouse BM-MSCs [26].
Although most Ebf2+ cells were found at the endosteal niche, some of them were also observed around blood vessels, although at relatively low frequencies [72].
More recently, Chan et al. [74] employed a Rainbow mouse model [75] that allows the visualization of clonal regions to evaluate whether mesenchymal tissues in bone share a common progenitor. They observed that clonal regions were particularly concentrated at the growth plate and encompassed bone, cartilage, and stromal tissue, but not adipose tissue and muscle. In an independent study, Worthley et al. [76] compellingly demonstrated that the stem cells responsible for generation of bone, cartilage, and stroma during bone development, remodeling, and repair are concentrated close to the bony tissue within the metaphysis of long bones and not in the perisinusoidal space.
The occurrence of MSCs at extravascular sites seems to be a specific feature of the bone microenvironment. A model accommodating the recently found extravascular population of skeletal stem cells and the extensively characterized perisinusoidal skeletal stem cells has been proposed [77]. According to this theory, extravascular skeletal stem cells would be the major source of mesenchymal tissues during bone formation and development and the perisinusoidal skeletal stem cells would contribute to bone homeostasis later in the postnatal life. Of note, models comprising the existence of multiple populations of MSCs that do not coincide in space and time have not been sufficiently explored so far.
MSC as Result of Epithelial/Endothelial–Mesenchymal Transition
Epithelial–mesenchymal transition (EMT) is a biological process that enables polarized epithelial cells to undergo multiple biochemical changes, resulting in the acquisition of a mesenchymal phenotype [78]. Similarly, the endothelial–mesenchymal transition (EndMT) consists in the conversion of endothelial cells into mesenchymal cells [79]. Both processes are characterized by the disruption of intercellular adhesion, acquisition of a fibroblastic shape, and increased motility/invasiveness.
The conversion of epithelial/endothelial cells to mesenchymal cells is critical during ontogenesis [78]. For instance, during gastrulation in mammalians, epithelial cells at the primitive streak activate EMT and invade the subjacent matrix to originate the mesoderm and endoderm. Activation of EMT is also crucial for the dissemination of precursor cells from neural crest during ontogenesis [80]. In turn, induction of EndMT takes place during heart development when endothelial cells activate this morphogenetic program to originate the cardiac valves and to initiate heart septation [81].
Recapitulation of EMT and EndMT in postnatal life is often associated with pathological circumstances. It has been postulated that activation of EMT in carcinoma cells endows them with invasive traits, thereby facilitating metastasis [82], and induction of EndMT has been considered a source of stromal cells that contribute to cardiac fibrosis [83] and to the formation of tumor-supportive stroma [84].
It was previously demonstrated that induction of EMT endows breast epithelial cells with stem cell traits, as demonstrated by their capacity to regenerate mammary fat pads in mice [85]. However, the multilineage potential of EMT-derived cells was rarely explored. In 2010, two independent studies reported that the induction of EMT or EndMT might ultimately lead to the generation of stromal cells with MSC characteristics.
In one of these reports, Battula et al. [86] induced EMT in immortalized human breast epithelial cells by constitutive overexpression of the transcription factor, Twist or Snail, or by incubation in transforming growth factor (TGF)-β. The authors observed that unlike breast epithelial cells, EMT-derived cells displayed the antigenic profile commonly associated with MSCs (eg, CD73+, CD105+, CD44+, CD24-, and CD140b+) and were able to differentiate toward adipogenic, chondrogenic, and osteogenic lineages in vitro.
Although these EMT-derived cells generated by Battula et al. [86] displayed properties similar to that of BM-MSCs, Phinney published a letter highlighting one critical difference between these two cell populations with regard to Twist function [87]. While Battula et al. found that constitutive overexpression of Twist allowed EMT-derived cells to differentiate into multiple mesodermal lineages in vitro [86], Isenmann et al. had previously reported that overexpression of Twist in BM-MSCs blocked their osteogenic and chondrogenic differentiation in vitro and inhibited their ability to form ectopic ossicles in vivo [88]. It was speculated that these conflicting results might be explained by differences in gene dosage as the composition of Twist homodimers versus heterodimers influences its regulatory function [87]. Another possible explanation raised by Phinney was that the breast epithelial cells used by Battula et al. [86] were immortalized and Twist is known to interact with tumor suppressor proteins, such as p53 [87]. Therefore, more appropriated models for EMT induction and stringent assays to assess the stemness of EMT-derived cells are needed to compellingly answer whether activation of EMT could generate genuine MSCs.
In another study, Medici et al. observed that heterotopic bony tissue formed in patients with fibrodysplasia ossificans progressiva (FOP) is positive for endothelial cell markers, such as TIE2 and von Willebrand factor [89]. Since FOP is caused by constitutive activation of the ALK-2 receptor, the authors overexpressed ALK-2 in endothelial cells and investigated their phenotype. Overexpression of constitutively active ALK-2 induced the loss of endothelial markers and acquisition of a mesenchymal phenotype in endothelial cells. Such activation of EndMT was also observed after incubation of endothelial cells with TGF-β2 or bone morphogenetic protein-4, both ligands of ALK-2. Markedly, EndMT-derived cells became positive for MSC markers, such as STRO-1, CD44, and CD90, and were also capable of osteogenic, chondrogenic, and adipogenic differentiation in vitro.
Indeed, the generation of MSC-like osteoprogenitors through activation of EndMT provides a suitable mechanistic explanation for the clinical manifestations of FOP. However, it remains to be determined whether activation of EndMT represents a biologically relevant source of genuine MSCs in vivo under physiological homeostasis since the endothelium is known to be quiescent unless perturbed by pathological circumstances such as tissue injury, inflammation, and cancer.
Concluding Remarks
MSCs have been often defined using in vitro properties that are shared with other stromal cells and are not stringent enough to access stemness. Based on these criteria, stromal cells occupying distinct niches and also derived from biological processes such as the EMT/EndMT have been reported to originate cultured cells with the in vitro properties of MSCs. This approach has limited the precise identification of a bona fide MSC endowed with the capacity to self-renew in vivo and to regenerate the stromal compartment during cellular turnover and tissue injury.
It is currently recognized that different organs posses their own MSC population whose biological properties such as differentiation capacity and expression of molecular markers are influenced by their native microenvironment. However, except for the bone where evidences point for the existence of an additional pool of extravascular MSCs, the location at the perivascular space is a common feature of MSCs studied in virtually all organs so far. However, the determination of their identity within the vascular wall has been controversial.
Recent reports identifying molecular markers capable of discriminating perivascular populations in adipose tissue and stringent in vivo lineage tracing studies in long bones and incisor teeth have been challenging the claim that MSCs would correspond to pericytes in vivo. Evidences from the comparison between cultured adventitial cells and pericytes from adipose tissue and also from lineage tracing experiments in the incisor teeth indicate that the adventitia contains genuine MSCs that are responsible for originating all the stromal progeny, including pericytes. In this scenario, MSCs are perivascular adventitial stromal cells capable of replenishing the local mesenchyme during tissue homeostasis and injury repair (Fig. 1). This model accommodates both adventitial cells and pericytes as critical cellular elements for tissue maintenance and repair. In this model, pericytes represent a more stable cell phenotype with minimal contribution to tissue homeostasis in the absence of injury. However, upon tissue injury, pericytes would be activated in response to surrounding signals and act as genuine MSCs, thereby promoting tissue repair. Indeed, the concept of cultured MSCs as being activated pericytes has already been postulated [90].
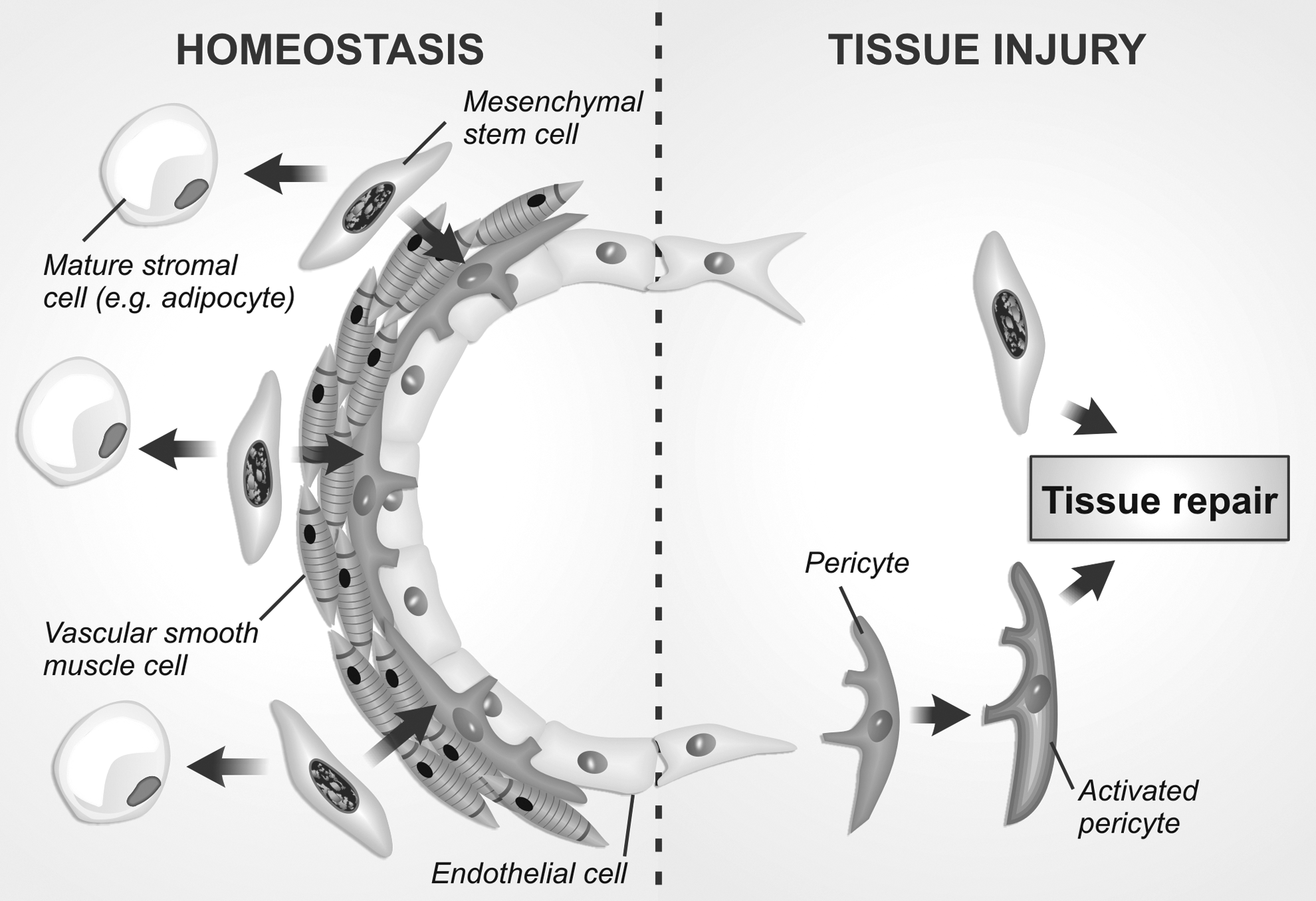
A model for the in vivo identity and function of perivascular MSCs. Recent evidence suggest that perivascular MSCs occupy the outermost perivascular cell layer of blood vessels, acting as adventitial cells. During homeostasis (left), MSCs would behave as progenitor cells of perivascular populations (eg, smooth muscle cells and pericytes) and of surrounding stromal cells, such as adipocytes. After a tissue injury (right), pericytes can be activated by microenvironmental signals and promote tissue repair along with MSCs through direct differentiation and/or secretion of proregenerative molecules. MSCs, mesenchymal stem cells.
The mesenchyme turnover is slow in most organs under homeostasis. However, in the face of an injury, the regenerative response is comparatively faster. In this case, the activation of pericytes to a proregenerative state by local signaling would be very convenient since pericytes span all types of blood vessels and therefore can provide a rapid response against tissue injury [90].
In conclusion, the use of nonstringent criteria to access MSC stemness represented a major roadblock that delayed the clarification of MSC native identity. However, the use of molecular markers to separate adventitial cells from other perivascular populations and evidences from refined lineage tracing models indicate that the perivascular MSCs might correspond to adventitial fibroblastic cells in vivo and that these cells might even originate pericytes in some organs. Furthermore, stringent in vivo studies should be encouraged since they have been promoting conspicuous shifts in the concept of MSCs, paving the way for clarifying their native activity and for refining their application as therapeutic tools.
Footnotes
Acknowledgments
This work was supported with grants from the Coordination for the Improvement of Higher Level Education (CAPES), São Paulo Research Foundation (FAPESP), and National Counsel of Technological and Scientific Development (CNPq), Brazil.
Author Disclosure Statement
No competing financial interests exist.